Resumos
O estudo da marcha de absorção de nutrientes e da produção de matéria seca em razão do estádio de desenvolvimento das culturas é de fundamental importância para subsidiar estratégias de adubação. Este trabalho objetivou determinar a produção de matéria seca e o acúmulo de macronutrientes na parte aérea das plantas de Guizotia abyssinica (niger), em diferentes estádios de crescimento e desenvolvimento. O delineamento experimental foi em blocos ao acaso, com cinco repetições. Amostras da parte aérea das plantas foram coletadas aos 15, 30, 45, 60, 75 e 90 dias após a emergência (DAE). Em cada coleta, as plantas foram divididas em folha, caule + ramos, capítulo e aquênios (grãos), secas até peso constante para posterior determinação da massa de matéria seca e teor de N, P, K, Ca, Mg e S. Os teores dos macronutrientes na parte aérea das plantas de niger, seguiram a seguinte ordem de exportação de nutrientes: N > Ca > Mg > P > K > S. Em contrapartida, quanto aos teores dos nutrientes nos grãos por ocasião da colheita do experimento, a quantidade dos elementos exportados seguiu a ordem: N > P > Ca > Mg > S > K. Atenção especial deve ser dada à manutenção da adequada disponibilidade de N e P nas fases inicias por causa do ciclo curto e do rápido surgimento das estruturas reprodutivas. A exportação de nutrientes e o índice de colheita são baixos para as plantas de niger, resultando maior oferta de nutrientes para o cultivo em rotação.
Guizotia abyssinica; marcha de absorção; macronutrientes; biodiesel
Studying nutrient uptake and dry matter accumulation in plants as affected by crop developmental stages is essential to establish and assist strategies for crop fertilization. The aim of this study was to assess shoot dry matter and macronutrient accumulation in Guizotia abyssinica (niger) at different growth and development stages. A randomized block experimental design with five replications was used. Shoot samples of the plants were taken at 15, 30, 45, 60, 75, and 90 days after seedling emergence (DAE). At each sampling, plants were separated into leaves, stem + branches, flower heads, and achenes. The samples were then dried until reaching constant weight to evaluate dry matter and N, P, K, Ca, Mg, and S contents. Macronutrient concentration in niger shoots showed the following order of nutrient export: N > Ca > Mg > P > K > S. Conversely, concentration in achenes at harvest followed the order N > P > Ca > Mg > S > K. Special attention must be paid to maintaining appropriate availability of N and P in initial growth stages due to the short developmental cycle and early flowering of niger. Nutrient export and harvest index are low in niger plants, resulting in a higher nutrient supply for the crop in rotation.
Guizotia abyssinica; nutrient uptake rate; biodiesel
INTRODUÇÃO
O estudo da marcha de absorção de nutrientes e da produção de matéria seca por causa do estádio de desenvolvimento das culturas é de fundamental importância para subsidiar estratégias de definição das quantidades e das épocas de realizações de adubação e das quantidades mínimas que devem ser restituídas ao solo para fins de manutenção da fertilidade. Isso contribui para o aumento da eficiência no manejo das culturas, proporcionando ganhos em produtividade e redução de custos, com o consequente uso racional e eficiente dos insumos aplicados no solo (Franco, 2011Franco AAN. Marcha de absorção e acúmulo de nutrientes na cultura do sorgo (dissertação). Janaúba: Universidade Estadual de Montes Claro; 2011.).
O niger (Guizotia abyssinica), uma oleaginosa da família das Asteraceae, nativa da África, tem grande potencial para produção de óleo comestível de alta qualidade, pois, além do elevado teor de óleo (30 a 45 %), apresenta excelente qualidade do perfil de ácidos graxos poli-insaturados, composto principalmente pelo ácido linoleico, cerca de 70 % (Ramdan e Morsel, 2003Ramdan MF, Morsel JT. Determination of the lip classes and fatty acid profile of Niger (Guizoitia abyssinica Cass.) seed oil. Phytochem Anal. 2003;14:366-70.). O ácido linoleico desempenha importante função na estrutura das membranas celulares e nos processos metabólicos, como a redução dos riscos de doenças cardiovasculares, por meio das melhorias dos teores de triglicérides, colesterol total e da lipoproteína de baixa densidade (Fagherazi et al., 2008Fagherazi S, Dias RL, Bortolon, F. Impacto do exercício físico isolado e combinado com dieta sobre os níveis séricos de HDL, LDL, colesterol total e triglicérides. R Bras Med Esporte. 2008;14:381-6.). A utilização do óleo de niger está ligada à fonte de proteína na dieta humana, à indústria química na produção de sabonetes, tintas lubrificantes e biodiesel (Sarin et al., 2009Sarin R, Sharamaa M, Khanb AA. Studies on Guizotia abyssinica L. oil: Biodiesel synthesis and process optimization. Biores Technol Essex. 2009;100:4187-92.).
O niger é uma planta dicotiledônea herbácea anual, que pode atingir um porte de 0,5-1,5 m de altura. Suas folhas são opostas, sésseis, ovaladas-lanceoladas, serrilhadas, podendo atingir em torno de 22 cm de comprimento. Já suas flores são amarelas, a polinização é cruzada, realizada por abelhas (Getinet e Sharma, 1996Getinet A, Sharma SM. Niger, Guizotia abyssinica (L. f.) Cass., promoting the conservation and use of underutilized and neglected crops. Rome: International Plant Genetic Resources Institute, 1996.; Bessa et al., 2008Bessa OR, Lima MV, Elias KFM, Fraga AC, Castro Neto P. Rendimento de extração mecânico-química e caracterização físico-química do óleo de niger (Guizotia abyssinica). In: Anais do 5º Congresso Brasileiro de Plantas Oleaginosas, Óleos, Gorduras e Biodiesel [CD-ROM]; 2008; Lavras. Lavras; Universidade Federal de Lavras; 2008.). As sementes são negras brilhantes, e muito leves, com peso de 1.000 sementes variando de 3 a 5 g (Ramdan e Morsel, 2002Ramdan MF, Morsel JT. Proximate neutral lipid composition of Niger (Guizotia abyssinica Cass.) seed. Czech J Food Sci. 2002;20:98-104.).
Essa espécie é cultivada com sucesso em sistemas de rotação com a soja, milho e trigo (Getinet e Sharma, 1996Getinet A, Sharma SM. Niger, Guizotia abyssinica (L. f.) Cass., promoting the conservation and use of underutilized and neglected crops. Rome: International Plant Genetic Resources Institute, 1996.) e tem sido estudada no Brasil como alternativa para compor sistema de rotação de cultura na safrinha, em regiões onde o período de semeadura de culturas tradicionais como o milho é limitado por restrições térmicas e hídricas (Ambrosano, 2012Ambrosano L. Avaliação de plantas oleaginosas potenciais para cultivo na safrinha [dissertação]. Lavras: Universidade Federal de Lavras; 2012.; Franchini, 2014Franchini RG. Estudos de sistemas de produção, envolvendo rotação de culturas, com ênfase em culturas para produção de grãos, de óleo e de torta em plantio direto [tese]. Dourados: Universidade Federal da Grande Dourados; 2014.). Contudo, ainda não há informações disponíveis na literatura sobre a demanda por nutrientes pela cultura do niger. Nesse aspecto, o conhecimento da marcha de absorção dos nutrientes pelo niger é essencial para definir as estratégias de adubação para obter elevados rendimentos por essa cultura (Zobiole et al., 2010Zobiole LHS, Castro C, Oliveria FA, Oliveira Junior A. Marcha de absorção de macronutrientes na cultura do girassol. R Bras Ci Solo. 2010;34:425-33.).
Objetivou-se com este trabalho determinar a produção de matéria seca e o acúmulo de macronutrientes na parte aérea das plantas de Guizotia abyssinica (niger), em diferentes estádios de crescimento e desenvolvimento.
MATERIAL E MÉTODOS
O experimento foi realizado na Fazenda Experimental de Ciências Agrárias da Universidade Federal da Grande Dourados (UFGD), município de Dourados, MS, localizado nas coordenadas geográficas de 54° 56' W e 22° 12' S e a 452 m de altitude. O clima conforme a classificação de Köppen é do tipo Cwa, que se caracteriza como mesotérmico úmido com verão chuvoso. As precipitações pluviais e as temperaturas médias, registradas durante a realização do experimento, estão apresentadas na figura 1. O solo da área experimental foi classificado como Latossolo Vermelho distroférrico muito argiloso (Embrapa, 2013Empresa Brasileira de Pesquisa Agropecuária - Embrapa. Sistema brasileiro de classificação de solos 3ª ed. Brasília, DF; 2013.), apresentando nos 20 cm superficiais MO: 23,6 g dm-3; pH(CaCl2): 5,9; P(resina): 16,6 mg dm-3; K+, Ca2+e Mg2+: 5,6; 57,4 e 19,5 mmolc dm-3, respectivamente; S-SO4-2: 6,6 mg dm-3 e V = 76 %. A análise granulométrica do solo desta camada revelou valores de: 644, 203 e 152 g kg-1 de argila, silte e areia, respectivamente.
Precipitação pluvial mensal e temperatura média no período experimental. Fonte: Estação Agrometereológica Embrapa Agropecuária Oeste, Dourados, MS.
O delineamento experimental foi em blocos ao acaso, com seis épocas de coleta (15, 30, 45, 60, 75 e 90 dias, após a emergência das plântulas) e cinco repetições, totalizando 30 unidades experimentais. A semeadura do niger foi realizada em maio de 2013, utilizando-se uma semeadora-adubadora de 13 linhas. A adubação foi de 300 kg ha-1 da fórmula 08-20-20 de NPK.
Cada unidade experimental foi constituída de seis linhas de 10,0 m de comprimento, espaçadas entre si por 0,45 m e densidade de semeadura de 10 plantas m-1. A área útil foi constituída de quatro linhas centrais, desprezando-se 1,0 m em ambas as extremidades.
Amostras da parte aérea das plantas foram coletadas aos 15, 30, 45, 60, 75 e 90 dias, após a emergência (DAE). Em cada metro de linha, foram coletadas 10 plantas. A cada coleta, as plantas foram divididas em folha (limbo e pecíolo), caule + ramos, capitulo e aquênios. As diversas partes das plantas foram lavadas e secas em estufa de circulação forçada de ar a 65 ºC, até atingirem peso constante. Em seguida, quantificou-se o peso da matéria seca das amostras e essas foram moídas em moinho tipo Willey. O teor de N, P, K, Ca, Mg e S no tecido vegetal foi determinada em cada parte da planta, de acordo com os procedimentos propostos por Malavolta et al. (1997)Malavolta E, Vitti GC, Oliveira SA. Avaliação do estado nutricional das plantas: Princípios e aplicações. Piracicaba: Potafos; 1997..
O acúmulo de matéria seca e dos nutrientes, em cada estrutura da planta e na planta inteira em função dos dias após a semeadura foram ajustados ao modelo Gaussian com três parâmetros (Zobiole et al., 2010Zobiole LHS, Castro C, Oliveria FA, Oliveira Junior A. Marcha de absorção de macronutrientes na cultura do girassol. R Bras Ci Solo. 2010;34:425-33.), descrito a partir da equação 1:

em que ŷ é o acúmulo de nutrientes; a, o valor de máximo acúmulo; x0, o valor de x, em DAE, que proporciona o máximo em ŷ; e b, a amplitude no valor de x, em DAE, entre o ponto de inflexão e o ponto de máximo. Assim, a partir do modelo ajustado, foi possível determinar, com exatidão, o valor do ponto de inflexão (PI) na curva de acordo com a equação 2:

Matematicamente, o PI corresponde ao valor de x em que a curvatura do modelo ajustado muda de sinal; fisiologicamente, isso corresponde ao valor de x, em DAE, em que a taxa de acúmulo diário, ainda que positiva, passa a decrescer.
RESULTADO E DISCUSSÕES
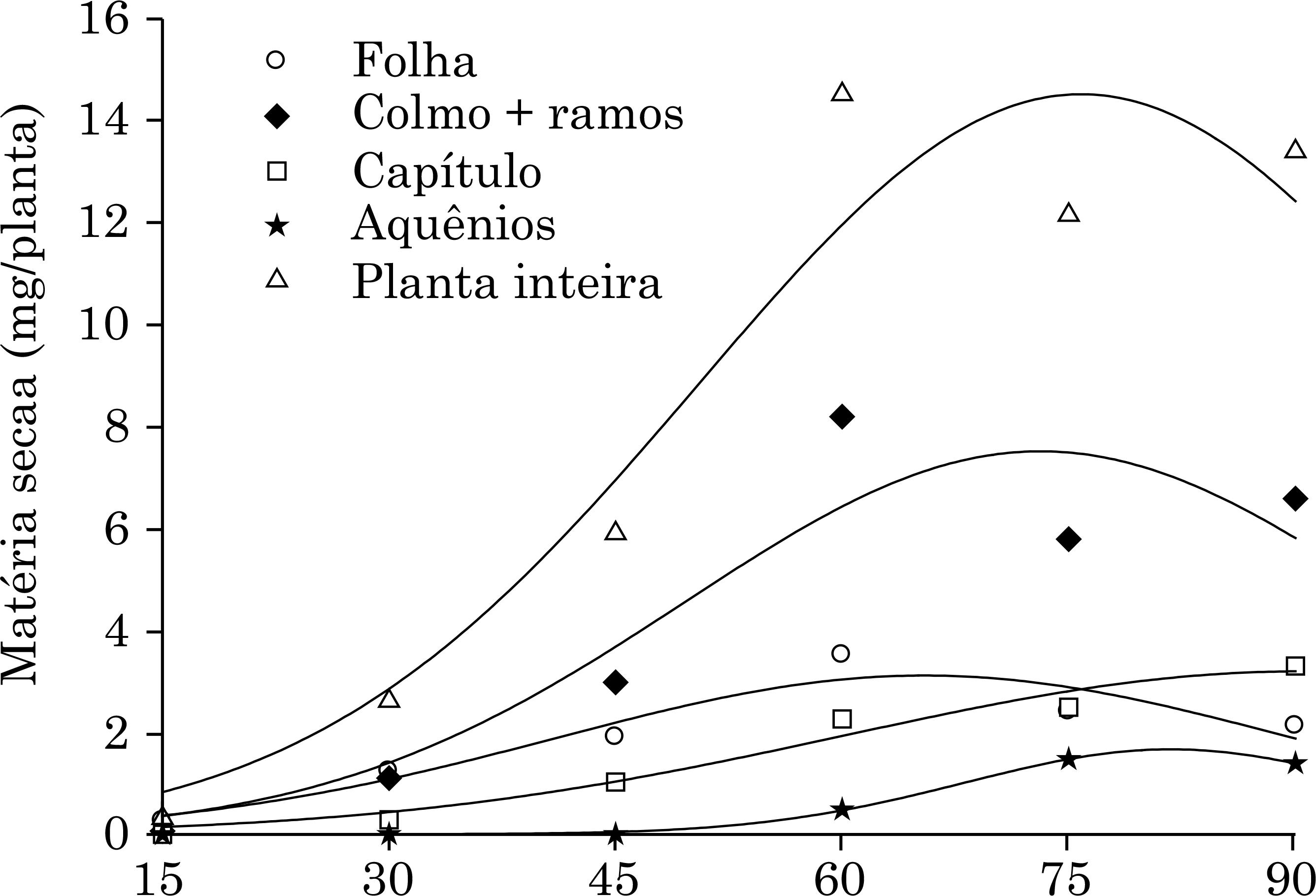
A produção de massa seca das plantas de niger foi lenta até o 30º DAE (Figura 1), com folhas e colmos contribuindo com percentuais próximos (47,1 e 42,1 %, respectivamente) para o incremento de massa de matéria seca nesse período. Essa observação pode ser explicada pela forma de roseta adquirida pela planta, com grande número de folhas expandidas e entrenós curtos.
A partir dos 30 DAE, os colmos + ramos começam a contribuir de forma mais intensa para a produção de matéria seca total (Figura 2), sendo o valor máximo dessas estruturas alcançado aos 73 dias após a emergência das plântulas, com 7,50 g/planta (Figura 2, Quadro 1). Esse comportamento é explicado pelo início do crescimento das ramificações que brotaram das axilas das folhas do colmo principal. Constatação similar foi relatada por Ambrosano (2012)Ambrosano L. Avaliação de plantas oleaginosas potenciais para cultivo na safrinha [dissertação]. Lavras: Universidade Federal de Lavras; 2012., o qual verificou intenso crescimento das ramificações e do diâmetro de colmo de plantas de niger no período de 30 a 60 DAE, em Lavras, MG.
Produção de matéria seca da parte aérea das plantas de niger em função dos dias após a emergência (DAE).
Estimativa dos parâmetros das equações de regressões de acordo com o modelo Gaussian ajustadas para a produção de matéria seca e o acúmulo de macronutrientes nas diferentes estruturas da parte aérea de plantas de niger em função do tempo (dia) desde a emergência e os respectivos tempos em que ocorreu o ponto de inflexão
A produção máxima de matéria seca da planta inteira ocorreu aos 76 dias com 14,51 g (Quadro 1, Figura 2). Para os grãos, a produção máxima foi alcançada aos 82 dias após a emergência, com 1,16 g/planta. Tomando-se como base os teores dos macronutrientes no final do experimento, na planta inteira e nos aquênios (Quadro 2), observou-se, respectivamente, a ordem: N > Ca > Mg > P > K > S e N > P > Ca > Mg > S > K. De acordo com os teores de nutrientes nos aquênios do niger (Quadro 2) e massa de matéria seca dos aquênios, verificou-se exportação de nutrientes na ordem de 8,66; 2,23; 0,45; 1,20; 0,90; e 0,66 kg ha-1 para N, P, K, Ca, Mg e S, respectivamente. A produtividade de grãos foi de 258 kg ha-1. Essa produtividade ficou dentro dos valores relatados na literatura para essa espécie, de 200 a 300 kg ha-1 (Getinet e Sharma, 1996)Getinet A, Sharma SM. Niger, Guizotia abyssinica (L. f.) Cass., promoting the conservation and use of underutilized and neglected crops. Rome: International Plant Genetic Resources Institute, 1996.. Nesse sentido, informações referentes às quantidades de nutrientes extraídos e exportados via grãos são relevantes para o programa de recomendações de adubação (Werle et al., 2008)Werle R, Garcia RA, Rosolem CA. Lixiviação de potássio em função da textura e da disponibilidade do nutriente no solo. R Bras Ci Solo. 2008;32:2297-305..
Teores médios de N, P, K, Ca, Mg e S na planta inteira e nos componentes da parte aérea de planta de niger em função dos dias após a emergência (DAE)
Os teores de N nas plantas de niger foram elevadas desde as fases iniciais de desenvolvimento, evidenciando a importância da disponibilidade de N às plantas por ocasião da semeadura (Quadro 2). No caso das folhas, aos 15 dias após a emergência (DAE), o teor de N foi de 63,28 g kg-1, com posterior decréscimo ao longo do desenvolvimento das plantas, visto que outras estruturas das plantas começaram a se desenvolver, como os capítulos e aquênios (Figura 2). Como o início da formação das estruturas reprodutivas é relativamente rápido, aos 30 dias após a emergência, acredita-se que, logo nas fases iniciais de desenvolvimento das folhas, as plantas acumularam N para suprir a posterior demanda dos capítulos e aquênios.
O crescimento vegetativo promoveu o desenvolvimento rápido da área foliar e com isso a formação de um aparto fotossintético capaz de suportar rendimentos elevados. As plantas de niger também podem ter acumulado proteínas de armazenamento vegetativo que poderão posteriormente ser translocadas para os grãos. Staswick et al. (1991)Staswick PE, Huang JF, Rhee Y. Nitrogen and methyl jasmonate induction of soybean vegetative storage protein genes. Plant Physiol. 1991;96:130-6. e Staswick (1994)Staswick PE. Storage proteins of vegetative plant tissues. Ann Rev Plant Physiol Plant Mol Biol. 1994;45:303-22. relataram a importância de proteínas armazenadas nos tecidos vegetativos, que servem para armazenar N durante condições de suprimento adequado desse nutriente e que, durante o enchimento de grãos, é redistribuído para esses órgãos. Um fator que pode ter contribuído para esses valores elevados de N no tecido vegetal é que, entre as dicotiledôneas, normalmente as espécies da família Asteraceae apresentam as maiores quantidades de frutano, que, depois do amido e da sacarose, é o carboidrato mais distribuído nos vegetais (Meier e Reid, 1982Meier H, Reid JSG. Reserve polysaccharides other than starch in higher plants. In: Loewus FA, Tanner W, editors. Encyclopedia of plant physiology. New series. Plant carbohydrates I. London: Springer-Verlag; 1982. 13A. p.418-471.). A elevação nos teores de frutano sugere aumento nas atividades das enzimas de síntese, evidenciando a participação do N nesse processo (Taiz e Zeiger, 2013Taiz L, Zeiger E. Fisiologia vegetal. 5ª ed. Porto Alegre: Artmed; 2013.).
Quanto ao acúmulo de N na parte aérea das plantas (Figura 3), as folhas e os colmos foram responsáveis pelo armazenamento de grande parte do N extraído até os 60 DAE. Além dos elevados teores nessas estruturas das plantas até esse período (Quadro 2), a contribuição na produção de fitomassa também foi maior (Figura 2), ocasionando maiores quantidades de N armazenadas nas folhas e nos colmos. A partir dos 60 DAE, fica evidente a contribuição das estruturas vegetativas com nitrogênio para a formação dos capítulos e grãos das plantas de niger. De acordo com Marschner (2012)Marschner H. Mineral nutrition of higher plants. Amsterdam: Academic Press; 2012., o N é um elemento móvel na planta e transloca-se facilmente de um órgão para outro; no florescimento é o período em que há maior translocação da parte aérea para a inflorescência. Contudo, a translocação depende de vários fatores, entre eles do teor de N na parte aérea, que no caso do niger nesta pesquisa foi alta, intensificando esse processo (Quadro 2).
Acúmulo de nitrogênio, fósforo, enxofre, potássio, cálcio e magnésio na planta inteira e nos diferentes componentes da parte aérea das plantas de niger em função dos dias após emergência (DAE).
Para o P, o máximo acúmulo considerando a planta inteira foi de 53,36 mg/planta e ocorreu aos 72 DAE, enquanto para o capítulo e aquênio ocorreram aos 74 e 80 DAE, com 17,73 e 15,52 mg/planta, respectivamente (Quadro 3). Comportamento similar às plantas inteiras foi detectado para os acúmulos de P nas folhas e nos caules + ramos, obtidos próximo aos 62 DAE, o que coincide com o início da maturação dos aquênios, proveniente dos primeiros capítulos que surgiram 30 DAE (Figura 2). O niger produz vários capítulos em suas ramificações; assim como o início do florescimento aos 30 DAE (Figura 2), os primeiros aquênios chegaram à maturação cerca de 30 após o surgimento dos capítulos, ou seja, 60 DAE. Nesse momento, há na planta de niger capítulos e aquênios em diferentes estádios de desenvolvimento. O máximo acúmulo determinado a partir do modelo ajustado ocorreu aos 61 e 68 dias para a produção de matéria seca de capítulo e aquênios, respectivamente (Quadro 1), enquanto para o acúmulo de P foi aos 52 e 67 dias para os capítulos e os aquênios, respectivamente (Quadro 3). Esses resultados evidenciaram grande demanda por P a partir do surgimento dos primeiros capítulos (30 DAE) e se intensificaram a partir do inicio da maturação dos aquênios dos primeiros capítulos (Figura 3).
O aumento da demanda de P para a formação dos capítulos e aquênios foi atendido a partir da realocação do P contido em folhas e caule + ramos, que apresentaram decréscimos a partir de 40 e 50 DAE, respectivamente (Figura 3, Quadro 3). Assim, a disponibilidade de P nas fases iniciais das plantas foi de extrema importância, em razão do rápido surgimento dos capítulos e consequente formação dos aquênios. Constatações similares foram observadas no trabalho de Zobiole et al. (2010)Zobiole LHS, Castro C, Oliveria FA, Oliveira Junior A. Marcha de absorção de macronutrientes na cultura do girassol. R Bras Ci Solo. 2010;34:425-33. com a cultura do girassol e no de Mauad et al. (2013)Mauad M, Garcia RA, Vitorino ACT, Silva RMF, Garbiate MV, Coelho CF. Matéria seca e acúmulo de macronutrientes na parte aérea das plantas de Crambe. Ci Rural. 2013;43:771-8. com a cultura do crambe, em que o máximo acúmulo de P foi obtido na formação e no enchimento dos grãos, uma vez que o P contido em folhas e colmos + ramos tem considerável contribuição na formação dos aquênios da oleaginosa.
Levando em consideração a produção dos aquênios, o P passou a ser o segundo elemento mais requerido pelas plantas. Uma possível explicação para esse fato está relacionada ao acúmulo de lipídios; enquanto para a produção de uma unidade de carboidrato a planta necessita de 1,2 unidade de glicose, para a produção da mesma quantidade de lipídeos essa consome três unidades de glicose (Penning de Vries, 1974Penning de Vries FWT. Substrate utilization and respiration in relation to growth and maintenance in higher plants. Neth J Agric Sci. 1974;22:40-4.). Cabe lembrar que a glicose é quebrada durante a respiração e gera compostos ricos em energia como ATP, que necessita do P para sua síntese.
O K foi o nutriente exigido em menores quantidades pelas plantas de niger, tendo em vista que o valor do ponto de máximo de acúmulo de K foi de 37,03 mg/planta (Quadro 3), aos 70 DAE. Comportamento semelhante foi relatado por Mauad et al. (2013)Mauad M, Garcia RA, Vitorino ACT, Silva RMF, Garbiate MV, Coelho CF. Matéria seca e acúmulo de macronutrientes na parte aérea das plantas de Crambe. Ci Rural. 2013;43:771-8., os quais, estudando o acúmulo de nutrientes em plantas de Crambe abyssinica, observaram que o K foi o nutriente de menor acúmulo por essa oleaginosa.
Quanto à marcha de absorção do K (Figura 3), destacam-se os caules + ramos com 20,26 mg/planta como os órgãos que mais acumularam esse elemento, seguido pelas folhas com 9,19 mg/planta, capítulo com 9,07 mg/planta e aquênios com 2,80 mg/plant (Figura 3, Quadro 3). O capítulo parece ser o dreno preferencial do K, enquanto os caules e ramos são as fontes para essa redistribuição (Quadro 2).
Observou-se que aproximadamente 92 % do K estavam alocados no caule + ramos, nas folhas e nos capítulos das plantas de niger. Dessa forma, pode-se dizer que a maior parte do K absorvido pelo niger é passível de retornar ao solo, pois a liberação de K da palhada é rápida (Rosolem et al., 2006Rosolem CA, Garcia RA, Foloni JSS, Calonego JC. Lixiviação de potássio no solo de acordo com suas doses aplicadas sobre palha de milheto. R Bras Ci Solo. 2006;30:813-9.).
O Ca foi o segundo elemento mais exigido pelas plantas de niger com o máximo acúmulo de 201,86 mg/planta (Quadro 4). Esse nutriente parece ter papel de destaque em oleaginosas, uma vez que Mauad et al. (2013)Mauad M, Garcia RA, Vitorino ACT, Silva RMF, Garbiate MV, Coelho CF. Matéria seca e acúmulo de macronutrientes na parte aérea das plantas de Crambe. Ci Rural. 2013;43:771-8. observaram que para a cultura do crambe o Ca foi o segundo elemento mais exigido, e o mesmo foi relatado por Zobiole et al.(2010)Zobiole LHS, Castro C, Oliveria FA, Oliveira Junior A. Marcha de absorção de macronutrientes na cultura do girassol. R Bras Ci Solo. 2010;34:425-33. para a cultura do girassol que extraiu 116 kg ha-1 de Ca.
O acúmulo de cálcio nos órgãos das plantas seguiu a seguinte ordem: folhas > colmo + ramos > capítulo > aquênio (Quadro 4). Os teores de Ca nas folhas e nos colmos + ramos começam a diminuir a partir de 60 DAE, enquanto os teores nos capítulos e aquênios apresentaram pouca variação (Figura 3, Quadro 4). O decréscimo nos teores de Ca nas folhas e no colmo + ramos está relacionado ao menor crescimento e à demanda por esses órgãos, uma vez que o ponto de máxima da curva para a produção de matéria seca ocorreu aos 41 e 50 dias para folha e colmo, respectivamente.
Uma possível explicação para a ausência de queda acentuada nos teores de Ca no capítulo e aquênio (Figura 3, Quadro 4) está relacionada ao crescimento e desenvolvimento da planta de niger. O Ca é um elemento estrutural que participa da constituição da parede celular (Marschner, 2012Marschner H. Mineral nutrition of higher plants. Amsterdam: Academic Press; 2012.); assim, durante o ciclo da planta de niger ocorre emissão e formação de vários capítulos que começam a surgir a partir de 30 DAE (Figura 2) e atingem o máximo aos 62 DAE (Quadro 1), isso faz com que ocorra grande demanda de Ca para formação e constituição dos capítulos e aquênios, o que torna esses dois órgãos o dreno principal na planta, mantendo seus teores pouco alteradas até o final do ciclo.
O acúmulo de Mg nas plantas de niger seguiu a mesma tendência que para o Ca. O máximo acúmulo desse nutriente foi 66,93 mg/planta e ocorreu aos 70 dias (Quadro 4). Até 30 DAE, folhas e colmos + ramos apresentaram a mesma contribuição para o acúmulo de Mg (Figura 3); após esse período, colmos e ramos contribuíram de maneira mais efetiva, com o máximo acumulado de 35,72 mg/planta aos 67 DAE (Quadro 4). Por serem elementos móveis na planta, colmo e ramos são as principais fontes de Mg para o capítulo e aquênios (Figura 3, Quadro 2), embora a exportação de Mg pelos aquênio tenha sido baixa.
A curva de acúmulo de S apresentou comportamento muito semelhante ao Ca e Mg, o ponto de inflexão foi verificado aos 46 DAE, bem como o ponto de máxima de acúmulo de nutrientes, aos 66 (Quadro 4). Isso indica que o acúmulo desses nutrientes segue a mesma tendência, variando somente a magnitude da quantidade acumulada, que foi maior para o Ca, seguido pelo Mg. O acúmulo de S seguiu a seguinte ordem: colmo + ramos > folhas > capitulo > aquênio.
CONCLUSÕES
Os teores dos macronutrientes na parte aérea das plantas de niger seguiram a seguinte ordem de exportação de nutrientes: N > Ca > Mg > P > K > S. Em contrapartida, quanto aos teores dos nutrientes nos grãos por ocasião da colheita do experimento, a quantidade dos elementos exportados seguiu a ordem: N > P > Ca > Mg > S > K.
Atenção especial deve ser dada à manutenção da adequada disponibilidade de N e P nas fases inicias em razão do ciclo curto e do rápido surgimento das estruturas reprodutivas.
A exportação de nutrientes e o índice de colheita são baixos para as plantas de niger, resultando maior oferta de nutrientes para o cultivo em rotação.
REFERÊNCIAS
- Ambrosano L. Avaliação de plantas oleaginosas potenciais para cultivo na safrinha [dissertação]. Lavras: Universidade Federal de Lavras; 2012.
- Bessa OR, Lima MV, Elias KFM, Fraga AC, Castro Neto P. Rendimento de extração mecânico-química e caracterização físico-química do óleo de niger (Guizotia abyssinica). In: Anais do 5º Congresso Brasileiro de Plantas Oleaginosas, Óleos, Gorduras e Biodiesel [CD-ROM]; 2008; Lavras. Lavras; Universidade Federal de Lavras; 2008.
- Empresa Brasileira de Pesquisa Agropecuária - Embrapa. Sistema brasileiro de classificação de solos 3ª ed. Brasília, DF; 2013.
- Fagherazi S, Dias RL, Bortolon, F. Impacto do exercício físico isolado e combinado com dieta sobre os níveis séricos de HDL, LDL, colesterol total e triglicérides. R Bras Med Esporte. 2008;14:381-6.
- Franchini RG. Estudos de sistemas de produção, envolvendo rotação de culturas, com ênfase em culturas para produção de grãos, de óleo e de torta em plantio direto [tese]. Dourados: Universidade Federal da Grande Dourados; 2014.
- Franco AAN. Marcha de absorção e acúmulo de nutrientes na cultura do sorgo (dissertação). Janaúba: Universidade Estadual de Montes Claro; 2011.
- Getinet A, Sharma SM. Niger, Guizotia abyssinica (L. f.) Cass., promoting the conservation and use of underutilized and neglected crops. Rome: International Plant Genetic Resources Institute, 1996.
- Malavolta E, Vitti GC, Oliveira SA. Avaliação do estado nutricional das plantas: Princípios e aplicações. Piracicaba: Potafos; 1997.
- Marschner H. Mineral nutrition of higher plants. Amsterdam: Academic Press; 2012.
- Mauad M, Garcia RA, Vitorino ACT, Silva RMF, Garbiate MV, Coelho CF. Matéria seca e acúmulo de macronutrientes na parte aérea das plantas de Crambe. Ci Rural. 2013;43:771-8.
- Meier H, Reid JSG. Reserve polysaccharides other than starch in higher plants. In: Loewus FA, Tanner W, editors. Encyclopedia of plant physiology. New series. Plant carbohydrates I. London: Springer-Verlag; 1982. 13A. p.418-471.
- Penning de Vries FWT. Substrate utilization and respiration in relation to growth and maintenance in higher plants. Neth J Agric Sci. 1974;22:40-4.
- Ramdan MF, Morsel JT. Proximate neutral lipid composition of Niger (Guizotia abyssinica Cass.) seed. Czech J Food Sci. 2002;20:98-104.
- Ramdan MF, Morsel JT. Determination of the lip classes and fatty acid profile of Niger (Guizoitia abyssinica Cass.) seed oil. Phytochem Anal. 2003;14:366-70.
- Rosolem CA, Garcia RA, Foloni JSS, Calonego JC. Lixiviação de potássio no solo de acordo com suas doses aplicadas sobre palha de milheto. R Bras Ci Solo. 2006;30:813-9.
- Sarin R, Sharamaa M, Khanb AA. Studies on Guizotia abyssinica L. oil: Biodiesel synthesis and process optimization. Biores Technol Essex. 2009;100:4187-92.
- Staswick PE, Huang JF, Rhee Y. Nitrogen and methyl jasmonate induction of soybean vegetative storage protein genes. Plant Physiol. 1991;96:130-6.
- Staswick PE. Storage proteins of vegetative plant tissues. Ann Rev Plant Physiol Plant Mol Biol. 1994;45:303-22.
- Taiz L, Zeiger E. Fisiologia vegetal. 5ª ed. Porto Alegre: Artmed; 2013.
- Werle R, Garcia RA, Rosolem CA. Lixiviação de potássio em função da textura e da disponibilidade do nutriente no solo. R Bras Ci Solo. 2008;32:2297-305.
- Zobiole LHS, Castro C, Oliveria FA, Oliveira Junior A. Marcha de absorção de macronutrientes na cultura do girassol. R Bras Ci Solo. 2010;34:425-33.
Datas de Publicação
-
Publicação nesta coleção
Mar-Apr 2015
Histórico
-
Recebido
7 Mar 2014 -
Aceito
22 Dez 2014