ABSTRACT
Cover crops contribute to nutrient cycling and may improve soil chemical properties and, consequently, increase crop yield. The aim of this study was to evaluate cover crop residue decomposition and nutrient release, and the effects of these plants on soil chemical properties and on onion (Allium cepa L.) yield in a no-tillage system. The experiment was carried out in an Inceptisol in southern Brazil, where cover crops were sown in April 2012 and 2013. In July 2013, shoots of weeds (WD), black oats (BO), rye (RY), oilseed radish (RD), oilseed radish + black oats (RD + BO), and oilseed radish + rye (RD + RY) were cut at ground level and part of these material from each treatment was placed in litter bags. The litter bags were distributed on the soil surface and were collected at 0, 30, 45, 60, 75, and 90 days after distribution (DAD). The residues in the litter bags were dried, weighed, and ground, and then analyzed to quantify lignin, cellulose, non-structural biomass, total organic carbon (TOC), N, P, K, Ca, and Mg. In November 2012 and 2013, onion crops were harvested to quantify yield, and bulbs were classified according to diameter, and the number of rotted and flowering bulbs was determined. Soil in the 0.00-0.10 m layer was collected for chemical analysis before transplanting and after harvesting onion in December 2012 and 2013. The rye plant residues presented the highest half-life and they released less nutrients until 90 DAD. The great permanence of rye residue was considered a protection to soil surface, the opposite was observed with spontaneous vegetation. The cultivation and addition of dry residue of cover crops increased the onion yield at 2.5 Mg ha-1.
Allium cepa; nutrient cycling; no-till system
INTRODUCTION
In southern Brazil, and specifically in the Itajaí Valley, onion (Allium cepa L.) is typically grown in a conventional tillage system (CTS), in which losses of soil, water, and nutrients are increased by water erosion (Castro et al., 2011Castro NEA, Silva MLN, Freitas DAF, Carvalho GJ, Marques RM, Gontijo Neto GF. Plantas de cobertura no controle da erosão hídrica sob chuvas naturais. Biosci J. 2011;27:775-85.). Since the beginning of the 1980s, adoption of the no-tillage system (NTS) has been encouraged. Cover crops, especially grasses such as black oat (Avena strigosa Schreb.) and rye (Secale cereale L.), and some dicots, especially oilseed radish (Raphanus sativus), are grown as single or mixed crops in fall/winter (Souza et al., 2013Souza M, Comin JJ, Leguizamón ES, Kurtz C, Brunetto G, Müller Júnior V, Ventura B, Camargo AP. Matéria seca de plantas de cobertura, produção de cebola e atributos químicos do solo em sistema plantio direto agroecológico. Cienc Rural. 2013;43:21-7. doi:10.1590/S0103-84782012005000150).
The sustainability of NTS depends on the quality and quantity of plant residues and the time they remain on the soil surface, among other factors (Teixeira et al., 2011Teixeira MB, Loss A, Pereira MG, Pimentel C. Decomposição e liberação de nutrientes da parte aérea de plantas de milheto e sorgo. Rev Bras Cienc Solo. 2011;35:867-76. doi:10.1590/S0100-06832011000300021). In the case of onions, cover crop residues are deposited between plant rows, especially in late winter and early spring. Residues protect the soil surface from the impact of raindrops and reduces erosion, suppresses the incidence of weeds, and increases water storage in the soil profile (Moraes et al., 2013Moraes PVD, Agostinetto D, Panozzo LE, Oliveira C, Vignolo GK, Markus C. Manejo de plantas de cobertura no controle de plantas daninhas e desempenho produtivo da cultura do milho. Semina: Cienc Agrár. 2013;34:497-508. doi:10.5433/1679-0359.2013v34n2p497; Chen et al., 2014Chen B, Liu E, Tian Q, Yan C, Zhang Y. Soil nitrogen dynamics and crop residues. A review. Agron Sust Dev. 2014;34:429-42. doi:10.1007/s13593-014-0207-8). Nutrients are gradually mineralized during cover crop decomposition, which reduces leaching and favors greater uptake by onion plants. During decomposition of plant residues, microbial populations use organic C as an energy source (Brunetto et al., 2014Brunetto G, Ceretta CA, Melo GWB, Kaminski J, Trentin G, Girotto E, Ferreira PAA, Miotto A, Trivelin PCO. Contribution of nitrogen from agricultural residues of rye to “Niagara Rosada” grape nutrition. Sci Hortic. 2014;169:66-70. doi:10.1016/j.scienta.2014.02.019), and N, P, K, Ca, and Mg are mineralized and they eventually return to the soil, increasing their availability and uptake during the onion cycle (Souza et al., 2013Souza M, Comin JJ, Leguizamón ES, Kurtz C, Brunetto G, Müller Júnior V, Ventura B, Camargo AP. Matéria seca de plantas de cobertura, produção de cebola e atributos químicos do solo em sistema plantio direto agroecológico. Cienc Rural. 2013;43:21-7. doi:10.1590/S0103-84782012005000150; Martins et al., 2014Martins RP, Comin JJ, Gatiboni LC, Roberto C, Soares F, Couto R, Brunetto G. Mineralização do nitrogênio de plantas de cobertura, solteiras e consorciadas, depositadas sobre um solo com histórico de cultivo de cebola. Rev Ceres. 2014;61:587-96. doi:10.1590/0034-737X201461040020).
Decomposition of plant residues and the consequent release of nutrients depend on climate and soil conditions, such as temperature, moisture, pH, oxygen, and content of plant nutrients in the soil; furthermore, the chemical composition of plant biomass, such as cellulose, lignin, and non-structural biomass contents are relevant, as well as C/N, lignin/N, and lignin/P ratios (Trinsoutrot et al., 2000Trinsoutrot I, Recous S, Bentz B, Linères M, Chèneby D, Nicolardot B. Biochemical quality of crop residues and carbon and nitrogen mineralization kinetics under nonlimiting nitrogen conditions. Soil Sci Soc Am J. 2000;64:918-26. doi:10.2136/sssaj2000.643918x; Tagliavini et al., 2007Tagliavini M, Tonon G, Scandellari F, Quiñones A, Palmieri S, Menarbin G, Gioacchini P, Masia A. Nutrient recycling during the decomposition of apple leaves (Malus domestica) and mowed grasses in an orchard. Agric Ecosyst Environ. 2007;118:191-200. doi:10.1016/j.agee.2006.05.018; Cornwell et al., 2008Cornwell WK, Cornelissen JHC, Amatangelo K, Dorrepaal E, Eviner VT, Godoy O, Hobbie SE, Hoorens B, Kurokawa H, Pérez-Harguindeguy N, Quested HM, Santiago LS, Wardle D, Wright IJ, Aerts R, Allison SD, van Bodegom P, Brovkin V, Chatain A, Callaghan TV, Díaz S, Garnier E, Gurvich D, Kazakou E, Klein JA, Read J, Reich PB, Soudzilovskaia NA, Vaieretti MV, Westoby M. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol Lett. 2008;11:1065-71. doi:10.1111/j.1461-0248.2008.01219.x; Teixeira et al., 2012Teixeira MB, Loss A, Pereira MG, Pimentel C. Decomposição e ciclagem de nutrientes dos resíduos de quatro plantas de cobertura do solo. Idesia. 2012;30:55-64. doi:10.4067/S0718-34292012000100007). In general, cover crop plant residues with more lignin and higher C/N, lignin/N, and lignin/P ratios, such as those of black oat and rye, decompose and release nutrients to the soil more slowly and gradually (Giacomini et al., 2003Giacomini SJ, Aita C, Hubner AP, Lunkes A, Guidini E, Amaral E. Liberação de fósforo e potássio durante a decomposição de resíduos culturais em plantio direto. Pesq Agropec Bras. 2003;38:1097-104. doi:10.1590/S0100-204X2003000900011). In contrast, residues with lower lignin content, and thus higher cellulose content and lower C/N, lignin/N, and lignin/P ratios, such as those of oilseed radish, have faster decomposition and subsequent nutrient release (Crusciol et al., 2005Crusciol CAC, Cottica RL, Lima EV, Andreotti M, Moro E, Marcon E. Persistência de palhada e liberação de nutrientes do nabo-forrageiro no plantio direto. Pesq Agropec Bras. 2005;40:161-8. doi:10.1590/S0100-204X2005000200009). Thus, intermediate levels of lignin, cellulose, non-structural biomass, contents of N, P, K, Ca, and Mg, and of C/N, lignin/N, and lignin/P ratios are expected with a mix of oilseed radish and a grass species such as black oat or rye. Therefore, mixtures of cover crop species aim to maintain adequate amounts of plant residues on the soil surface, in addition to gradual release of nutrients (Giacomini et al., 2003Giacomini SJ, Aita C, Hubner AP, Lunkes A, Guidini E, Amaral E. Liberação de fósforo e potássio durante a decomposição de resíduos culturais em plantio direto. Pesq Agropec Bras. 2003;38:1097-104. doi:10.1590/S0100-204X2003000900011).
In an NTS, single or mixed cover crops can take up nutrients from deeper layers of the soil and accumulate them in their roots or shoots. When placed on the soil surface, their decomposition alters nutrient content in surface layers throughout the next crop cycle, as shown for onions (Souza et al., 2013Souza M, Comin JJ, Leguizamón ES, Kurtz C, Brunetto G, Müller Júnior V, Ventura B, Camargo AP. Matéria seca de plantas de cobertura, produção de cebola e atributos químicos do solo em sistema plantio direto agroecológico. Cienc Rural. 2013;43:21-7. doi:10.1590/S0103-84782012005000150). Since onion plants have a low leaf area index, due to their reduced number of leaves and small size, use of cover crops is essential at the start of the NTS, which contributes to increases in onion yields and, in some cases, to reduction in fertilizer applications (Wang et al., 2008Wang G, Ngouajio M, Warncke DD. Nutrient cycling, weed suppression, and onion yield following brassica and sorghum sudangrass cover crops. HortTechnology. 2008;18:68-74. doi:10.1007/s13165-010-0002-z; Souza et al., 2013Souza M, Comin JJ, Leguizamón ES, Kurtz C, Brunetto G, Müller Júnior V, Ventura B, Camargo AP. Matéria seca de plantas de cobertura, produção de cebola e atributos químicos do solo em sistema plantio direto agroecológico. Cienc Rural. 2013;43:21-7. doi:10.1590/S0103-84782012005000150; Martins et al., 2014Martins RP, Comin JJ, Gatiboni LC, Roberto C, Soares F, Couto R, Brunetto G. Mineralização do nitrogênio de plantas de cobertura, solteiras e consorciadas, depositadas sobre um solo com histórico de cultivo de cebola. Rev Ceres. 2014;61:587-96. doi:10.1590/0034-737X201461040020). However, in southern Brazil, there are few studies on the dynamics of decomposition and nutrient release from cover crop residues and their impact on soil chemical properties and onion yield.
It is assumed that the different species of winter cover crops grown under no-tillage system, depending on their composition, can release different amounts of nutrients and promote different effects on the yield of onion and soil chemical properties. The aim of this study was to evaluate cover crop residue decomposition and nutrient release, as well as its effects on soil chemical properties and onion yield in a no-till system.
MATERIALS AND METHODS
Location of the experimental area and treatments
The experiment was carried out from July 2012 to November 2013 in the experimental area of the Empresa de Pesquisa e Extensão Agropecuária do Estado de Santa Catarina (Epagri) in Ituporanga, in the Upper Itajai Valley region, SC, Brazil (27° 22’ S, 49° 35’ W, and 475 m altitude). According to the Köppen classification system, the climate is humid subtropical (Cfa) (Kottek et al., 2006Kottek M, Grieser J, Beck C, Rudolf B, Rubel F. World map of the Köppen-Geiger climate classification updated. Meteorol Z. 2006;15:259-63. doi:10.1127/0941-2948/2006/0130). Climate data collected during the experiment are shown in figure 1. The soil was classified as Cambissolo Húmico (Souza et al., 2013Souza M, Comin JJ, Leguizamón ES, Kurtz C, Brunetto G, Müller Júnior V, Ventura B, Camargo AP. Matéria seca de plantas de cobertura, produção de cebola e atributos químicos do solo em sistema plantio direto agroecológico. Cienc Rural. 2013;43:21-7. doi:10.1590/S0103-84782012005000150), an Inceptisol (Soil Survey Staff, 2006Soil Survey Staff. Soil Taxonomy: a basic system of soil classification for making and interpreting soil surveys. 2nd ed. Washington, DC: US Government Printing Office: 2006.), with 380, 200, and 420 g kg-1 of clay, silt, and sand (clay loam texture), respectively (Claessen, 1997Claessen MEC, organizador. Manual de métodos de análise de solo. 2ª ed. Rio de Janeiro: Centro Nacional de Pesquisa de Solos; 1997.). At the beginning of the experiment (April 2009), the 0.00-0.10 m layer had the following chemical characteristics (Tedesco et al., 1995Tedesco MJ, Gianello C, Bissani CA, Bohnen H, Volkweiss SJ. Análises de solo, planta e outros materiais. 2ª. ed. Porto Alegre: UFRGS; 1995.): organic matter 40 g kg-1; pH(H2O) 6.0; P 26.6 mg kg-1 and K 145.2 mg kg-1 (extracted by Mehlich-1); Al3+ 0.0 cmolc kg-1; Ca2+ 7.2 cmolc kg-1; Mg2+ 3.4 cmolc kg-1 (extracted by 1 mol L-1 KCl); CECpH7.0 14.32 cmolc kg-1; base saturation (V) 76 %; and Al saturation (m) 34 %.
Rainfall and monthly minimum temperature in 2012 and 2013, and monthly average temperature in 2013 (a); Rainfall in 2012 and 2013, and daily average temperature in 2013 at 0, 30, 45, 60, 75, and 90 DAD in the experimental area (b).
The experiment was established in an area with a 20-years history of onion growing under conventional management (plowing and harrowing) until 1996. From that year on, a minimum-tillage system of onion was established with crop rotation and cover crops (black oat – Avena strigosa, velvet bean – Mucuna aterrima, pearl millet – Pennisetum glaucum, brown hemp – Crotalaria juncea, and common vetch – Vicia sativa). This system was in place from 1996 to 2007, and then sweet potato was grown until 2009. Since that time, an NTS experiment of onion has been carried out. In April 2009, weeds were desiccated and lime was applied and incorporated to raise pH in water to 6.0.
Treatments consisted of sowing single or mixed cover crops. A fallow treatment used as control was dominated by weeds (WD) comprising 20 botanical families, mostly bermuda grass (Cynodum sp.), bitter dock (Rumex obtusifolius), staggerweed (Stachys arvensis), purple amaranth (Amaranthus lividus), flatsedge (Cyperus spp.), radishroot woodsorrel (Oxalis corniculata), hairy beggarticks (Bidens pilosa), and gallant soldier (Galinsoga parviflora). Cover crops were black oats (Avena strigosa Schreb – 120 kg seeds ha-1) (BO), rye (Secale cereale L. – 120 kg seeds ha-1) (RY), oilseed radish (Raphanus sativus – 20 kg seeds ha-1) (RD), oilseed radish (10 kg seeds ha-1) + rye (60 kg seeds ha-1) (RD + RY), and oilseed radish (10 kg seeds ha-1) + black oats (60 kg seeds ha-1) (RD + BO). At the beginning of April of each year as of the time the experiment was set up in 2009, cover crops were broadcast on the soil surface. The amount of seeds per hectare used was 50 % of the highest values recommended by Monegat (1991)Monegat C. Manejo de plantas de cobertura do solo em pequenas propriedades. In: Plantas de cobertura do solo: características e manejo em pequenas propriedades. Chapecó: Edição do Autor; 1991. p.146-239.. The proportion of seeds of winter species in the RD + RY and RD + BO treatments was 86 % for cruciferous and 14 % for the grass species. In the summer of each year, all plots were sown with velvet bean (Mucuna aterrima). A randomized block experimental design was used, with eight replicates. Each experimental unit was 5 × 5 m (25 m2).
In July of each year, all winter species and weeds were rolled down with a knife roller. Subsequently, 42 kg ha-1 P2O5 (Gafsa rock phosphate), 104 kg ha-1 K2O and 160 kg ha-1 N in the form of poultry manure were broadcast on the soil surface. Half of the fertilizer was applied at seedling transplanting and half 30 days after transplanting. In the 2011 crop, rock phosphate was not applied because P content was very high according to regional soil fertility parameters (CQFSRS/SC, 2004Comissão de Química e Fertilidade do Solo – RS/SC – CQFSRS/SC. Manual de adubação e de calagem para os estados para os estados do Rio Grande do Sul e Santa Catarina. 10a ed. Porto Alegre: Sociedade Brasileira de Ciência do Solo; 2004.). Seed furrows were opened with a machine adapted for no-tillage, and onion seedlings (cv. EMPASC 352 – Bola Precoce) were manually transplanted. Plants were placed in rows 0.40 m apart, with 0.10 m between plants. Each plot had 10 rows, for a total of 500 onion plants per plot. Weeding was done at 60 and 90 days after seedling transplanting. These procedures were repeated each year.
Cover crop shoot sampling and litter bag preparation and placement on plots
On July 11, 2013, samples were taken of cover crop shoots from each treatment. Plants were cut at ground level in a 0.5 × 0.5 m (0.25 m2) area for each subsample. Five fresh sub-samples were collected in each plot and dried at 65 °C until constant weight, and plant dry matter (DM) yield was measured. On July 22, 2013, shoots of the single and mixed species of winter cover crops remaining in the field were cut at ground level and left in the area. Samples of plant shoot matter were dried at 65 °C. After drying, the DM was ground and set aside for characterization of chemical composition (Table 1). The other part of fresh shoot matter was homogenized, weighed, and placed in nylon fabric litter bags (0.40 × 0.40 m, with 2 mm mesh) (Tagliavini et al., 2007Tagliavini M, Tonon G, Scandellari F, Quiñones A, Palmieri S, Menarbin G, Gioacchini P, Masia A. Nutrient recycling during the decomposition of apple leaves (Malus domestica) and mowed grasses in an orchard. Agric Ecosyst Environ. 2007;118:191-200. doi:10.1016/j.agee.2006.05.018). The following amounts of fresh shoot matter were placed in each litter bag: 235.13 g WD, 767.99 g of BO, 434.50 g of RY, 460.46 g of RD, 570.67 g of RD + BO, and 416.82 g of RD + RY. These values, in terms of DM, amounted to 4620 kg ha-1 of WD, 5263 kg ha-1 of BO, 5060 kg ha-1 of RY, 3640 kg ha-1 of RD, 4030 kg ha-1 of RD + BO, and 3730 kg ha-1 of RD + RY, and are equivalent to DM amounts obtained in the field at the sampling carried out on July 11, 2013. The other initial chemical characteristics of the residues and amounts of nutrients added in each treatment are shown in table 1.
On July 22, 2013, the litter bags were placed directly on the soil surface, between onion rows. Twenty-four bags of each treatment were placed together (six bags in each plot), for a total of 144 litter bags.
Collection of litter bags, plant residue analysis, and soil chemical analysis
The litter bags were collected at time zero (at the time of their placement on the soil surface) and at 30, 45, 60, 75, and 90 days after deposition (DAD). At each collection time, four litter bags per treatment were evaluated. The residues were removed from the bags, rinsed with distilled water, washed with 0.1 mol L-1 HCl solution for 2 min, and rinsed again with distilled water. Immediately after, the residues were dried in a laboratory oven at 65 °C until constant weight, at which time the remaining DM was measured. The residues were then ground and stored for chemical analyses.
Cover crop plant residues were analyzed for lignin, cellulose, and non-structural biomass, which refers to the fraction that does not correspond to the lignin and cellulose (Aber and Martin, 1999Aber JD, Martin M. Leaf chemistry. 1992-1993 (ACCP). Oak Ridge: Oak Ridge National Laboratory; 1999.). Total organic carbon (TOC), N, P, K, Ca, and Mg contents were quantified according to Tedesco et al. (1995)Tedesco MJ, Gianello C, Bissani CA, Bohnen H, Volkweiss SJ. Análises de solo, planta e outros materiais. 2ª. ed. Porto Alegre: UFRGS; 1995..
In July 2012 and 2013, after onion seedling transplanting, and in December of each year after onion harvest, five soil sub-samples from the 0.00-0.10 m layer were collected in each plot and mixed to compose a combined sample. The soil was air-dried, ground, passed through a 2 mm sieve, and analyzed to determine TOC; pH in water (1:1 ratio); Ca, Mg, Al (extracted by 1 mol L-1 KCl); and availables P and K (extracted by Mehlich-1) (Tedesco et al., 1995Tedesco MJ, Gianello C, Bissani CA, Bohnen H, Volkweiss SJ. Análises de solo, planta e outros materiais. 2ª. ed. Porto Alegre: UFRGS; 1995.). Ca and Mg were determined by atomic absorption spectrophotometry. Al was determined by titration with NaOH. Phosphorus was determined by using a visible light spectrophotometer (λ 660 nm) by the molybdenum blue colorimetric method, while K content was determined by flame photometry (Tedesco et al., 1995Tedesco MJ, Gianello C, Bissani CA, Bohnen H, Volkweiss SJ. Análises de solo, planta e outros materiais. 2ª. ed. Porto Alegre: UFRGS; 1995.). Cation exchange capacity at pH 7.0 (CECpH7.0) and percent base saturation (V) were calculated according to CQFSRS/SC (2004).
Onion yield
On November 20, 2012 and November 23, 2013, onions were harvested manually in six central rows of 4-m length in each plot, for a total area of 9.6 m2 (4 × 2.4 m). The bulbs were left on the soil surface for 10 days for curing (loss of water and leaf drying). Subsequently, bulbs were classified according to their diameter (Epagri, 2013Empresa de Pesquisa Agropecuária e Extensão Rural – Epagri. Sistema de produção para a cebola de Santa Catarina. 4ª ed. Florianópolis: 2013.): class 0 (<15 mm), class 1 (>15 and <35 mm), class 2 (>35 and <50 mm), class 3 (>50 and <60 mm), class 4 (>70 and <90 mm) and class 5 (>90 mm), besides weighting rotted and flowered bulbs.
Statistical analysis
Data of DM, cellulose, lignin, non-structural biomass, TOC, N, P, K, Ca, and Mg contents were presented as percentage remaining in each period of time in comparison to their initial amounts. Those results were fitted to the exponential mathematical model described by Wieder and Lang (1982)Wieder RK, Lang GE. A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology. 1982;63:1636-42. doi:10.2307/1940104: X = Xoe(-kt), where X = the amount of DM or nutrient remaining after a period of time t, in days; Xo = initial amount of DM or nutrient; and k = constant of decomposition. Half-life (t½ = 0.693/k) was calculated from the k value (Paul and Clark, 1996Paul EA, Clark FE. Dynamics of residue decomposition and soil organic matter turnover. In: Paul EA, Clark FE, editors. Soil microbiology and biochemistry. 2nd ed. San Diego: Academic; 1996. p.158-79.), which expresses the time required for half of the residues to be decomposed or for half of the nutrients contained in the residues to be released.
The results of the onion yield parameters, soil properties and decomposition parameters (Xo, k, and t½) were analyzed for normality and homogeneity using the Shapiro-Wilk test and subjected to analysis of variance. When effects were significant, averages were compared by the Tukey test at 5 % (SAS, 2003SAS Institute Inc®. SAS versão 9.1.3. Cary: 2003.).
RESULTS AND DISCUSSION
Residue decomposition and nutrient release
The dynamics of DM, TOC, lignin, cellulose, and non-structural biomass loss, as well as N, P, K, Ca, and Mg mineralization, up to 90 DAD were explained by the exponential decay model (Table 2; Figures 2 and 3). That time frame corresponded approximately to the growth period until onion bulb formation. The remaining DM residue in all treatments decreased over time (Table 2; Figure 2). This is due to fragmentation and uptake by soil fauna, and to leaching of water-soluble compounds degraded by the soil microbial population. According to Aita and Giacomini (2003)Aita C, Giacomini S. Decomposição e liberação de nitrogênio de resíduos culturais de plantas de cobertura de solo solteiras e consorciadas. Rev Bras Cienc Solo. 2003;27:601-12. doi:10.1590/S0100-06832003000400004, it is likely that, in addition to microbial decomposition, part of the decreases in DM, TOC, non-structural biomass, N, and K were caused by leaching promoted by rainfall that occurred during the experiment (Figures 1a and 1b).
Parameters fitted (X = Xo e(-kt)) to the values of remaining dry matter (DM), total organic carbon (TOC), lignin (Lig), cellulose (Cel), non-structural biomass (Bio), nitrogen (N), phosphorus (P), potassium (K), calcium (Ca) and magnesium (Mg); decomposition constant (k); and half-life (t½) of each compartment and R2 values for the residues of weeds (WD), black oat (BO), rye (RY), oilseed radish (RD), oilseed radish + black oat (RD + BO), and oilseed radish + rye (RD + RY) deposited on the onion row
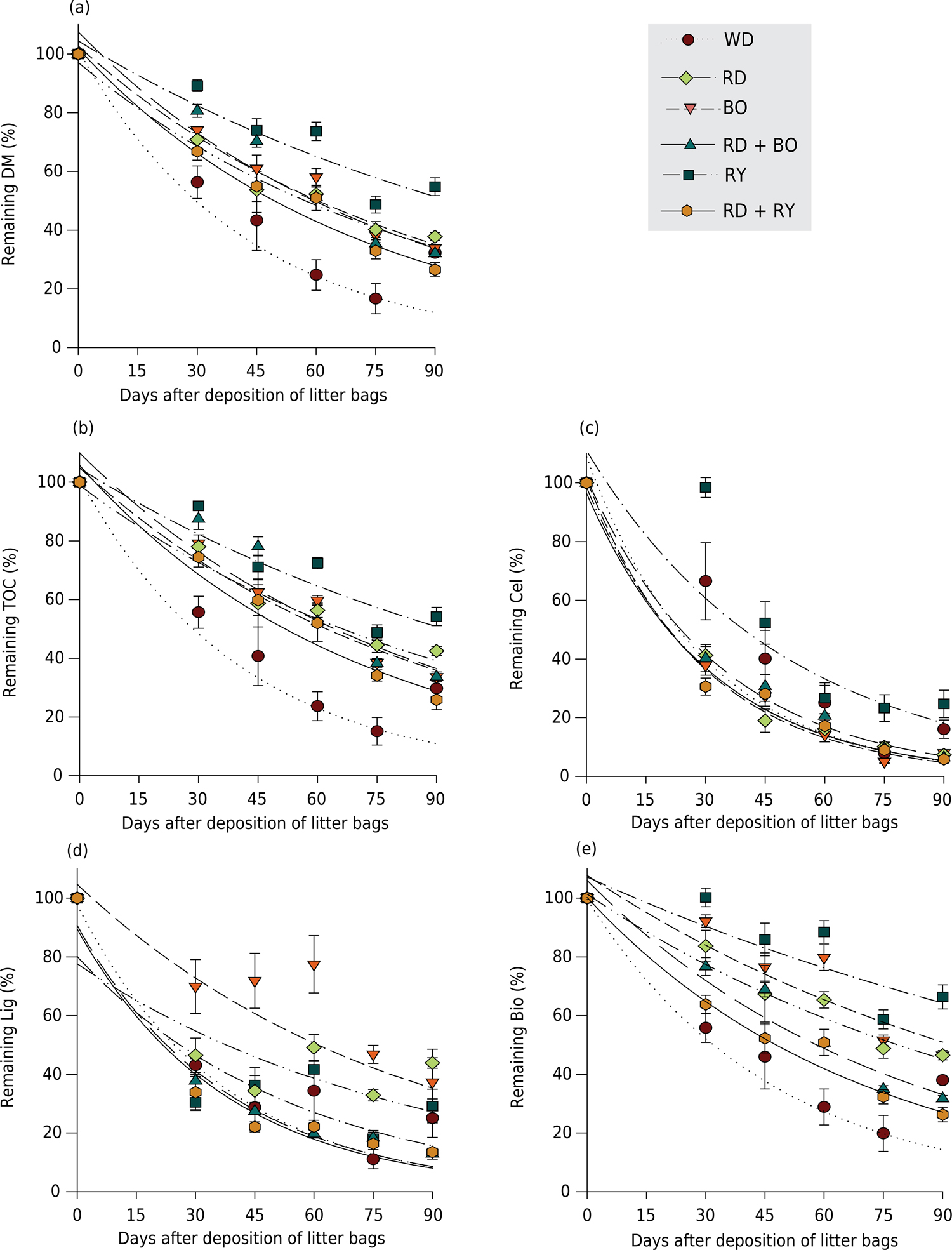
Remaining percentage of dry matter (DM) (a), total organic carbon (TOC) (b), cellulose (Cel) (c), lignin (Lig) (d), and non-structural biomass (Bio) (e) in the residues of weeds (WD), black oat (BO), rye (RY), oilseed radish (RD), oilseed radish + black oat (RD + BO), and oilseed radish + rye (RD + RY) deposited on the onion row for a period of 90 days. Vertical bars represent standard error of the mean.
Remaining percentage of nitrogen (N) (a), phosphorus (P) (b), potassium (K) (c), calcium (Ca) (d), and magnesium (Mg) (e) in the residues of weeds (WD), black oat (BO), rye (RY), oilseed radish (RD), oilseed radish + black oat (RD + BO), and oilseed radish + rye (RD + RY) deposited on the onion crop row for a period of 90 days. Vertical bars represent standard error of the mean.
Soil microorganisms, especially fungi and bacteria, colonize plant residues; part of their TOC is used during decomposition and part of it is released in the form of CO2 into the atmosphere (Manzoni et al., 2008Manzoni S, Jackson RB, Trofymow JA, Porporato A. The global stoichiometry of litter nitrogen mineralization. Science. 2008;321:684-6. doi:10.1126/science.1159792). In addition, the increase in air temperature, and consequently in soil temperature, as well as rainfall, that occurred during the experiment may have contributed to increase breakdown of residues by microorganisms (Figures 1a and 1b) (Teixeira et al., 2011Teixeira MB, Loss A, Pereira MG, Pimentel C. Decomposição e liberação de nutrientes da parte aérea de plantas de milheto e sorgo. Rev Bras Cienc Solo. 2011;35:867-76. doi:10.1590/S0100-06832011000300021, 2012Teixeira MB, Loss A, Pereira MG, Pimentel C. Decomposição e ciclagem de nutrientes dos resíduos de quatro plantas de cobertura do solo. Idesia. 2012;30:55-64. doi:10.4067/S0718-34292012000100007).
From 30 to 90 DAD, the highest values of remaining DM were observed in the RY residue, with a half-life (t½) of 90 days (Table 2; Figure 2a). Intermediate percentages of DM were observed from 45 to 90 DAD in residues of RD, BO, and RD+BO, with t½ of 61, 58, and 54 days, respectively (Table 2; Figure 2a). The lowest percentage of remaining DM, with a t½ of 30 days, was observed in the WD residue. The larger amount of DM of RY remaining at 90 DAD may be attributed to its high initial lignin content (102.81 g kg-1) and C/N ratio value (22), and its low Cel/Lig ratios (3) (Table 1). Residues with high levels of lignin, and consequently higher Lig/N ratio values and lower Cel/Lig values, have slower decomposition, since lignin protects cell wall cellulose from microbial degradation (Berg and McClaugherty, 2008Berg B, Mcclaugherty C. Plant Litter: decomposition, humus formation, carbon sequestration. 2nd. ed. Heidelberg: Springer-Verlag; 2008.). Typically, decomposition of plant residues placed on the soil surface is determined in part by their C/N ratio, as well as their Lig/N and Cel/Lig ratios (Manzoni et al., 2008Manzoni S, Jackson RB, Trofymow JA, Porporato A. The global stoichiometry of litter nitrogen mineralization. Science. 2008;321:684-6. doi:10.1126/science.1159792; Gentile et al., 2009Gentile R, Vanlauwe B, Kessel CV, Six J. Managing N availability and losses by combining fertilizer-N with different quality residues in Kenya. Agric Ecosyst Environ. 2009;131:308-14. doi:10.1016/j.agee.2009.02.003). Residues with a C/N ratio below 30, as in WD (16), are more readily colonized by soil microbial populations than those with a C/N ratio above 30, since there is more available N to incorporate in their tissues. That increases mineralization of residue components and, therefore, results in a smaller amount of remaining DM (Ferreira et al., 2014Ferreira PAA, Girotto E, Trentin G, Miotto A, Melo GW, Ceretta CA, Kaminski J, Frari BKD, Marchezan C, Silva L, Faversani JC, Brunetto G. Biomass decomposition and nutrient release from black oat and hairy vetch residues deposited in a vineyard. Rev Bras Cienc Solo. 2014;38:1621-32. doi:10.1590/S0100-06832014000500027). The smaller amount of remaining DM of WD from 30 to 90 DAD (Table 3) is due in part to its C/N ratio, which was lower due to the mixture of several species, including grasses and leguminous plants, and to its Lig/N and Cel/Lig ratio values.
The remaining TOC decreased over time in the residues of all treatments (Table 2; Figure 2b). This may be attributed to leaching of soluble organic C, which can be readily used by the microbial population (Carneiro et al., 2008Carneiro MAC, Cordeiro MAS, Assis PCR, Moraes ES, Pereira HS, Paulino HB, Souza ED. Produção de fitomassa de diferentes espécies de cobertura e suas alterações na atividade microbiana de solo de cerrado. Bragantia. 2008;67:455-62. doi:10.1590/S0006-87052008000200021). Part of that C may be released into the atmosphere as CO2 (Carneiro et al., 2008Carneiro MAC, Cordeiro MAS, Assis PCR, Moraes ES, Pereira HS, Paulino HB, Souza ED. Produção de fitomassa de diferentes espécies de cobertura e suas alterações na atividade microbiana de solo de cerrado. Bragantia. 2008;67:455-62. doi:10.1590/S0006-87052008000200021; Manzoni et al., 2008Manzoni S, Jackson RB, Trofymow JA, Porporato A. The global stoichiometry of litter nitrogen mineralization. Science. 2008;321:684-6. doi:10.1126/science.1159792), and degradation of recalcitrant organic compounds from the residues may also contribute to that effect (Berg and McClaugherty, 2008Berg B, Mcclaugherty C. Plant Litter: decomposition, humus formation, carbon sequestration. 2nd. ed. Heidelberg: Springer-Verlag; 2008.). The highest percentage of remaining TOC from 30 to 90 DAD was observed in the RY residue, followed by RD, with t½ of 88 and 68 days, respectively (Table 2; Figure 2b). Intermediate TOC percentages were recorded from 45 to 90 DAD in residues of BO and RD + BO, with t½ of 58 and 57 days, respectively. The lowest percentage of remaining TOC as of 30 DAD was observed in the WD residues with a t½ of 29 days. The results reported for RY are due to their high lignin content and high C/N and Lig/N ratios. The RY species therefore decomposes and releases nutrients to the soil more slowly and gradually (Giacomini et al., 2003Giacomini SJ, Aita C, Hubner AP, Lunkes A, Guidini E, Amaral E. Liberação de fósforo e potássio durante a decomposição de resíduos culturais em plantio direto. Pesq Agropec Bras. 2003;38:1097-104. doi:10.1590/S0100-204X2003000900011) (Table 1). In contrast, the lower TOC percentages recorded in WD are the result of their low lignin contents and, hence, high cellulose contents, and lower C/N and Lig/N ratio values, which promotes faster decomposition and nutrient release (Crusciol et al., 2005Crusciol CAC, Cottica RL, Lima EV, Andreotti M, Moro E, Marcon E. Persistência de palhada e liberação de nutrientes do nabo-forrageiro no plantio direto. Pesq Agropec Bras. 2005;40:161-8. doi:10.1590/S0100-204X2005000200009). The intermediate TOC values recorded in the RD + BO intercrop are due, in part, to the intermediate contents of lignin (93.51 g kg-1), cellulose (220.69 g kg-1), non-structural biomass (685, 57 g kg-1), N, and the C/N, C/P, and Lig/N ratios, which result in intermediate decomposition and mineralization rates, therefore maintaining adequate amounts of plant residues on the soil surface (Giacomini et al., 2003Giacomini SJ, Aita C, Hubner AP, Lunkes A, Guidini E, Amaral E. Liberação de fósforo e potássio durante a decomposição de resíduos culturais em plantio direto. Pesq Agropec Bras. 2003;38:1097-104. doi:10.1590/S0100-204X2003000900011).
Percentages of lignin, cellulose, and non-structural biomass decreased over time (Table 2, Figures 2c, 2d, and 2e). From 30 to 90 DAD, the highest percentage of remaining cellulose, with a t½ of 35 days up to 90 DAD, was observed in the RY residue (Table 2; Figure 2c). Residues of WD, RD+BO, RD+RY, RD, and BO had lower remaining cellulose percentages in all samplings, with t½ of 24, 23, 22, 22, and 20 days, respectively. Cellulose, an insoluble component mainly found in the primary plant cell wall, is one of the first components to decompose (Berg and McClaugherty, 2008Berg B, Mcclaugherty C. Plant Litter: decomposition, humus formation, carbon sequestration. 2nd. ed. Heidelberg: Springer-Verlag; 2008.). The smallest initial amount of cellulose at the flowering stage, when cover crops are rolled down, was in the RY residues (161.81 g kg-1) (Table 1). That period precedes plant flowering, when rye plants were cut. This may partially explain the pattern presented by RY DM, i.e., slower decomposition rates in comparison to all other treatments. The DM of BO had the highest initial cellulose content (239.59 g kg-1), that partially explains the rapid decomposition of remaining cellulose (Table 1).
Black oats residues had the highest percentage of remaining lignin from 30 to 90 DAD, with a t½ of 74 days (Figure 2d). Intermediate amounts of remaining lignin were observed in the RD and RY residues from 45 to 90 DAD, with t½ of 67 and 40 days, respectively (Table 2). The smallest percentages of remaining lignin were observed in RD + BO, RD + RY, and WD residues, with t½ of 26, 26, and 28 days, respectively. Lignin, found mainly in the secondary cell wall and in the middle lamella, is a major component of plant fibers and a structural component that is highly resistant to microbial decomposition (Berg and McClaugherty, 2008Berg B, Mcclaugherty C. Plant Litter: decomposition, humus formation, carbon sequestration. 2nd. ed. Heidelberg: Springer-Verlag; 2008.; Talbot et al., 2011Talbot JM, Yelle DJ, Nowick J, Treseder KK. Litter decay rates are determined by lignin chemistry. Biogeochemistry. 2011;108:279-5. doi:10.1007/s10533-011-9599-6). There was more remaining lignin in BO and RY residues (Figure 2d), which resulted, in part, in lower decomposition since lignin acts mechanically to protect cellulose (Talbot et al., 2011Talbot JM, Yelle DJ, Nowick J, Treseder KK. Litter decay rates are determined by lignin chemistry. Biogeochemistry. 2011;108:279-5. doi:10.1007/s10533-011-9599-6). The smaller amount of remaining lignin in the RD residue at 90 DAD may be attributed to its low initial lignin content (71.68 g kg-1) and high cellulose content (211.05 g kg-1), which intensified decomposition (Table 1).
Non-structural biomass corresponds to components that do not participate in tissue structure as lignin and cellulose do in the plant cell wall (Tagliavini et al., 2007Tagliavini M, Tonon G, Scandellari F, Quiñones A, Palmieri S, Menarbin G, Gioacchini P, Masia A. Nutrient recycling during the decomposition of apple leaves (Malus domestica) and mowed grasses in an orchard. Agric Ecosyst Environ. 2007;118:191-200. doi:10.1016/j.agee.2006.05.018). The highest percentage of remaining non-structural biomass from 30 to 90 DAD was observed in the RY residue with a t½ of 128 days (Table 2; Figure 2e). Intermediate values were found in the RD + BO and RD + RY residues with t½ of 54 and 48 days, respectively. However, the lowest percentage of remaining non-structural biomass as of 30 DAD was observed in the WD residue, averaging 38 % up to DAD, with a t½ of 34 days. The higher percentage of non-structural biomass in the RY residue can be attributed to its high lignin content (102.81 g kg-1) and low cellulose content (161.81 g kg-1) and to its high C/N ratio (22). The intermediate percentages found in RD + BO and RD + RY residues may be due to intermediate contents of lignin (93.51 and 80.59 g kg-1, respectively) and cellulose (220.69 and 192.91 g kg-1, respectively), and to their C/N ratios (17 and 15 g kg-1, respectively). The lower percentages of remaining non-structural biomass in the WD residue can be associated with its low C/N ratio values (Table 3).
The highest percentage in remaining N from 30 to 90 DAD was observed in RY residue with a t½ of 84 (Figure 3a), while intermediate percentages were observed in RD and RD + BO residues, with t½ of 52 and 42 days, respectively (Table 2). The lowest percentage of remaining N up to 90 DAD was observed in RD + RY and WD residues, with t½ of 28 and 30 days, respectively. The low mineralization rate observed in the RY residues may be due in part to their C/N (22) and Lig/N (6) ratios, low initial N concentration (18.00 g kg-1), high lignin content (102.81 g kg-1), and low cellulose content (161.81 g kg-1), compared to other cover crop species (Table 1). Doneda et al. (2012)Doneda A, Aita C, Giacomini SJ, Miola ECC, Giacomini DA, Schirmann J, Gonzatto R. Fitomassa e decomposição de resíduos de plantas de cobertura puras e consorciadas. Rev Bras Cienc Solo. 2012;36:1714-23. doi:10.1590/S0100-06832012000600005 evaluated the C/N ratio in rye, black oat, radish, and hairy vetch in a NTS in an Udox (Latossolo Vermelho), and found that rye with a C/N ratio of 34, higher than that of other cover crops, had lower plant residue decomposition on the soil surface. While the smaller amount of remaining N in WD can be attributed in part to its low C/N ratio from 30 at 90 DAD (Table 3), the intermediate remaining N levels for the RD + BO intercrop are mainly due to its C/N ratio (17), which resulted in intermediate N release in relation to the other cover crops (Giacomini et al., 2003Giacomini SJ, Aita C, Hubner AP, Lunkes A, Guidini E, Amaral E. Liberação de fósforo e potássio durante a decomposição de resíduos culturais em plantio direto. Pesq Agropec Bras. 2003;38:1097-104. doi:10.1590/S0100-204X2003000900011).
The highest rates of remaining P occurred in the RY residue, and intermediate values were observed in the BO residue with t½ of 39 and 34 days, respectively (Table 2; Figure 3b). The lowest percentage of remaining P was observed in the RD residue with a t½ of 26, which did not differ statistically from the percentage of WD, RD + BO, and RD + RY, with t½ of 28, 28, and 29, respectively (Table 2; Figure 3b). At 90 DAD, the cover crop residues on the soil surface showed remaining P rates lower than 40 %, indicating the release of more than half of the P contained in the residues. The high P retention in RY residues may be due to the predominance of organic P forms, which depend on DM mineralization for their release, and RY had the highest t½ (39 days), as well as the highest lignin levels. The lower content of remaining P in the RD residue can be explained by its low C/P ratio (56) (Table 1) (Giacomini et al., 2003Giacomini SJ, Aita C, Hubner AP, Lunkes A, Guidini E, Amaral E. Liberação de fósforo e potássio durante a decomposição de resíduos culturais em plantio direto. Pesq Agropec Bras. 2003;38:1097-104. doi:10.1590/S0100-204X2003000900011; Ferreira et al., 2014Ferreira PAA, Girotto E, Trentin G, Miotto A, Melo GW, Ceretta CA, Kaminski J, Frari BKD, Marchezan C, Silva L, Faversani JC, Brunetto G. Biomass decomposition and nutrient release from black oat and hairy vetch residues deposited in a vineyard. Rev Bras Cienc Solo. 2014;38:1621-32. doi:10.1590/S0100-06832014000500027). The intermediate values of remaining P for BO are partly due to their C/P ratio (62), which resulted in an intermediate value in comparison to all other species (Table 1). Unlike N, P showed much faster initial release (Figures 3a and 3b), an effect possibly caused by the release of soluble P accumulated in the vacuoles of the plant tissues (Buchanan and King, 1993Buchanan M, King L. Carbon and phosphorus losses from decomposing crop residues in no till and conventional till agroecosystems. Agron J. 1993;85:631-8. doi:10.2134/agronj1993.00021962008500030021x).
The highest percentage of remaining K from 30 to 90 DAD was observed in the RY residue with a t½ of 17 days up to 90 DAD (Table 3; Figure 3c). A lower percentage of remaining K was observed in all other treatments, with an average t½ of 12 days (Table 2; Figure 3c). The higher remaining K content in the RY residue can be explained by its high t½ (90 days) of DM (Table 2), and the high content of non-structural biomass (735.48 g kg-1) (Table 1). The low contents of remaining K at 30 DAD (Figure 3c) are due to the high solubility of this element in the tissue, since it does not bind to any structural component, which facilitates its quick release (Pavinato and Rosolem, 2008Pavinato PS, Rosolem CA. Disponibilidade de nutrientes no solo – decomposição e liberação de compostos orgânicos de resíduos vegetais. Rev Bras Cienc Solo. 2008;32:911-20. doi:10.1590/S0100-06832008000300001) and high content in the soil (Teixeira et al., 2012Teixeira MB, Loss A, Pereira MG, Pimentel C. Decomposição e ciclagem de nutrientes dos resíduos de quatro plantas de cobertura do solo. Idesia. 2012;30:55-64. doi:10.4067/S0718-34292012000100007; Ferreira et al., 2014Ferreira PAA, Girotto E, Trentin G, Miotto A, Melo GW, Ceretta CA, Kaminski J, Frari BKD, Marchezan C, Silva L, Faversani JC, Brunetto G. Biomass decomposition and nutrient release from black oat and hairy vetch residues deposited in a vineyard. Rev Bras Cienc Solo. 2014;38:1621-32. doi:10.1590/S0100-06832014000500027).
The highest percentage of remaining Ca from 45 to 90 DAD was observed in the BO residue, followed by the RY residue with t½ of 50 and 36 days, respectively (Table 2; Figure 3d). Lower values of remaining Ca were observed in the residues of RD + BO, WD, RD, and RD + RY, with t½ of 24, 24, 26, and 26 days, respectively (Table 2; Figure 3d). The high remaining Ca in the BO residue is related to the high remaining lignin t½ (74 days) in those residues (Table 2). This occurred because Ca is part of the structural composition of the cell, acts as a cofactor in enzymes involved in ATP and phospholipid hydrolysis, and is a second messenger in metabolic regulation (Taiz and Zeiger, 2013Taiz L, Zeiger E. Fisiologia vegetal. 5ª. ed. Porto Alegre: Artmed; 2013.). That results in an additional difficulty to its release from decomposing residues into the soil (Teixeira et al., 2011Teixeira MB, Loss A, Pereira MG, Pimentel C. Decomposição e liberação de nutrientes da parte aérea de plantas de milheto e sorgo. Rev Bras Cienc Solo. 2011;35:867-76. doi:10.1590/S0100-06832011000300021). The smallest remaining Ca content in the WD residue is due in part to a lower t½ (34 days) in non-structural biomass (Table 2) and the low remaining C/N ratio (Table 3), which increased decomposition and thus intensified Ca release.
The highest percentage of remaining Mg from 30 to 90 DAD was observed in the RY residue, followed by BO, with t½ of 88 and 77 days, respectively (Table 2; Figure 3e). Intermediate values were found in RD residues, with a t½ of 52 days. Lower percentages of remaining Mg were observed in the residues of RD + BO, WD, and RD + RY, with t½ of 26, 30, and 35 days, respectively (Table 2; Figure 3e). Mg is the main component of chlorophyll and has a specific role in plant cells, especially in photosynthesis and in the activation of enzymes involved in respiration (Taiz and Zeiger, 2013Taiz L, Zeiger E. Fisiologia vegetal. 5ª. ed. Porto Alegre: Artmed; 2013.). The intermediate Mg value recorded in RD may be due in part to its intermediate content in DM (2.11 g kg1), which led to a mean initial addition of 7.7 kg ha-1 of this element (Table 1). The lower remaining Mg contents in WD are due in part to its C/N ratios of 17.0, 17.1, 14.5, 14.3, and 16.4 at 30, 45, 60, 75, and 90 DAD, respectively, which are low in comparison to all other treatments (Table 4). Magnesium is part of ionic compounds and soluble molecules, which facilitates its release, and this is enhanced by the low C/N ratio of the remaining DM (Oliveira et al., 1999Oliveira MW, Trivelin PCO, Penatti CP, Piccolo MC. Decomposição de nutrientes da palhada de cana-de-açúcar em campo. Pesq Agropec Bras. 1999;34:2359-62. doi:10.1590/S0103-90161999000400006).
Onion bulb yield for commercial classes 2 and 3, rotted bulbs/flowering bulbs, and total yield in the 2012 and 2013 crop seasons under the management practice of cover crops with residues of weeds (WD), black oat (BO), rye (RY), oilseed radish (RD), oilseed radish + black oat (RD + BO), and oilseed radish + rye (RD + RY) deposited on the onion row
Onion Yield
In the 2012 crop, the highest and lowest overall onion yields were observed in the soil with deposition of BO and WD residues, respectively (Table 4). In 2013, higher total onion yield was observed in soil with cover crop residues, regardless of the plant species. The lower onion yield in WD in both crops can be explained by the presence of weeds, which compete with onion for water and nutrients (Souza et al., 2013Souza M, Comin JJ, Leguizamón ES, Kurtz C, Brunetto G, Müller Júnior V, Ventura B, Camargo AP. Matéria seca de plantas de cobertura, produção de cebola e atributos químicos do solo em sistema plantio direto agroecológico. Cienc Rural. 2013;43:21-7. doi:10.1590/S0103-84782012005000150) and which have a more accelerated decomposition of DM in comparison to cover crops (Tables 2 and 3, Figure 2a). Furthermore, this results in faster release of nutrients to the soil, promoting higher losses of these elements. Lower onion yield and smaller bulb size in plots with weeds were also found by Wang et al. (2008)Wang G, Ngouajio M, Warncke DD. Nutrient cycling, weed suppression, and onion yield following brassica and sorghum sudangrass cover crops. HortTechnology. 2008;18:68-74. doi:10.1007/s13165-010-0002-z.
Bulb classes 2 and 3 were found in the 2012 and 2013 crops (Table 4). Class 3 bulb yield did not differ among treatments, while higher yield in class 2 bulbs was found in soil with cover crop residues. In 2012, rotted and flowering bulbs were found in all plots, with no differences among treatments. In the 2013 crop, rotted and flowering bulbs were also found in all treatments, and the rotted and flowering bulbs was observed in the treatment with BO residues. The higher yield of onion and class 2 bulbs in the 2012 and 2013 crops in all treatments with cover crops compared to plots with WD can be explained by increased DM yield (Figure 2a), which increases protection of the soil surface against the impact of rain drops and consequently reduces the loss of soil, water, and nutrients by runoff (Bertol et al., 2011Bertol I, Gobbi E, Barbosa FT, Paz-Ferreiro J, Gebler L, Ramos JC, Werner RS. Erosão hídrica em campo nativo sob diversos manejos: perdas de água e solo e de fósforo, potássio e amônio na água de enxurrada. Rev Bras Cienc Solo. 2011;35:1421-30. doi:10.1590/S0100-06832011000400036). This increased yield may also be related to the amounts of nutrients added (Table 1) and released during the decomposition of DM (Figures 2 and 3), because part of these released nutrients were able to be taken up by onion throughout the 90 DAD period (Souza et al., 2013Souza M, Comin JJ, Leguizamón ES, Kurtz C, Brunetto G, Müller Júnior V, Ventura B, Camargo AP. Matéria seca de plantas de cobertura, produção de cebola e atributos químicos do solo em sistema plantio direto agroecológico. Cienc Rural. 2013;43:21-7. doi:10.1590/S0103-84782012005000150).
Soil chemical properties
In the 2012 crop, organic matter (OM), pH in water, Ca2+, Mg2+, and Al3+, and available P, CECpH7.0, and base saturation (V) in the 0.00-0.10 m layer were not affected by cover crop residue decomposition, except for available K contents, which were higher in soil with WD (Table 5). The higher contents of available K in soil with WD reflect the ability of weed species in nutrient cycling, together with faster decomposition of their residues.
Organic matter content (OM), pH in water, exchangeable calcium, magnesium and aluminum, available potassium and phosphorus, and CECpH7.0 and base saturation (V) in the 0.00-0.10 m layer of an Inceptisol cropped to onion, in the presence of residues of weeds (WD), black oat (BO), rye (RY), oilseed radish (RD), oilseed radish + black oat (RD + BO), and oilseed radish + rye (RD + RY) deposited on the onion row
In the 2012 crop, after the onion harvest, OM, Al3+, P, K, and CECpH7.0 in the 0.00-0.10 m layer were not affected by the presence of residues (Table 5). However, higher values of pH in water, Ca and Mg contents, and base saturation (V) after onion harvest were found in plots with WD, RD, and RD + BO residues compared to plots with BO (Table 5). This may be due to increased Ca and Mg uptake and accumulation by BO throughout its cycle (Figures 3d and 3e), which may have promoted high onion yield in the 2012 crop (Table 4). The high soil pH in the 2012 crop can be attributed to the anionic characteristic of the residues, with consequent complexation of H+ and Al3+ ions in the soil (Pavinato and Rosolem, 2008Pavinato PS, Rosolem CA. Disponibilidade de nutrientes no solo – decomposição e liberação de compostos orgânicos de resíduos vegetais. Rev Bras Cienc Solo. 2008;32:911-20. doi:10.1590/S0100-06832008000300001). In the 2013 crop, the contents of OM, pH in water, Ca2+, Mg2+, Al3+, P, K, CECpH7.0, and V in the 0.00-0.10 m layer before seedling transplanting and after onion harvest were not affected by residue decomposition (Table 5).
CONCLUSIONS
The rye plant residues presented the highest half-life and they released less nutrients until 90 DAD. The great permanence of rye residue was considered a protection to soil surface, the opposite was observed with spontaneous vegetation.
The cultivation and addition of dry residue of cover crops increased the onion yield at 2.5 Mg ha-1.
ACKNOWLEDGMENTS
We thank the CNPq (MCTI/MAPA/MDA/MEC/MPA/CNPq N° 81/2013 and the MCTI/CNPq N° 14/2014) for financial support and for research productivity scholarships, and the CAPES for scholarships.
REFERENCES
- Aber JD, Martin M. Leaf chemistry. 1992-1993 (ACCP). Oak Ridge: Oak Ridge National Laboratory; 1999.
- Aita C, Giacomini S. Decomposição e liberação de nitrogênio de resíduos culturais de plantas de cobertura de solo solteiras e consorciadas. Rev Bras Cienc Solo. 2003;27:601-12. doi:10.1590/S0100-06832003000400004
- Berg B, Mcclaugherty C. Plant Litter: decomposition, humus formation, carbon sequestration. 2nd. ed. Heidelberg: Springer-Verlag; 2008.
- Bertol I, Gobbi E, Barbosa FT, Paz-Ferreiro J, Gebler L, Ramos JC, Werner RS. Erosão hídrica em campo nativo sob diversos manejos: perdas de água e solo e de fósforo, potássio e amônio na água de enxurrada. Rev Bras Cienc Solo. 2011;35:1421-30. doi:10.1590/S0100-06832011000400036
- Brunetto G, Ceretta CA, Melo GWB, Kaminski J, Trentin G, Girotto E, Ferreira PAA, Miotto A, Trivelin PCO. Contribution of nitrogen from agricultural residues of rye to “Niagara Rosada” grape nutrition. Sci Hortic. 2014;169:66-70. doi:10.1016/j.scienta.2014.02.019
- Buchanan M, King L. Carbon and phosphorus losses from decomposing crop residues in no till and conventional till agroecosystems. Agron J. 1993;85:631-8. doi:10.2134/agronj1993.00021962008500030021x
- Carneiro MAC, Cordeiro MAS, Assis PCR, Moraes ES, Pereira HS, Paulino HB, Souza ED. Produção de fitomassa de diferentes espécies de cobertura e suas alterações na atividade microbiana de solo de cerrado. Bragantia. 2008;67:455-62. doi:10.1590/S0006-87052008000200021
- Castro NEA, Silva MLN, Freitas DAF, Carvalho GJ, Marques RM, Gontijo Neto GF. Plantas de cobertura no controle da erosão hídrica sob chuvas naturais. Biosci J. 2011;27:775-85.
- Chen B, Liu E, Tian Q, Yan C, Zhang Y. Soil nitrogen dynamics and crop residues. A review. Agron Sust Dev. 2014;34:429-42. doi:10.1007/s13593-014-0207-8
- Claessen MEC, organizador. Manual de métodos de análise de solo. 2ª ed. Rio de Janeiro: Centro Nacional de Pesquisa de Solos; 1997.
- Comissão de Química e Fertilidade do Solo – RS/SC – CQFSRS/SC. Manual de adubação e de calagem para os estados para os estados do Rio Grande do Sul e Santa Catarina. 10a ed. Porto Alegre: Sociedade Brasileira de Ciência do Solo; 2004.
- Cornwell WK, Cornelissen JHC, Amatangelo K, Dorrepaal E, Eviner VT, Godoy O, Hobbie SE, Hoorens B, Kurokawa H, Pérez-Harguindeguy N, Quested HM, Santiago LS, Wardle D, Wright IJ, Aerts R, Allison SD, van Bodegom P, Brovkin V, Chatain A, Callaghan TV, Díaz S, Garnier E, Gurvich D, Kazakou E, Klein JA, Read J, Reich PB, Soudzilovskaia NA, Vaieretti MV, Westoby M. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol Lett. 2008;11:1065-71. doi:10.1111/j.1461-0248.2008.01219.x
- Crusciol CAC, Cottica RL, Lima EV, Andreotti M, Moro E, Marcon E. Persistência de palhada e liberação de nutrientes do nabo-forrageiro no plantio direto. Pesq Agropec Bras. 2005;40:161-8. doi:10.1590/S0100-204X2005000200009
- Doneda A, Aita C, Giacomini SJ, Miola ECC, Giacomini DA, Schirmann J, Gonzatto R. Fitomassa e decomposição de resíduos de plantas de cobertura puras e consorciadas. Rev Bras Cienc Solo. 2012;36:1714-23. doi:10.1590/S0100-06832012000600005
- Empresa de Pesquisa Agropecuária e Extensão Rural – Epagri. Sistema de produção para a cebola de Santa Catarina. 4ª ed. Florianópolis: 2013.
- Ferreira PAA, Girotto E, Trentin G, Miotto A, Melo GW, Ceretta CA, Kaminski J, Frari BKD, Marchezan C, Silva L, Faversani JC, Brunetto G. Biomass decomposition and nutrient release from black oat and hairy vetch residues deposited in a vineyard. Rev Bras Cienc Solo. 2014;38:1621-32. doi:10.1590/S0100-06832014000500027
- Gentile R, Vanlauwe B, Kessel CV, Six J. Managing N availability and losses by combining fertilizer-N with different quality residues in Kenya. Agric Ecosyst Environ. 2009;131:308-14. doi:10.1016/j.agee.2009.02.003
- Giacomini SJ, Aita C, Hubner AP, Lunkes A, Guidini E, Amaral E. Liberação de fósforo e potássio durante a decomposição de resíduos culturais em plantio direto. Pesq Agropec Bras. 2003;38:1097-104. doi:10.1590/S0100-204X2003000900011
- Kottek M, Grieser J, Beck C, Rudolf B, Rubel F. World map of the Köppen-Geiger climate classification updated. Meteorol Z. 2006;15:259-63. doi:10.1127/0941-2948/2006/0130
- Manzoni S, Jackson RB, Trofymow JA, Porporato A. The global stoichiometry of litter nitrogen mineralization. Science. 2008;321:684-6. doi:10.1126/science.1159792
- Martins RP, Comin JJ, Gatiboni LC, Roberto C, Soares F, Couto R, Brunetto G. Mineralização do nitrogênio de plantas de cobertura, solteiras e consorciadas, depositadas sobre um solo com histórico de cultivo de cebola. Rev Ceres. 2014;61:587-96. doi:10.1590/0034-737X201461040020
- Monegat C. Manejo de plantas de cobertura do solo em pequenas propriedades. In: Plantas de cobertura do solo: características e manejo em pequenas propriedades. Chapecó: Edição do Autor; 1991. p.146-239.
- Moraes PVD, Agostinetto D, Panozzo LE, Oliveira C, Vignolo GK, Markus C. Manejo de plantas de cobertura no controle de plantas daninhas e desempenho produtivo da cultura do milho. Semina: Cienc Agrár. 2013;34:497-508. doi:10.5433/1679-0359.2013v34n2p497
- Oliveira MW, Trivelin PCO, Penatti CP, Piccolo MC. Decomposição de nutrientes da palhada de cana-de-açúcar em campo. Pesq Agropec Bras. 1999;34:2359-62. doi:10.1590/S0103-90161999000400006
- Paul EA, Clark FE. Dynamics of residue decomposition and soil organic matter turnover. In: Paul EA, Clark FE, editors. Soil microbiology and biochemistry. 2nd ed. San Diego: Academic; 1996. p.158-79.
- Pavinato PS, Rosolem CA. Disponibilidade de nutrientes no solo – decomposição e liberação de compostos orgânicos de resíduos vegetais. Rev Bras Cienc Solo. 2008;32:911-20. doi:10.1590/S0100-06832008000300001
- SAS Institute Inc®. SAS versão 9.1.3. Cary: 2003.
- Soil Survey Staff. Soil Taxonomy: a basic system of soil classification for making and interpreting soil surveys. 2nd ed. Washington, DC: US Government Printing Office: 2006.
- Souza M, Comin JJ, Leguizamón ES, Kurtz C, Brunetto G, Müller Júnior V, Ventura B, Camargo AP. Matéria seca de plantas de cobertura, produção de cebola e atributos químicos do solo em sistema plantio direto agroecológico. Cienc Rural. 2013;43:21-7. doi:10.1590/S0103-84782012005000150
- Tagliavini M, Tonon G, Scandellari F, Quiñones A, Palmieri S, Menarbin G, Gioacchini P, Masia A. Nutrient recycling during the decomposition of apple leaves (Malus domestica) and mowed grasses in an orchard. Agric Ecosyst Environ. 2007;118:191-200. doi:10.1016/j.agee.2006.05.018
- Taiz L, Zeiger E. Fisiologia vegetal. 5ª. ed. Porto Alegre: Artmed; 2013.
- Talbot JM, Yelle DJ, Nowick J, Treseder KK. Litter decay rates are determined by lignin chemistry. Biogeochemistry. 2011;108:279-5. doi:10.1007/s10533-011-9599-6
- Tedesco MJ, Gianello C, Bissani CA, Bohnen H, Volkweiss SJ. Análises de solo, planta e outros materiais. 2ª. ed. Porto Alegre: UFRGS; 1995.
- Teixeira MB, Loss A, Pereira MG, Pimentel C. Decomposição e ciclagem de nutrientes dos resíduos de quatro plantas de cobertura do solo. Idesia. 2012;30:55-64. doi:10.4067/S0718-34292012000100007
- Teixeira MB, Loss A, Pereira MG, Pimentel C. Decomposição e liberação de nutrientes da parte aérea de plantas de milheto e sorgo. Rev Bras Cienc Solo. 2011;35:867-76. doi:10.1590/S0100-06832011000300021
- Trinsoutrot I, Recous S, Bentz B, Linères M, Chèneby D, Nicolardot B. Biochemical quality of crop residues and carbon and nitrogen mineralization kinetics under nonlimiting nitrogen conditions. Soil Sci Soc Am J. 2000;64:918-26. doi:10.2136/sssaj2000.643918x
- Wang G, Ngouajio M, Warncke DD. Nutrient cycling, weed suppression, and onion yield following brassica and sorghum sudangrass cover crops. HortTechnology. 2008;18:68-74. doi:10.1007/s13165-010-0002-z
- Wieder RK, Lang GE. A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology. 1982;63:1636-42. doi:10.2307/1940104
Publication Dates
-
Publication in this collection
2016
History
-
Received
19 Feb 2015 -
Accepted
22 Dec 2015