Resumos
O objetivo deste trabalho foi simular o aparecimento de folhas em genótipos de arroz cultivados e em biótipos de arroz-vermelho por meio da adaptação do modelo de Wang e Engel, modificado por Streck et al. Dois experimentos foram conduzidos em Santa Maria, RS, em 2004/2005 e 2005/2006. Foram utilizadas as cultivares IRGA 417 e EEA 406, um híbrido e dois biótipos de arroz-vermelho. O modelo de aparecimento de folhas foi ajustado para os genótipos usando-se dados de estágio de Haun (HS), de cinco épocas de semeadura, em 2004/2005, e a avaliação do modelo foi feita com dados de HS medidos em três épocas de semeadura, em 2005/2006. Realizou-se também um experimento numérico utilizando-se dados de temperatura de 2003/2004 e de 2006/2007, e comparou-se o HS simulado dos genótipos cultivados em relação aos biótipos de arroz-vermelho. O modelo apresentou bom desempenho para simular o HS com raiz do quadrado médio do erro geralmente menor do que uma folha. A emissão de folhas na haste principal foi maior nos genótipos modernos - IRGA 417 e híbrido - do que no genótipo tradicional EEA 406. A emissão de folhas é diferente em distintos biótipos de arroz-vermelho.
Oryza sativa; simulação; desempenho; estágio de Haun
The objective of this work was to simulate leaf appearance in cultivated rice genotypes and in red rice biotypes, by adapting the Wang and Engel model modified by Streck et al. Two experiments were conducted in Santa Maria, Rio Grande do Sul State, Brazil, during 2004/2005 and 2005/2006 growing seasons. Cultivars IRGA 417 and EEA 406, one hybrid, and two red rice biotypes were used. The leaf appearance model was adjusted, using Haun Stage (HS) data from five sowing dates of 2004/2005 growing season, and the model evaluation was performed using HS data from three sowing dates of 2005/2006 growing season. A numerical experiment was also carried out using temperature data occurred during 2003/2004 and 2006/2007 growing seasons, and simulated HS of the cultivated rice genotypes was compared with the red rice biotypes. The model had a good performance, with a root mean square error usually less than one leaf. Main stem leaf emergence was greater in modern genotypes IRGA 417 and hybrid than in the old genotype EEA 406. Leaf emergence is different among distinct red rice biotypes.
Oryza sativa; simulation; performance; Haun stage
AGROMETEOROLOGIA
Modelagem da emissão de folhas em arroz cultivado e em arroz-vermelho
Modeling leaf appearance in cultivated rice and red rice
Nereu Augusto Streck; Leosane Cristina Bosco; Dionéia Daiane Pitol Lucas; Isabel Lago
Universidade Federal de Santa Maria, Centro de Ciências Rurais, Departamento de Fitotecnia, CEP 97105-900 Santa Maria, RS. E-mail: nstreck2@yahoo.com.br, leosaneb@yahoo.com.br, dio.pitol@gmail.com, isalago08@yahoo.com.br
RESUMO
O objetivo deste trabalho foi simular o aparecimento de folhas em genótipos de arroz cultivados e em biótipos de arroz-vermelho por meio da adaptação do modelo de Wang e Engel, modificado por Streck et al. Dois experimentos foram conduzidos em Santa Maria, RS, em 2004/2005 e 2005/2006. Foram utilizadas as cultivares IRGA 417 e EEA 406, um híbrido e dois biótipos de arroz-vermelho. O modelo de aparecimento de folhas foi ajustado para os genótipos usando-se dados de estágio de Haun (HS), de cinco épocas de semeadura, em 2004/2005, e a avaliação do modelo foi feita com dados de HS medidos em três épocas de semeadura, em 2005/2006. Realizou-se também um experimento numérico utilizando-se dados de temperatura de 2003/2004 e de 2006/2007, e comparou-se o HS simulado dos genótipos cultivados em relação aos biótipos de arroz-vermelho. O modelo apresentou bom desempenho para simular o HS com raiz do quadrado médio do erro geralmente menor do que uma folha. A emissão de folhas na haste principal foi maior nos genótipos modernos - IRGA 417 e híbrido - do que no genótipo tradicional EEA 406. A emissão de folhas é diferente em distintos biótipos de arroz-vermelho.
Termos para indexação: Oryza sativa, simulação, desempenho, estágio de Haun.
ABSTRACT
The objective of this work was to simulate leaf appearance in cultivated rice genotypes and in red rice biotypes, by adapting the Wang and Engel model modified by Streck et al. Two experiments were conducted in Santa Maria, Rio Grande do Sul State, Brazil, during 2004/2005 and 2005/2006 growing seasons. Cultivars IRGA 417 and EEA 406, one hybrid, and two red rice biotypes were used. The leaf appearance model was adjusted, using Haun Stage (HS) data from five sowing dates of 2004/2005 growing season, and the model evaluation was performed using HS data from three sowing dates of 2005/2006 growing season. A numerical experiment was also carried out using temperature data occurred during 2003/2004 and 2006/2007 growing seasons, and simulated HS of the cultivated rice genotypes was compared with the red rice biotypes. The model had a good performance, with a root mean square error usually less than one leaf. Main stem leaf emergence was greater in modern genotypes IRGA 417 and hybrid than in the old genotype EEA 406. Leaf emergence is different among distinct red rice biotypes.
Index terms:Oryza sativa, simulation, performance, Haun stage.
Introdução
O arroz (Oryza sativa L.) é um dos três principais cereais produzidos no mundo, e é considerado o alimento básico para mais da metade da população mundial (FAO, 2004). O Brasil é o primeiro país não asiático com maior produção - 11,2 milhões de Mg por ano-, e maior consumo - 13 milhões de Mg por ano - de arroz no mundo (Conab, 2007). O arroz-vermelho (Oryza sativa L.) é a principal planta daninha de lavouras orizícolas e sua presença nas lavouras é um dos fatores que mais contribuem para que a produtividade de arroz irrigado seja menor que a produtividade potencial (Marchezan, 1994; Marchezan et al., 2004).
Caracterizar o crescimento e o desenvolvimento de diferentes genótipos de arroz (cultivares tradicionais, modernas e híbridos) e biótipos de arroz-vermelho pode ser importante para o estabelecimento de estratégias de controle do arroz-vermelho e para auxiliar no melhoramento do arroz cultivado, a fim de aumentar sua competitividade frente a esta planta daninha (Kwon et al., 1992; Noldin et al., 1999; Streck et al., 2006).
Uma maneira de caracterizar o crescimento e o desenvolvimento de plantas é pelo uso da modelagem. Um parâmetro de desenvolvimento de poáceas, bastante estudado pela sua importância durante a fase vegetativa, é a taxa de aparecimento de folhas (TAF) (Streck et al., 2003a, 2003b). O cálculo da TAF é uma importante parte de modelos de simulação de culturas (Hodges, 1991), incluindo modelos de arroz (Lee et al., 2001). A integração da TAF no tempo resulta no número de folhas (NF) acumuladas ou emergidas no colmo principal, que é considerada uma excelente medida de desenvolvimento. O NF em arroz está relacionado com o momento de ocorrência de estágios de desenvolvimento da planta como: afilhamento (Tivet et al., 2001; Jaffuel & Dauzat, 2005; Watanabe et al., 2005), iniciação da panícula (Ellis et al., 1993; Lee et al., 2001; Watanabe et al., 2005), emborrachamento e antese (Counce et al., 2000; Watanabe et al., 2005). A expansão da área foliar, que intercepta a radiação solar para fotossíntese no dossel das plantas e posterior acúmulo de matéria seca e produtividade da cultura, está também relacionada ao NF (Tivet et al., 2001). Assim, a TAF e o NF são parâmetros importantes em estudos de competição cultura-planta daninha. Nas poáceas, o NF é freqüentemente representado pelo estágio de Haun (HS), calculado pelo número de folhas completamente expandidas, mais a razão entre o comprimento da última (Ln) e o comprimento da penúltima folha (Ln-1) (Haun, 1973).
A temperatura é o fator do ambiente que mais influencia a taxa de aparecimento de folhas em arroz (Gao et al., 1992; Ellis et al., 1993; Sié et al., 1998). Um modelo bastante usado para estimar a TAF, em trigo, tem sido o modelo de Wang e Engel (WE) (Wang & Engel, 1998; Streck et al., 2003b, 2007). Esse modelo é simples e leva em conta a resposta não linear da temperatura sobre a TAF e combina os efeitos de fatores do ambiente sobre a TAF de forma multiplicativa, sendo por isso realístico do ponto vista biológico. Streck et al. (2003b) incorporaram uma função de resposta à cronologia no modelo WE que leva em consideração o efeito das reservas da semente na velocidade de aparecimento das primeiras duas folhas e a diminuição na TAF com o aumento do número de folhas. Esse modelo foi desenvolvido para trigo, uma espécie similar ao arroz em morfologia e hábito de crescimento.
A adaptação do modelo de WE, modificado por Streck et al. (2003b), para arroz e arroz-vermelho ainda não foi realizada. Uma vez adaptado para essa cultura e para essa planta daninha, esse modelo será uma importante ferramenta para estudar a habilidade competitiva de genótipos de arroz cultivado à presença de biótipos de arroz-vermelho.
O objetivo deste trabalho foi simular o aparecimento de folhas em genótipos de arroz cultivados e em biótipos de arroz-vermelho, por meio da adaptação do modelo de WE modificado por Streck et al. (2003b).
Material e Métodos
Dois experimentos em campo (nos anos agrícolas 2004/2005 e 2005/2006) foram realizados na área experimental do Departamento de Fitotecnia da Universidade Federal de Santa Maria (Santa Maria, RS, a 29º43'S, 53°43'W e 95 m de altitude). As plantas de arroz foram cultivadas em baldes (um genótipo por balde) de 12 L com solo Argissolo Vermelho distrófico arênico (Embrapa, 2006), enterrados, deixando-se uma borda de 5 cm acima do nível do solo. O espaçamento entre os baldes foi de 1,5x0,8 m.
As cultivares utilizadas foram IRGA 417, EEA 406, genótipo do Instituto Rio Grandense do Arroz (IRGA) - ainda não liberado comercialmente e, por isso, denominado neste trabalho como híbrido - e dois biótipos de arroz-vermelho, um com casca preta e aristado (AVCP) e o outro com casca amarela e aristado (AVCA). A cultivar IRGA 417 é da subespécie indica, tipo moderno, ciclo precoce (Sosbai, 2003), de porte baixo e uma das mais cultivadas atualmente no Rio Grande do Sul. O híbrido também é da subespécie indica e foi usado devido à heterose. A cultivar EEA 406 é da subespécie japonica, tipo tradicional, ciclo médio, de porte alto, cultivada nos anos 60 e 70 no Rio Grande do Sul e muito utilizada em experimentos como simuladora do arroz-vermelho (Agostinetto, 2003; Agostinetto et al., 2004).
As sementes dos dois biótipos de arroz-vermelho foram oriundas de uma única planta de cada biótipo, que haviam sido cultivados na área experimental do IRGA, em Cachoeirinha, RS, em 2003/2004, e representam dois biótipos freqüentemente encontrados nas lavouras arrozeiras do Rio Grande do Sul (Menezes et al., 2002). Anteriormente à semeadura foi realizada a quebra de dormência das sementes dos biótipos de arroz-vermelho (Marchezan, 1994), deixando-se as sementes em estufa durante cinco dias a 42°C.
Foram semeadas 30 sementes por balde, em cinco datas de semeadura em 2004/2005 (2/9/2004, de semeadura em 2005/2006 (26/9/2005, 25/11/2005, 2/2/2006). Estas datas foram usadas por se situarem antes, durante e depois da época recomendada de semeadura para a região que é de 1º/10 a 10/12 (Sosbai, 2003), para se obter o crescimento e o desenvolvimento das plantas em condições meteorológicas distintas. O delineamento experimental foi o inteiramente ao acaso, com quatro repetições, sendo que cada repetição (unidade experimental) foi composta por um balde com dez plantas.
O dia da emergência foi considerado quando 50% das plântulas estavam visíveis acima do solo. No estágio V3 da escala de Counce et al. (2000) foi realizado o desbaste, deixando-se dez plântulas por balde, cinco das quais foram marcadas com arames coloridos, o que resultou em população de aproximadamente 200 plantas m-2. O manejo das plantas foi baseado nas recomendações técnicas para a cultura do arroz irrigado (Sosbai, 2003) e a irrigação foi realizada deixando-se uma lâmina de água de 5 a 7 cm nos baldes a partir do estágio V3 da escala de Counce et al. (2000).
Nas plantas marcadas, foram medidos o número de folhas e o comprimento da última e penúltima folha do colmo principal, uma vez por semana. Com esse dados foi calculado o estágio de Haun - HS (Haun, 1973) pela da relação:

em que NF é o número de folhas do colmo principal, Ln é o comprimento da última folha e Ln - 1 é o comprimento da penúltima folha do colmo principal.
As temperaturas mínima e máxima diária do ar durante o experimento foram medidas na estação meteorológica convencional, pertencente ao 8º Distrito de Meteorologia, a aproximadamente 200 m da área experimental.
Utilizou-se o modelo da taxa de aparecimento de folhas de WE (Wang & Engel, 1998) modificado por Streck et al. (2003b) que, adaptado para arroz, tem a forma geral:

em que TAF é a taxa diária de aparecimento de folhas (folhas por dia), TAFmáx é a taxa diária máxima de aparecimento de folhas (folhas por dia), f(T) e f(C) são as funções de resposta à temperatura e à cronologia para TAF, respectivamente.
A função de temperatura f(T) é:
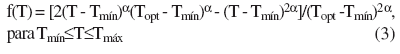

em que: Tmín, Topt, e Tmáx são as temperaturas cardinais (mínima, ótima e máxima, respectivamente) para TAF; e T é a temperatura média diária do ar (°C). As temperaturas cardinais mínima, ótima e máxima para TAF, em arroz, são 11°C (Ellis et al., 1993; Infeld et al., 1998), 26°C (Ellis et al., 1993) e 40°C (Gao et al., 1992), respectivamente.
A função de cronologia f(C) é:

em que: HS é o estágio de Haun e b é um coeficiente de sensibilidade com valor de -0,3 (Streck et al., 2003b). O HS no modelo de WE é calculado pelo acúmulo da TAF,ou seja, HS = STAF.
O coeficiente TAFmáx (equação 2) é dependente do genótipo. Esse coeficiente foi estimado para cada genótipo neste trabalho usando os dados de HS coletados nas cinco datas de semeadura em 2004/2005, pelo método dos mínimos quadrados (Streck et al., 2007). Os valores de TAFmáx estimados para cada genótipo e biótipo em cada época de semeadura foram submetidos à análise de variância e os valores médios das cinco datas de semeadura foram comparados entre genótipos pelo teste Tukey, a 5% de probabilidade.
Os valores de HS simulados pelo modelo foram comparados com os valores de HS observados para cada cultivar, durante as três datas de semeadura em 2005/2006, os quais são dados independentes. A estatística usada para avaliar o desempenho do modelo foi a raiz do quadrado médio do erro (RQME), calculada segundo a equação proposta por Janssen & Heuberger (1995):

em que: s são valores simulados de HS; o são valores observados de HS; e N é o número de observações. A unidade do RQME é a mesma da simulada ou observada, ou seja, folhas. Quanto menor o valor de RQME melhor a simulação.
Após a avaliação do desempenho do modelo, realizou-se experimento numérico no qual o HS foi simulado usando-se dados de temperatura coletados em campo em 2003/2004 e 2006/2007, com a finalidade de estudar a emissão de folhas dos biótipos de arroz-vermelho, em relação à emissão de folhas dos genótipos cultivados. Estes dois anos foram selecionados por apresentar condições meteorológicas distintas e são independentes dos dois anos de experimento, o que é importante em estudos numéricos. Para tanto, foram consideradas três datas de emergência do arroz cultivado dentro da época recomendada (1º/10, 5/11 e 10/12) e cinco datas de emergência do arroz-vermelho em relação à emergência do arroz cultivado (+4, +2, 0, -2, -4 dias), representando diferentes fluxos de emergência de arroz-vermelho em uma lavoura. Esse experimento numérico foi analisado separadamente comparando-se o HS dos biótipos de arroz-vermelho com o HS dos genótipos cultivados.
Resultados e Discussão
As condições meteorológicas foram distintas durante os anos do experimento de campo (2004/2005 e 2005/2006). A temperatura mínima do ar variou de 6,6 a 25,4°C em 2004/2005 e de 6,9 a 25°C em 2005/2006, enquanto a temperatura máxima do ar variou de 17,2 a 38,4°C em 2004/2005, e de 16,6 a 38,6°C em 2005/2006. A exposição das plantas de arroz a distintas condições meteorológicas é importante para calibrar e avaliar o modelo de TAF. Durante os dois anos usados no experimento numérico (2003/2004 e 2006/2007), também houve variação na temperatura do ar, tendo a temperatura mínima do ar de 27/9 (primeiro dia de simulação) a 12/2 (último dia de simulação) variado de 8 a 23,4°C em 2003/2004, e de 10 a 25,8°C em 2006/2007, enquanto a temperatura máxima do ar variou de 18,2 a 38,4°C em 2003/2004, e de 18,4 a 37°C em 2006/2007.
A TAFmáx da cultivar IRGA 417 só diferiu significativamente da TAFmáx da cultivar EEA 406 (Tabela 1), e essa diferença indica que o melhoramento genético alterou o desenvolvimento, aumentando significativamente a velocidade de emissão de folhas na cultivar moderna. Por outro lado, o fato de a TAFmáx não ser diferente estatisticamente, entre os dois biótipos de arroz-vermelho (AVCA e AVCP) e a cultivar EEA 406, indica a similaridade também no desenvolvimento entre o arroz-vermelho e esta cultivar, que tem sido usada como planta simuladora do arroz-vermelho em estudos de competição desta planta daninha com o arroz cultivado (Agostinetto, 2003; Agostinetto et al., 2004).
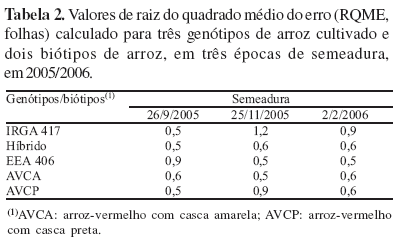
No modelo (equação 2), optou-se em usar os valores de TAFmáx estimados para cada genótipo (Tabela 1), apesar de não ter havido diferença significativa entre vários genótipos. O desempenho do modelo em simular o HS na haste principal variou entre os genótipos e biótipos com RQME geral - considerando todas as datas de semeadura - de 0,9 folha para a cultivar IRGA 417; 0,6 folha para o Híbrido; 0,7 folha para a cultivar EEA 406; 0,6 folha para o biótipo AVCA e 0,7 folha para o biótipo AVCP (Tabela 2). Estes valores de RQME são similares aos valores de RQME em trigo (Streck et al., 2003b, 2007).
Não se observou diminuição ou aumento do RQME em relação à data de semeadura (Tabela 2). Os valores de RQME indicaram que, na maioria das épocas e genótipos, o erro da estimativa de HS foi inferior a uma folha (Tabela 2), erro considerado pequeno, caracterizando bom desempenho do modelo (Streck et al., 2007). Um erro de até uma folha, em geral, não modifica a época de aplicação de medidas de controle de plantas daninhas na cultura do arroz, como por exemplo, a aplicação de herbicida pós-emergente (Sosbai, 2003). Esses resultados indicam que o modelo da TAF, inicialmente proposto e testado para trigo (Streck et al., 2003b), funciona bem para arroz.
O bom desempenho do modelo da TAF justificou sua utilização no experimento numérico visando a estudar a emissão de folhas dos genótipos cultivados em relação à emissão de folhas dos biótipos de arroz-vermelho, em diferentes épocas de semeadura, em 2003/2004 e 2006/2007. Os resultados deste estudo numérico foram similares entre os anos agrícolas, mesmo esses anos sendo distintos em relação às condições meteorológicas (Figuras 1 a 4).
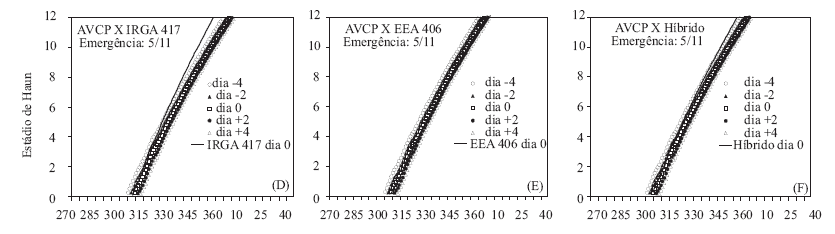
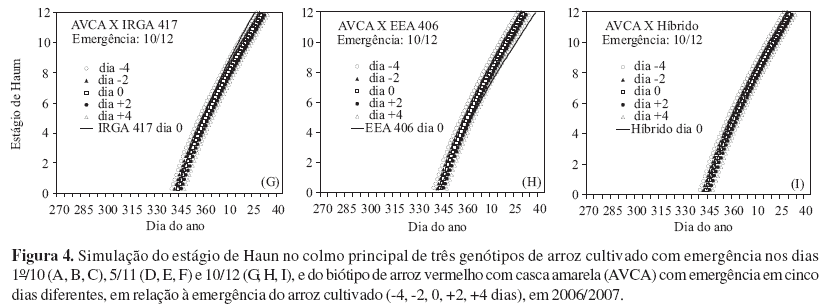
As Figuras 1 e 3 apresentam a comparação do HS simulado para os genótipos cultivados e para distintos dias de emergência do arroz-vermelho AVCP. Devido à menor TAF do AVCP, nos dois anos e nas três épocas de semeadura, o HS deste foi menor que o HS do IRGA 417 e do híbrido, a partir de 5-6 folhas, mesmo quando a emergência do AVCP ocorreu quatro dias antes desses genótipos (Figuras 1 A, D e G e 3 A, D e G). Já o genótipo EEA 406, devido à menor TAF, emitiu folhas mais lentamente que o AVCP e, mesmo quando o biótipo de arroz-vermelho emergiu dois dias após o EEA 406, o HS no final da fase vegetativa foi maior no AVCP (Figuras 1 B, E e H e 3 B, E e H). Com relação ao híbrido, o HS é maior que o AVCP emergido quatro dias antes, quando o número de folhas foi maior que oito (Figuras 1 C, F e I e 3 C, F e I), indicando que o vigor ou heterose também se manifestou na velocidade de emissão de folhas.
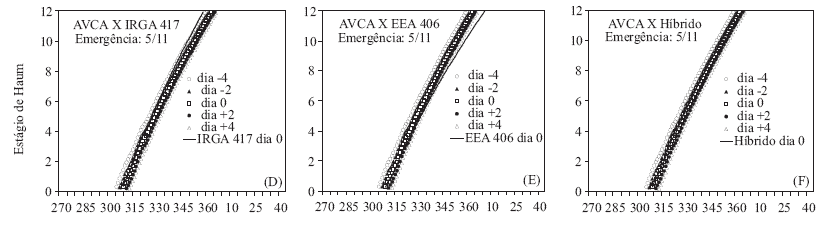
Quanto à comparação do HS simulado, para os genótipos cultivados e para distintos dias de emergência do arroz-vermelho AVCA, nota-se que também o HS dos dois genótipos cultivados (IRGA 417 e híbrido) foram maiores no final da fase vegetativa, mas a diferença não foi grande quanto simulado para o outro biótipo de arroz-vermelho, pois a TAFmáx do AVCA é maior que do AVCP, inclusive com o híbrido não tendo maior HS do que o AVCA, quando este emergiu quatro dias antes (Figuras 2 C, F e I e 4 C, F e I). Com relação ao EEA 406, comparado com o AVCA, após aproximadamente oito folhas, o HS do AVCA foi sempre superior (Figuras 2 B, E e H e 4 B, E e H), possivelmente, indicando que o biótipo AVCA é mais competitivo com o arroz cultivado do que o biótipo de arroz-vermelho AVCP.
A maior velocidade de aparecimento de folhas do biótipo de arroz-vermelho AVCA, comparado com o biótipo de arroz-vermelho AVCP, talvez esteja relacionada com diferentes graus de cruzamento dos dois biótipos com o arroz cultivado. Através do fluxo gênico, os biótipos de arroz-vermelho podem sofrer introgressão em maior ou menor intensidade de características de genótipos de arroz cultivado, conferindo características diferentes entre eles (Gealy et al., 2003). Ainda, por estar sujeitos ao processo de seleção natural, fixam as características relacionadas à maior velocidade de crescimento inicial, maior área foliar e maior afilhamento (Diarra et al., 1985; Gealy et al., 2003). Isso reflete a condição de planta daninha em constante processo de evolução e associação com a cultura que infesta, e é por isso que deve haver um continuo processo de melhoramento genético em relação aos genótipos atuais.
Conclusões
1. O modelo de taxa de aparecimento de folhas de WE modificado por Streck et al. tem bom desempenho na simulação de estágio de Haun, em arroz cultivado e em arroz-vermelho, com RQME menor do que uma folha.
2. A emissão de folhas na haste principal é maior em genótipos de arroz moderno do que em genótipo de arroz cultivado tradicional.
3. A emissão de folhas é diferente em distintos biótipos de arroz-vermelho.
Agradecimentos
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico e à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, pela concessão de bolsas.
Recebido em 26 de outubro de 2007 e aprovado em 14 de maio de 2008
- AGOSTINETTO, D. Aplicação de modelos matemáticos para estimar interferência de genótipos de arroz concorrentes e níveis de dano econômico em arroz irrigado 2003. 200p. Tese (Doutorado) - Universidade Federal do Rio Grande do Sul, Porto Alegre.
- AGOSTINETTO, D.; FLECK, N.G.; RIZZARDI, M.A.; BIANCHI, M.A.; SILVA, P.R.F. da. Comparação de modelos matemáticos na estimativa das perdas de produtividade de grãos em arroz irrigado. Pesquisa Agropecuária Brasileira, v.39, p.847-856, 2004.
- CONAB. Evolução da produção de grãos no Brasil Disponível em: http://www.conab.gov.br Acesso em: 5 fev. 2007.
- COUNCE, P.; KEISLING, T.C.; MITCHELL, A.J. A uniform, objective, and adaptive system for expressing rice development. Crop Science, v.40, p.436-443, 2000.
- DIARRA, A.; SMITH JÚNIOR, R.J.; TALBERT, R.E. Growth and morphological characteristics of red rice (Oryza sativa) biotypes. Weed Science, v.33, p.310-314, 1985.
- ELLIS, R.H.; QI, A.; SUMMERFIELD, R.J.; ROBERTS, E.H. Rates of leaf appearance and panicle development in rice (Oryza sativa L.): a comparison at three temperatures. Agricultural and Forest Meteorology, v.66, p.129-138, 1993.
- EMBRAPA. Sistema brasileiro de classificação de solos 2.ed. Brasília: Embrapa Informação Tecnológica; Rio de Janeiro: Embrapa Solos, 2006. 306p.
- FAO. International year of rice 2004. Disponível em: http://www.fao.org/rice2004/en/speeches.htm Acesso em: 5 fev. 2007.
- GAO, L.; JIN, Z.; HUANG, Y.; ZHANG, L. Rice clock model: a computer model to simulate rice development. Agricultural and Forest Meteorology, v.60, p.1-16, 1992.
- GEALY, D.R.; MITTEN, D.H.; RUTGER, J.N. Gene flow between red rice (Oryza sativa) and herbicide-resistant rice (O. sativa): implications for weed management. Weed technology, v.17, p.627-645, 2003.
- HAUN, J.R. Visual quantification of wheat development. Agronomy Journal, v.65, p.116-119, 1973.
- HODGES, T. Crop growth simulation and the role of phenological models. In: HODGES, T. (Ed.). Predicting crop phenology Boston: CRC Press, 1991. p.3-5.
- INFELD, J.A.; SILVA, J.B.; ASSIS, F.N. Temperatura-base e graus-dia durante o período vegetativo de três grupos de cultivares de arroz irrigado. Revista Brasileira de Agrometeorologia, v.6, p.187-191, 1998.
- JAFFUEL, S.; DAUZAT, J. Synchronism of leaf and tiller emergence relative to position and to main stem development stage in a rice cultivar. Annals of Botany, v.95, p.401-412, 2005.
- JANSSEN, P.H.M.; HEUBERGER, P.S.C. Calibration of process-oriented models. Ecological Modelling, v.83, p.55-56, 1995.
- KWON, S.L.; SMITH JÚNIOR, R.J.; TALBERT, R.E. Comparative growth and development of red rice (Oryza sativa) and rice (Oryza sativa). Weed Science, v.40, p.57-62, 1992.
- LEE, C.K.; LEE, B.W.; SHIN, J.C.; YOON, Y.H. Heading date and final leaf number as affected by sowing date and prediction of heading date based on leaf appearance model in rice. Korean Journal of Crop Science, v.46, p.195-201, 2001.
- MARCHEZAN, E. Arroz vermelho: caracterização, prejuízos e controle. Ciência Rural, v.24, p.415-421, 1994.
- MARCHEZAN, E.; ÁVILA, L.A.; MAGALHÃES JÚNIOR, A.M.; MACHADO, S.L.; PETRINI, J.A. Controle do arroz vermelho. In: GOMES, A.S.; MAGALHÃES JÚNIOR, A.M. Arroz irrigado no Sul do Brasil Brasília: Embrapa Informação Tecnológica, 2004. p.547-573.
- MENEZES, V.G.; AGOSTINETTO, D.; FLECK, N.G.; SILVEIRA, C.A. Caracterização de biótipos de arroz-vermelho em lavouras de arroz no Estado do Rio Grande do Sul. Planta Daninha, v.20, p.221-227, 2002.
- NOLDIN, J.A.; CHANDLER, J.M.; MCCAULEY, G.N. Red rice (Oryza sativa) biology. I. Characterization of red rice ecotypes. Weed Technology, v.13, p.12-18, 1999.
- SIÉ, M.; DINGKUHN, M.; WOPEREIS, M.C.S.; MIÉZAN, K.M. Rice crop duration and leaf appearance rate in a variable thermal environment. I. Development of an empirically based model. Field Crops Research, v.57, p.1-13, 1998.
- SOCIEDADE SUL BRASILEIRA DE ARROZ IRRIGADO. Arroz irrigado: recomendações técnicas para o sul do Brasil. Itajaí: Sosbai, 2003. 126p.
- STRECK, N.A.; BOSCO, L.C.; LAGO, I. Simulating leaf appearance of some Brazilian wheat genotypes. Revista Brasileira de Agrometeorologia, v.15, p.1-9, 2007.
- STRECK, N.A.; BOSCO, L.C.; MICHELON, S.; WALTER, L.C.; MARCOLIN, E. Duração do ciclo de desenvolvimento de cultivares de arroz em função da emissão de folhas no colmo principal. Ciência Rural, v.36, p.1086-1093, 2006.
- STRECK, N.A.; WEISS, A.; XUE, Q.; BAENZIGER, P.S. Improving predictions of developmental stages in winter wheat: a modified Wang and Engel model. Agricultural and Forest Meteorology, v.115, p.139-150, 2003a.
- STRECK, N.A.; WEISS, A.; XUE, Q.; BAENZIGER, P.S. Incorporating a chronology response into the prediction of leaf appearance rate in winter wheat. Annals of Botany, v.92, p.181-190, 2003b.
- TIVET, F.; PINHEIRO, B.S.; RAISSAC, M.; DINGKUHN, M. Leaf blade dimensions of rice (Oryza sativa L. and Oryza glaberrima Stend.). Relationships between tillers and the main stem. Annals of Botany, v.88, p.507-511, 2001.
- WANG, E.; ENGEL, T. Simulation of phenological development of wheat crops. Agricultural Systems, v.58, p.1-24, 1998.
- WATANABE, T.; HANAN, P.M.R.; HAZEGAWA, T.; NAKAGAWA, H.; TAKAHASHI, W. Rice morphogenesis and plant architecture: measurement, specification and the reconstruction of structural development by 3D architectural modeling. Annals of Botany, v.95, p.1131-1143, 2005.
Datas de Publicação
-
Publicação nesta coleção
19 Jun 2008 -
Data do Fascículo
Maio 2008
Histórico
-
Recebido
26 Out 2007 -
Aceito
14 Maio 2008