
Abstract:
The objective of this work was to evaluate, by genomic in situ hybridization (GISH), pairing configurations as potential indicators of recombination between chromosomes of different parental genomes, in two interspecific hybrids (elephant grass x pearl millet) artificially polyploidized. Anthers from young flower buds were used in the chromosomal preparations. The genomic probe was prepared with pearl millet DNA and labeled with biotin-16-dUTP by the nick translation reaction. Blocking DNA was prepared with genomic elephant grass DNA. The homoeologous intergenomic pairing, observed in the two hybrids, indicates the possibility of recombination between chromosomes of the parental genomes.
Index terms:
Pennisetum glaucum; Pennisetum purpureum; in situ hybridization; intergenomic pairing; interspecific hybridization
Resumo:
O objetivo deste trabalho foi avaliar, por meio da hibridização in situ genômica (GISH), as configurações de pareamento como indicadoras potenciais da recombinação entre cromossomos de diferentes genomas parentais, presentes em dois híbridos interespecíficos (capim-elefante x milheto) poliploidizados artificialmente. Anteras de botões florais jovens foram utilizadas nas preparações cromossômicas. A sonda genômica foi preparada com o DNA do milheto e marcada com biotina-16-dUTP pela reação de nick translation. O DNA de bloqueio foi preparado com o DNA genômico do capim-elefante. O pareamento intergenômico homeólogo, observado nos dois híbridos, indica a possibilidade de recombinação entre os cromossomos dos genomas parentais.
Termos para indexação:
Pennisetum glaucum; Pennisetum purpureum; hibridização in situ; pareamento intergenômico; hibridação interespecífica
Elephant grass (Pennisetum purpureum Schum., 2n=4x=28, A’A’BB) and pearl millet [Pennisetum glaucum (L.) R. BR., 2n=2x=14, AA genome] are important forage grasses (Pereira et al., 2001PEREIRA, A.V.; VALLE, C.B.; FERREIRA, R.P.; MILES, J.W. Melhoramento de forrageiras tropicais. In: NASS, L.L.; VALOIS, A.C.C.; MELO, I.S.; VALADARES-INGLIS, M.C. (Ed.). Recursos genéticos e melhoramento - plantas. Rondonópolis: Fundação Mato Grosso, 2001. p.549-602.). The cross between them produces a triploid (2n=3x=21, AA’B) and sterile hybrid (Techio et al., 2006TECHIO, V.H.; DAVIDE, L.C.; PEREIRA, A.V. Meiosis in elephant grass (Pennisetum purpureum), pearl millet (Pennisetum glaucum) (Poaceae, Poales) and their interspecific hybrids. Genetics and Molecular Biology, v.29, p.353-362, 2006. DOI: 10.1590/S1415-47572006000200025.
https://doi.org/10.1590/S1415-4757200600...
), with greater forage quality and better acceptance by cattle than the parental species of these plants (Pereira et al., 2001PEREIRA, A.V.; VALLE, C.B.; FERREIRA, R.P.; MILES, J.W. Melhoramento de forrageiras tropicais. In: NASS, L.L.; VALOIS, A.C.C.; MELO, I.S.; VALADARES-INGLIS, M.C. (Ed.). Recursos genéticos e melhoramento - plantas. Rondonópolis: Fundação Mato Grosso, 2001. p.549-602.). Production of artificial polyploid hybrids (2n=6x=42, AAA’A’BB) partially restores fertility and allow the incorporation of interspecific hybrid in breeding programs (Paiva et al., 2012PAIVA, E.A.A.; BUSTAMANTE, F.O.; BARBOSA, S.; PEREIRA, A.V.; DAVIDE, L.C. Meiotic behavior in early and recent duplicated hexaploid hybrids of napier grass (Pennisetum purpureum) and pearl millet (Pennisetum glaucum). Caryologia: International Journal of Cytology, Cytosystematics and Cytogenetics, v.65, p.114-120, 2012. DOI: 10.1080/00087114.2012.709805.
https://doi.org/10.1080/00087114.2012.70...
). However, these artificial hybrids are partial hexaploids, since they present mixoploidy (14 to 42 chromosomes), and mitotic (Abreu et al., 2006ABREU, J.C. de; DAVIDE, L.C.; PEREIRA, A.V.; BARBOSA, S. Mixoploidia em híbridos de capim-elefante x milheto tratados com agentes antimitóticos. Pesquisa Agropecuária Brasileira, v.41, p.1629-1635, 2006. DOI: 10.1590/S0100-204X2006001100009.
https://doi.org/10.1590/S0100-204X200600...
) and meiotic irregularities (Paiva et al., 2012PAIVA, E.A.A.; BUSTAMANTE, F.O.; BARBOSA, S.; PEREIRA, A.V.; DAVIDE, L.C. Meiotic behavior in early and recent duplicated hexaploid hybrids of napier grass (Pennisetum purpureum) and pearl millet (Pennisetum glaucum). Caryologia: International Journal of Cytology, Cytosystematics and Cytogenetics, v.65, p.114-120, 2012. DOI: 10.1080/00087114.2012.709805.
https://doi.org/10.1080/00087114.2012.70...
).
The genomic in situ hybridization (GISH) of pearl millet genomic DNA (AA) on pearl millet, elephant grass (A’A’BB), and on triploid hybrid (AA’B) chromosomes allows for distinguishing between the three genomes that make up the hybrid. GISH has confirmed that the A and A’ genomes are related to each other and, to a lower degree, to the B genome (Reis et al., 2014REIS, G.B. dos; MESQUITA, A.T.; TORRES, G.A.; ANDRADE-VIEIRA, L.F.; PEREIRA, A.V.; DAVIDE, L.C. Genomic homeology between Pennisetum purpureum and Pennisetum glaucum (Poaceae). Comparative Cytogenetics, v.8, p.199-209, 2014. DOI: 10.3897/CompCytogen.v8i3.7732.
https://doi.org/10.3897/CompCytogen.v8i3...
). The chromosomal homoeology between these genomes enables intergenomic recombination and, therefore, the generation of new gene combinations with potential use in breeding programs. However, the types and frequency of interaction between these genomes, in the meiosis of these interspecific polyploid hybrids, are still poorly understood.
The objective of this work was to evaluate, by genomic in situ hybridization (GISH), pairing configurations as potential indicators of recombination between chromosomes of different parental genomes, in two interspecific hybrids (elephant grass x pearl millet) artificially polyploidized.
Two hexaploid hybrids, obtained after chromosome doubling of interspecific triploid by treatment with mitosis inhibitor (Abreu et al., 2006ABREU, J.C. de; DAVIDE, L.C.; PEREIRA, A.V.; BARBOSA, S. Mixoploidia em híbridos de capim-elefante x milheto tratados com agentes antimitóticos. Pesquisa Agropecuária Brasileira, v.41, p.1629-1635, 2006. DOI: 10.1590/S0100-204X2006001100009.
https://doi.org/10.1590/S0100-204X200600...
), were used: the commercial cultivar Paraíso and PCM0702-30, from the breeding program of Embrapa Gado de Leite, Juiz de Fora, MG, Brazil. Young flower buds were collected from five plants of each genotype. Chromosome preparations were made with anthers from 1-2 buds per slide. The genomic probe was prepared with pearl millet DNA, and labeled with biotin-16-dUTP using nick translation reaction. The blocking DNA was prepared with elephant grass genomic DNA. Both DNAs were extracted by the 2% CTAB method (Doyle & Doyle, 1987DOYLE, J.J.; DOYLE, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin, v.19, p.11-15, 1987.). The identification of intragenomic (A/A, A’/A’ or B/B), intergenomic (A/A’, A/B, and A’/B), and interparental (A/A’B) pairing configuration was made in 15 diakinesis of the PCM0702-30 genotype and in 25 diakinesis of the Paraíso cultivar.
In both hybrids, intra- and intergenomic pairings were observed in univalent, bivalent (ring and rod), and multivalent configurations (Figure 1). For various pairing configuration, there was variation between the two hybrids and also in the same hybrid when compared the involvement of either pearl millet or Napier grass chromosomes (Table 1). These variations may be related to the homoeology between the three genomes (Figure 1), and to the mixoploid condition of the hybrids.
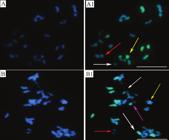
In situ hybridization of polyploid hybrids between elephant grass and pearl millet, using pearl millet genomic probe in diakinesis. Chromosomes are visualized with DAPI (A and B). Pairing configurations: signal of the probe (in green) shows millet chromosomes belonging to the univalents from genome A. In A1, white arrow points to the univalent belonging to genome B; red arrow, to the bivalent with intragenomic pairing; and yellow arrow, to the multivalent in intergenomic pairing, involving the genomes A, A’, and B. In B1, red arrow points to the open bivalent (rod) with intragenomic pairing; yellow arrow, to the bivalent intragenomic pairing between chromosomes of the genome A’; pink arrow, to the open bivalent (rod) with intergenomic pairing between genomes A and A’; and white arrows, to the genome multivalents. Bar: 10 μm.
Percentage of cells in diakinesis, showing each of the different configurations in the artificial polyploid hybrids between Pennisetum purpureum 'Paraíso' and P. glaucum 'PCM 0702-30'.
The number of counted chromosomes varied from 22 to 38 for the Paraíso cultivar and from 26 to 38 for the PCM0702-30 hybrid. The high frequency of univalents of both parentals is a consequence of mixoploidy, since it shows the absence of the homologues in the evaluated meiocytes. Univalents of both parents were observed in all cells of the Paraíso cultivar, whereas in the PCM0702-30 genotype the frequency of cells from elephant grass and pearl millet with univalent was 71.42 and 39.28%, respectively (Table 1).
The occurrence of ring bivalents is an indication of high affinity between chromosomes, as proposed by Singh (1993)SINGH, R.J. Plant cytogenetics. Boca Raton: CRC Press, 1993. 391p., and it suggests that these bivalents involve chromosomes belonging to the same genome (Table 1). The frequency of rod bivalents and multivalents formed by elephant grass chromosomes was high, in comparison to the pearl millet’s ones, indicating the possibility of recombination between the A’ and B genomes, despite the lower degree of homology between them when compared to genome A.
GISH also showed a low frequency of cells with intergenomic bivalents, formed by the chromosome pairing of genomes A and A’, or A’ and B. Nonetheless, about 90% of cells of both genotypes showed interparental multivalents involving chromosomes of genomes A, A’, and B, in different combinations (Table 1). Intergenomic bivalents and multivalents indicate high degree of homology between chromosomes of different species, and point to the possibility of allele transfer by recombination between their chromosomes in the interspecific hybrid (Techio & Davide, 2007TECHIO, V.H.; DAVIDE, L.C. Análise genômica em cromossomos de plantas com base no pareamento meiótico. Biotemas, v.20, p.7-18, 2007.).
Genomes A and A’ are more closely related because of their origin from a common ancestor (Reis et al., 2014REIS, G.B. dos; MESQUITA, A.T.; TORRES, G.A.; ANDRADE-VIEIRA, L.F.; PEREIRA, A.V.; DAVIDE, L.C. Genomic homeology between Pennisetum purpureum and Pennisetum glaucum (Poaceae). Comparative Cytogenetics, v.8, p.199-209, 2014. DOI: 10.3897/CompCytogen.v8i3.7732.
https://doi.org/10.3897/CompCytogen.v8i3...
). In the triploid hybrids, seven bivalents are formed between the A and A’ genomes, and seven univalents from the B genome (Techio et al., 2006TECHIO, V.H.; DAVIDE, L.C.; PEREIRA, A.V. Meiosis in elephant grass (Pennisetum purpureum), pearl millet (Pennisetum glaucum) (Poaceae, Poales) and their interspecific hybrids. Genetics and Molecular Biology, v.29, p.353-362, 2006. DOI: 10.1590/S1415-47572006000200025.
https://doi.org/10.1590/S1415-4757200600...
). Paiva et al. (2012)PAIVA, E.A.A.; BUSTAMANTE, F.O.; BARBOSA, S.; PEREIRA, A.V.; DAVIDE, L.C. Meiotic behavior in early and recent duplicated hexaploid hybrids of napier grass (Pennisetum purpureum) and pearl millet (Pennisetum glaucum). Caryologia: International Journal of Cytology, Cytosystematics and Cytogenetics, v.65, p.114-120, 2012. DOI: 10.1080/00087114.2012.709805.
https://doi.org/10.1080/00087114.2012.70...
had already shown the formation of univalents and multivalents in polyploid hybrids of elephant grass and pearl millet; however, they had not identified the parental genomes. In our work, the presence of intergenomic multivalents and bivalents confirms the occurrence of chromosome homoeology, verified by those authors, as well as the possibility of gene transfer (Figure 1).
The observation of multivalents involving chromosomes of both parents is an indication of the potential for obtaining new gene combinations in duplicate hybrids. These combinations can be used in elephant grass breeding programs, by transferring genes that control characteristics of interest, as it was already done in wheat breeding, with the use of partial hybrids among Triticum and Aegilops (Tiwari, 2010TIWARI, V.K.; RAWAT, N.; NEELAM, K.; KUMAR, S.; RANDHAWA, G.S.; DHALIWAL, H.S. Random chromosome elimination in synthetic Triticum-Aegilops amphiploids leads to development of a stable partial amphiploid with high grain micro- and macronutrient content and powdery mildew resistance. Genome, v.53, p.1053-1065, 2010. DOI: 10.1139/G10-083.
https://doi.org/10.1139/G10-083....
). The difference in the pairing behavior of the two hybrids suggests the need to evaluate different hybrids to maximize the chance of obtaining combinations with greater potential for use in breeding programs.
Acknowledgements
To Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and to the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (Fapemig), for financial support; to Dr. Juliane Dornelas Nunes, for her assistance in collecting the material.
References
- ABREU, J.C. de; DAVIDE, L.C.; PEREIRA, A.V.; BARBOSA, S. Mixoploidia em híbridos de capim-elefante x milheto tratados com agentes antimitóticos. Pesquisa Agropecuária Brasileira, v.41, p.1629-1635, 2006. DOI: 10.1590/S0100-204X2006001100009.
» https://doi.org/10.1590/S0100-204X2006001100009. - DOYLE, J.J.; DOYLE, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin, v.19, p.11-15, 1987.
- PAIVA, E.A.A.; BUSTAMANTE, F.O.; BARBOSA, S.; PEREIRA, A.V.; DAVIDE, L.C. Meiotic behavior in early and recent duplicated hexaploid hybrids of napier grass (Pennisetum purpureum) and pearl millet (Pennisetum glaucum). Caryologia: International Journal of Cytology, Cytosystematics and Cytogenetics, v.65, p.114-120, 2012. DOI: 10.1080/00087114.2012.709805.
» https://doi.org/10.1080/00087114.2012.709805. - PEREIRA, A.V.; VALLE, C.B.; FERREIRA, R.P.; MILES, J.W. Melhoramento de forrageiras tropicais. In: NASS, L.L.; VALOIS, A.C.C.; MELO, I.S.; VALADARES-INGLIS, M.C. (Ed.). Recursos genéticos e melhoramento - plantas. Rondonópolis: Fundação Mato Grosso, 2001. p.549-602.
- REIS, G.B. dos; MESQUITA, A.T.; TORRES, G.A.; ANDRADE-VIEIRA, L.F.; PEREIRA, A.V.; DAVIDE, L.C. Genomic homeology between Pennisetum purpureum and Pennisetum glaucum (Poaceae). Comparative Cytogenetics, v.8, p.199-209, 2014. DOI: 10.3897/CompCytogen.v8i3.7732.
» https://doi.org/10.3897/CompCytogen.v8i3.7732. - SINGH, R.J. Plant cytogenetics. Boca Raton: CRC Press, 1993. 391p.
- TECHIO, V.H.; DAVIDE, L.C. Análise genômica em cromossomos de plantas com base no pareamento meiótico. Biotemas, v.20, p.7-18, 2007.
- TECHIO, V.H.; DAVIDE, L.C.; PEREIRA, A.V. Meiosis in elephant grass (Pennisetum purpureum), pearl millet (Pennisetum glaucum) (Poaceae, Poales) and their interspecific hybrids. Genetics and Molecular Biology, v.29, p.353-362, 2006. DOI: 10.1590/S1415-47572006000200025.
» https://doi.org/10.1590/S1415-47572006000200025. - TIWARI, V.K.; RAWAT, N.; NEELAM, K.; KUMAR, S.; RANDHAWA, G.S.; DHALIWAL, H.S. Random chromosome elimination in synthetic Triticum-Aegilops amphiploids leads to development of a stable partial amphiploid with high grain micro- and macronutrient content and powdery mildew resistance. Genome, v.53, p.1053-1065, 2010. DOI: 10.1139/G10-083.
» https://doi.org/10.1139/G10-083.
Publication Dates
-
Publication in this collection
Sept 2017
History
-
Received
29 Nov 2016 -
Accepted
02 Mar 2017

