Resumos
Objetivou-se avaliar alterações físicas e físico-químicas, relacionando-as às taxas respiratória e de liberação de etileno, durante a maturação da graviola-'Morada'. Frutos colhidos na maturidade fisiológica foram armazenados (23,4 ± 1,1ºC e 81,8 ± 10,6% UR) e avaliados aos 0; 1; 2; 3; 4 e 6 dias, quanto a: atividade respiratória (RS), liberação de etileno (ET), perda de matéria fresca, cor da casca e da polpa, firmeza, pH, acidez total titulável (ATT), sólidos solúveis totais (SST) e açúcares solúveis totais (AST). Utilizou-se o delineamento experimental inteiramente casualizado, com quatro repetições. Aos dois dias, iniciou-se rápido aumento na RS, resultando no primeiro pico (197,60mg CO2·kg-1·h-1), que foi seguido por queda e uma fase lag. Ao término desta, iniciou-se o aumento climatérico (pico = 298,82mg CO2·kg-1·h-1). ET só foi detectada por ocasião do primeiro pico respiratório, atingindo o máximo aos quatro dias. A cor da casca tornou-se mais clara, enquanto na polpa se observou apenas redução da luminosidade. Houve coincidência entre o primeiro aumento na RS, o pico de ET e as mudanças mais significativas na firmeza, na ATT e nos teores de SST e AST. Durante o período, a firmeza diminuiu de 60 para 0,9N, e a ATT aumentou de 0,18 para 0,88% de ácido cítrico.
annonaceae; etileno; maturação; respiração
It was aimed to evaluate physical and physico-chemical changes during soursop 'Morada' maturation at room temperature and related them to respiration rate and ethylene production. Fruits harvested at physiological maturity were stored (23,4 ± 1,1ºC and 81,8 ± 10,6% RH) and evaluated at 0, 1, 2, 3, 4, and 6 days, according to: respiration rate (RS), ethylene production (ET), fresh weight lost, skin and pulp color, firmness, pH, total titrable acidity (TTA), total soluble solids (TSS) and total soluble sugars (TS). The experimental design was a completely randomized one with four replications. There was a fast increase in RS at two days that resulted in the first peak (197,60mg CO2·kg-1·h-1) that was followed by a decay and a lag phase. When that phase rad finished, it began the climateric raise (climateric peak = 298,82mg CO2·kg-1·h-1). ET was detected only when the first respiratory peak occurred and reached the maximum at the fourth day. The skin color became bright green while it was observed a decrease just on lightness in the pulp. There was a coincidence among the first increase on RS, the peak of ET and the most significant changes on firmness, TAA, TSS content and TS content. During this period, firmness reduced from 60 to 0,9N and TTA increased from 0,18 to 0,88% of citric acid.
annonaceae; ethylene; maturation; respiration
Comportamento respiratório e qualidade pós-colheita de graviola (Annona muricata L.) 'morada' sob temperatura ambiente1 1 (Trabalho 085/2002). Parte da tese de doutorado do primeiro autor apresentada à Universidade Federal do Ceará, financiada com recursos do Programa Avança Brasil.
Respiratory behavior and postharvest quality of 'Morada' soursop (Annona muricata L.) at room temperature
Maria Auxiliadora Coêlho de LimaI; Ricardo Elesbão AlvesII; Heloísa Almeida Cunha FilgueirasII; Joaquim Enéas-FilhoIII
IDr., Pesquisadora Embrapa Semi-Árido, CP 23, CEP 56300-970, Petrolina-PE. maclima@cpatsa.embrapa.br
IIDr., Bolsista PQ/CNPq, Pesquisador Embrapa Agroindústria Tropical, CP 3761, 60511-110, Fortaleza-CE. elesbao@cnpat.embrapa.br, heloisa@cnpat.embrapa.br
IIIDr., Professor, Depto. de Bioquímica e Biologia Molecular, UFC, CP 1065, 60541-970, Fortaleza-CE. joaquime@ufc.br
RESUMO
Objetivou-se avaliar alterações físicas e físico-químicas, relacionando-as às taxas respiratória e de liberação de etileno, durante a maturação da graviola-'Morada'. Frutos colhidos na maturidade fisiológica foram armazenados (23,4 ± 1,1ºC e 81,8 ± 10,6% UR) e avaliados aos 0; 1; 2; 3; 4 e 6 dias, quanto a: atividade respiratória (RS), liberação de etileno (ET), perda de matéria fresca, cor da casca e da polpa, firmeza, pH, acidez total titulável (ATT), sólidos solúveis totais (SST) e açúcares solúveis totais (AST). Utilizou-se o delineamento experimental inteiramente casualizado, com quatro repetições. Aos dois dias, iniciou-se rápido aumento na RS, resultando no primeiro pico (197,60mg CO2·kg-1·h-1), que foi seguido por queda e uma fase lag. Ao término desta, iniciou-se o aumento climatérico (pico = 298,82mg CO2·kg-1·h-1). ET só foi detectada por ocasião do primeiro pico respiratório, atingindo o máximo aos quatro dias. A cor da casca tornou-se mais clara, enquanto na polpa se observou apenas redução da luminosidade. Houve coincidência entre o primeiro aumento na RS, o pico de ET e as mudanças mais significativas na firmeza, na ATT e nos teores de SST e AST. Durante o período, a firmeza diminuiu de 60 para 0,9N, e a ATT aumentou de 0,18 para 0,88% de ácido cítrico.
Termos para indexação:annonaceae, etileno, maturação, respiração.
ABSTRACT
It was aimed to evaluate physical and physico-chemical changes during soursop 'Morada' maturation at room temperature and related them to respiration rate and ethylene production. Fruits harvested at physiological maturity were stored (23,4 ± 1,1ºC and 81,8 ± 10,6% RH) and evaluated at 0, 1, 2, 3, 4, and 6 days, according to: respiration rate (RS), ethylene production (ET), fresh weight lost, skin and pulp color, firmness, pH, total titrable acidity (TTA), total soluble solids (TSS) and total soluble sugars (TS). The experimental design was a completely randomized one with four replications. There was a fast increase in RS at two days that resulted in the first peak (197,60mg CO2·kg-1·h-1) that was followed by a decay and a lag phase. When that phase rad finished, it began the climateric raise (climateric peak = 298,82mg CO2·kg-1·h-1). ET was detected only when the first respiratory peak occurred and reached the maximum at the fourth day. The skin color became bright green while it was observed a decrease just on lightness in the pulp. There was a coincidence among the first increase on RS, the peak of ET and the most significant changes on firmness, TAA, TSS content and TS content. During this period, firmness reduced from 60 to 0,9N and TTA increased from 0,18 to 0,88% of citric acid.
Index terms:annonaceae, ethylene, maturation, respiration.
INTRODUÇÃO
A graviola (Annona muricata L.) é um fruto climatérico, mas apresenta padrão de liberação de CO2 bastante distinto da maioria das espécies. Segundo Biale & Barcus (1970), a graviola possui um climatério difuso com mais de um ponto máximo.
Alguns estudos têm confirmado aquelas observações e destacado que o primeiro pico não corresponde ao climatério (Paull, 1982; Bruinsma & Paull, 1984; Worrell et al., 1994). As primeiras idéias atribuíram a existência dos dois picos às diferenças de desenvolvimento e amadurecimento entre os frutilhos que compõem o órgão, que é um fruto composto (Biale & Barcus, 1970). No entanto, Bruinsma & Paull (1984) argumentaram que a própria colheita induz ao aumento respiratório. Nesta ocasião, a respiração mitocondrial é incrementada pelo maior fornecimento de carboxilatos induzido pela separação do fruto da planta. Além disso, a produção de etileno geralmente só começa a ser detectada quando já ocorreu o primeiro pico de CO2 (Paull, 1982; Bruinsma & Paull, 1984; Worrell et al., 1994).
Colhendo-se o fruto na maturidade fisiológica, a atividade respiratória aumenta nos dois primeiros dias, a partir do qual se desencadeia a síntese de etileno (Bruinsma & Paull, 1984). Mudanças na cor, textura, sabor e aroma da maioria dos frutos durante o amadurecimento estão associadas ao climatério (Biale & Barcus, 1970), e algumas são dependentes de etileno (Ayub et al., 1996). Bruinsma & Paull (1984) e Worrell et al. (1994) registraram mudanças no desenvolvimento do flavor, no escurecimento da casca e no amolecimento da polpa durante a produção autocatalítica de etileno.
A perecibilidade e o reduzido tempo de vida da graviola (Aziz & Yusof, 1994) certamente estão associados à velocidade com que ocorrem as mudanças características do amadurecimento. Um maior entendimento deste metabolismo subsidiaria propostas de tecnologias para a conservação do fruto por períodos maiores.
A partir destas considerações, o objetivo deste estudo foi avaliar as alterações físicas e físico-químicas durante a maturação, sob temperatura ambiente, da graviola-'Morada', relacionando-as às taxas respiratória e de liberação de etileno.
MATERIAL E MÉTODOS
Frutos, do plantio comercial de gravioleira-'Morada' da Fazenda Alto da Boa Vista, localizada em São Luiz do Curu-CE, foram colhidos na maturidade fisiológica, selecionados com base na sanidade e na uniformidade de formato, e divididos nas épocas de avaliação a serem estudadas: 0; 1; 2; 3; 4 e 6 dias após a colheita. Até a data da avaliação, os frutos foram armazenados em temperatura ambiente, a 23,4 ± 1,1ºC e 81,8 ± 10,6% UR.
As variáveis estudadas e as metodologias empregadas foram as seguintes: a) atividade respiratória (mg CO2·kg-1·h-1), determinada em cromatógrafo a gás através de detector de condutividade térmica (150ºC), em coluna PORAPAK N (4 m x 3,2 mm), usando H2 como gás de arraste (30mL·min-1) a uma temperatura de 60ºC na coluna; b) liberação de etileno (mL·kg-1·h-1), determinada em cromatógrafo a gás através de detector de ionização de chama (200ºC), com as características descritas anteriormente; c) perda de matéria fresca (%), obtida através do peso individual do fruto no dia da colheita e na data da avaliação, utilizando-se de balança semi-analítica; d) cor da casca e da polpa, determinadas através de reflectômetro, a partir dos parâmetros luminosidade, cromaticidade e ºHue; e) firmeza da polpa (N), obtida por meio de texturômetro eletrônico, equipado com ponteira de 6 mm de diâmetro e ajustado para uma distância de penetração de 20 mm, à velocidade de 2,0 mm·s-1; f) pH, obtido em potenciômetro digital com eletrodo de membrana de vidro (IAL, 1985); g) acidez total titulável (ATT, % de ácido cítrico), determinada por titulação com solução de NaOH 0,1N, conforme metodologia do IAL (1985); h) sólidos solúveis totais (SST, ºBrix), determinados por leitura em refratômetro digital (IAL, 1985); i) açúcares solúveis totais (AST, %), extraídos em álcool 80% e determinados usando o reagente antrona, conforme Yemn & Willis (1954).
O delineamento experimental utilizado foi o inteiramente casualizado, com seis tratamentos e quatro repetições, exceto o tratamento correspondente aos 2 dias após a colheita, que teve três repetições. Cada unidade experimental foi constituída por um fruto.
As variáveis atividade respiratória e liberação de etileno foram avaliadas a intervalos de tempo mais freqüentes que as demais a fim de obter-se o padrão respiratório característico do fruto. As mesmas não foram submetidas à análise de variância por não atenderem ao critério de homocedasticidade das variâncias, conforme teste de Bartlett. Neste caso, apresentaram-se as médias das variáveis em cada tempo. Para as demais, procedeu-se às análises de variância e, quando os tratamentos foram significativos, às análises de regressão polinomial, admitindo-se equações de até 3° grau.
Os valores de perda de matéria fresca foram analisados após transformação em arc-sen Öx/100. Para representação gráfica, utilizou-se das médias não transformadas obtidas a partir da operação inversa à transformação.
RESULTADOS E DISCUSSÃO
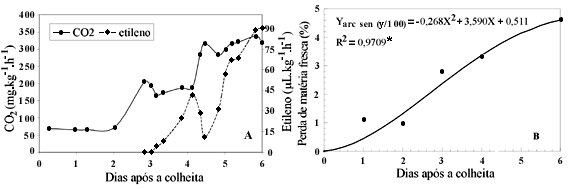
A atividade respiratória da graviola manteve-se estável em cerca de 70 mg CO2·kg-1·h-1 até dois dias após a colheita (Figura 1A). Nesta ocasião, começou um rápido aumento que resultou no primeiro pico respiratório e correspondeu à liberação de 197,60 mg CO2·kg-1·h-1.
Entre o terceiro e o quarto dia após a colheita, ocorreram poucas variações na atividade respiratória, caracterizando a fase lag, citada por Worrell et al. (1994). Ao término desta fase, o aumento respiratório seguinte resultou no pico climatérico propriamente dito (298,82 mg CO2·kg-1·h-1), observado entre o quarto e o quinto dia. Num intervalo de nove horas, a atividade respiratória caiu para 266,34 mg CO2·kg-1·h-1. A partir daí, observaram-se variações irregulares que sugerem o início da senescência do fruto.
O padrão respiratório obtido concorda com estudos anteriores (Biale & Barcus, 1970; Paull, 1982; Bruinsma & Paull, 1984; Worrell et al., 1994). Porém, a ocorrência do primeiro pico respiratório tem sido registrada desde o segundo até o quarto dia após a colheita, enquanto o pico climatérico pode ser observado do quarto ao sexto dia. Quanto aos valores, os autores citam atividade respiratória variando de 50 a 170 mg CO2·kg-1·h-1, no primeiro pico, e de 130 a 305 mg CO2·kg-1·h-1, no pico climatérico. Tais diferenças podem ser atribuídas aos estádios de maturação usados em cada estudo, aos materiais genéticos, aos métodos empregados e às condições a que os frutos foram submetidos, principalmente temperatura.
A liberação de etileno só começou a ser detectada aproximadamente aos três dias após a colheita, por ocasião do primeiro pico respiratório (Figura 1A). O pico (41,75 µL·kg-1·h-1) foi registrado no quarto dia, quando se iniciava o incremento respiratório para o pico climatérico, confirmando a idéia de que este aumento é uma resposta ao etileno (Dean & Matoo, 1991).
Da mesma forma que para a atividade respiratória, estudos têm registrado pico de liberação de etileno desde quatro até seis dias após a colheita, com taxas que variam de 80 até 350 µL·kg-1·h-1 (Paull, 1982; Bruinsma & Paull, 1984; Worrell et al., 1994).
A perda de matéria fresca aumentou durante o período, especialmente entre o segundo e o terceiro dia, até atingir 4,6% (Figura 1B). Entretanto, não repercutiu em sinais de enrugamento ou murcha de forma que não houve comprometimento da aparência. Mosca et al. (1997) obtiveram graviolas aptas para consumo mesmo após 11,8% de perda da matéria fresca.
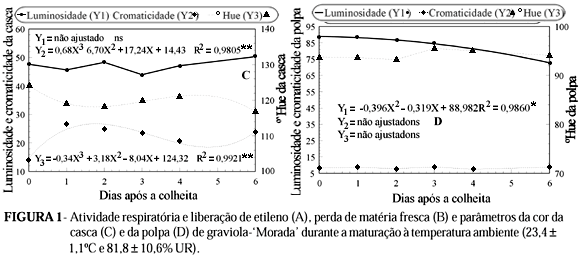
As variações na cor da casca foram devidas apenas à cromaticidade e ao ºHue já que não houve diferença estatisticamente significativa da luminosidade durante o período estudado (Figura 1C). Este resultado exclui o emprego da luminosidade como indicador seguro de maturidade em graviolas do tipo Morada. Contudo, na maioria dos fenótipos de graviola tem-se verificado aumento no brilho da casca durante a maturação (Salgado et al., 1998).
Por outro lado, as mudanças na cromaticidade e no ºHue asseguram diferenças de cor entre os frutos na maturidade fisiológica e os maduros (Figura 1C). Para cromaticidade, os valores não seguiram um padrão claro de resposta. É possível que a variabilidade natural da espécie dificulte uma caracterização precisa. Entretanto, a partir do primeiro dia após a colheita, os valores foram maiores do que no dia zero, o que sugere alguma degradação da clorofila.
O ºHue, que representa a variação de cor do verde para o amarelo, por sua vez, apresentou resposta mais característica (Figura 1C). A queda observada no primeiro dia em relação à data da colheita representou a fase de maior mudança na cor (de 124,4 para 118,9º). O período seguinte, até quatro dias após a colheita, pode ser descrito como estável. Somente após quatro dias é que ocorreu nova redução no ângulo, definindo uma cor verde mais clara aos seis dias. Mattiuz & Durigan (2001) também registraram aumento da cromaticidade e redução no ºHue em goiabas 'Paluma' e 'Pedro Sato' colhidas a partir da maturidade fisiológica.
Ao contrário da cor da casca, as variações na cor da polpa restringiram-se à luminosidade. Conforme Figura 1D, houve uma queda gradativa a partir dos dois dias após a colheita. Os valores de cromaticidade e ºHue tiveram variações pequenas, indicando que o amadurecimento da graviola não envolve mudanças expressivas na cor da polpa.
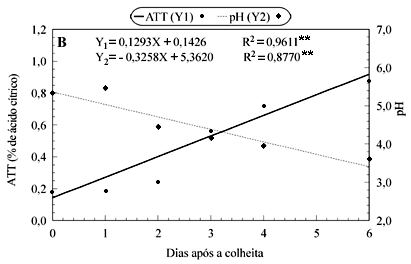
Durante o amadurecimento da graviola, a firmeza da polpa foi bruscamente reduzida (Figura 2A). Enquanto na maturidade fisiológica a firmeza era próxima de 60N, nos frutos maduros, este valor era de apenas 0,9N. As mudanças mais importantes ocorreram entre o segundo e o terceiro dia e a partir do quarto dia. Estas fases coincidiram com o primeiro aumento respiratório e com o pico de liberação de etileno, conforme Figura 1. Aos quatro dias após a colheita, quando a firmeza era de 19,1N, os frutos já podiam ser considerados moles.
Worrell et al. (1994) consideraram que o amolecimento da graviola começa com a liberação de etileno e que, no pico, todas as regiões do fruto já estão moles. Contudo, neste estudo, variações acentuadas na textura já eram observadas antes que a liberação de etileno tivesse sido desencadeada (Figuras 1A e 2A) e podem ser resultantes da hidrólise de poliuronídeos da parede celular, da perda de açúcares neutros (Mitcham & McDonald, 1992), da degradação de amido (Wills et al., 1998) ou perda de turgescência (Sams, 1999).
A ATT aumentou durante o período, partindo de 0,18 a 0,88% de ácido cítrico (Figura 2B). Outros estudos já destacaram este acúmulo na ATT, atípico em relação à maioria dos frutos (Wills et al., 1998), mas característico de algumas anonáceas (Paull, 1982; Bruinsma & Paull, 1984; Aziz & Yusof, 1994; Muñoz et al., 1997).
Considerando as médias dos valores originais não-ajustados, pode-se destacar o período de dois a três dias como o de maior incremento na ATT (0,32%). Este período coincidiu com o aumento respiratório e o primeiro pico de CO2, indicando que pode ser conseqüência da ativação da glicólise induzida pela colheita, com intensa oxidação de glicose e hidrólise de amido, conforme sugerido por Bruinsma & Paull (1984).
A coincidência entre o primeiro pico respiratório e o de liberação de etileno com as mudanças mais marcantes na ATT e na firmeza da polpa sugere que a atividade metabólica nesta fase promove degradação mais intensa de constituintes celulares, principalmente carboidratos, do que a verificada no climatério. É possível que, no primeiro pico de CO2, haja utilização reduzida de ácidos orgânicos nas vias oxidativas normais, favorecendo o acúmulo.
A ATT do fruto maduro (0,88% de ácido cítrico), aos seis dias após a colheita, é ligeiramente inferior àquela obtida por Paull (1982) e Cruz et al. (2000). É possível que as diferenças sejam de origem genética já que a graviola 'Morada', segundo Ramos (1999), tem sabor subácido a ácido, enquanto outros tipos podem variar de subácido a doce.
O pH, como conseqüência do aumento na ATT, diminuiu linearmente até os seis dias após a colheita (Figura 2B). A variação observada foi desde 5,46 a 3,60, nos frutos maduros. Estudos anteriores citam valores de pH, em graviola madura, variando de 4,2 a 3,7 (Paull, 1982; Aziz & Yusof, 1994; Mosca et al., 1997).
Em relação ao teor de SST, observou-se aumento até os quatro dias após a colheita (Figura 2C). Entre o quarto e o sexto dia, o incremento foi de apenas 0,9ºBrix, de forma que, no final do período de armazenamento, o teor de SST foi de 14,4ºBrix. Em relatos anteriores, alguns autores encontraram, em graviolas maduras, teores de SST variando de 10 a 18ºBrix (Paull, 1982; Aziz & Yusof, 1994; Mosca et al., 1997).
O maior acúmulo de SST ocorreu antes do aumento para o pico climatérico (Figura 1A). Portanto, a ocorrência do primeiro pico respiratório e do pico de etileno pode ter um envolvimento mais direto no ganho de SST, possivelmente por estimular reações que produzem precursores para compostos daquela natureza.
Durante o amadurecimento da graviola, conforme Figura 2C, o teor de AST aumentou linearmente, desde valores próximos de 5 até 9,98%, à semelhança do que obtiveram Aziz & Yusof (1994). A proporção de AST em relação aos SST, no entanto, variou pouco e correspondeu a 65-69% dos valores dos SST.
O acúmulo de açúcares durante o amadurecimento nos frutos climatéricos, ou mesmo em hortaliças que contenham reservas de amido, pode ser resultado da degradação deste carboidrato (Paull, 1982; Roe & Bruemmer, 1981) e possivelmente da quebra parcial de pectinas, celulose e outros polissacarídeos (Roe & Bruemmer, 1981).
CONCLUSÕES
1) A maturação da graviola-'Morada' caracterizou-se pela existência de dois picos respiratórios e pela liberação de etileno, detectada somente a partir do terceiro dia após a colheita.
2) A redução na cromaticidade da casca e na luminosidade da polpa indicaram que os frutos maduros tiveram cor mais clara e polpa menos brilhante que aqueles na maturidade fisiológica.
3) As modificações mais acentuadas na firmeza da polpa, na ATT e nos teores de SST e AST da graviola-'Morada' coincidiram com o período em que se verificou o primeiro aumento na respiração e o pico de liberação de etileno.
Recebido: 24/05/2002
Aceto para publicação: 13/02/2003
- AYUB, R.; GUIS, M.; AMOR, M.B.; GILLOT, L.; ROUSTAN, J.P.; LATCHÉ, A.; BOUZAYEN, M.; PECH, J.C. Expression of ACC oxidase antisense gene inhibits ripening of cantaloupe melon fruits. Nature Biotechnology, New York, v.14, p.862-866, 1996.
- AZIZ, P.A.; YUSOF, S. Physico-chemical characteristics of soursop fruit (Annona muricata) during growth and development. ASEAN Food Journal, New York, v.9, n.4, p.147-150, 1994.
- BIALE, J.B.; BARCUS, D.E. Respiratory patterns in tropical fruits of the Amazon basin. Tropical Science, London, v.12, n.2, p.93-104, 1970.
- BRUINSMA, J.: PAULL, R.E. Respiration during postharvest development of soursop fruit, Annona muricata L. Plant Physiology, Rockville, v.76, n.1, p.131-138, 1984.
- CRUZ, F.L. da; BARRETO, W.S.; MUNIZ, W. dos S.; DAMASCENO JÚNIOR, A.R.; OLIVEIRA, F.J. de; SACRAMENTO, C.K. do. Caracterização físico-química de três tipos de graviola (Annona muricata L.). In: CONGRESSO BRASILEIRO DE FRUTICULTURA, 16, 2000, Fortaleza. Resumos... Fortaleza: SBF, 2000. p.319. CD-ROM.
- DEAN, J.F.D.; MATTOO, A.K. The role of ethylene in fruit ripening and senescence. In: FOX, P.F. Food enzimology. London: Elsevier Applied Science, 1991, v.1, p.271-301.
- INSTITUTO ADOLFO LUTZ. Normas analíticas, métodos químicos e físicos para análise de alimentos. São Paulo: IAL, 1985. v.1, 371p.
- MATTIUZ, B.H.; DURIGAN, J.F. Efeito de injúrias mecânicas na firmeza e coloração de goiabas das cultivares Paluma e Pedro Sato. Revista Brasileira de Fruticultura, Jaboticabal, v.23, n.2, p.277-281, 2001.
- MITCHAM, E.J.; McDONALD, R.E. Cell wall modification during ripening of 'Keitt' and 'Tommy Atkins' mango fruit. Journal of the American Society for Horticultural Science, Alexandria, v.117, n.6, p.919-924, 1992.
- MOSCA, J.L.; ALVES, R.E.; FILGUEIRAS. H.A.C.; OLIVEIRA, J.F. de. Determination of harvest index for soursop fruits (Annona muricata L.) In: CONGRESO INTERNACIONAL DE ANONACEAS, I, 1997, Chapingo. Memorias... Chapingo: Universidad Autónoma Chapingo, p.315-322, 1997.
- MUÑOZ, M.T.; ESCRIBANO, M.I.; MERODIO, C. Ethanolic metabolism in cherimoya fruit during storage at ambient and under high CO2 atmosphere. The Journal of Horticultural Science, Ashford, v.72, n.3, p.363-370, 1997.
- PAULL, R.E. Postharvest variation in composition of soursop (Annona muricata L.) fruit in relation to respiration and ethylene production. Journal of the American Society for Horticultural Science, Alexandria, v.107, n.4, p.582-585. 1982.
- RAMOS, V.H.V. A potencialidade da gravioleira no cerrado. In: PINTO, A.C.Q. A cultura da graviola. Fortaleza: SINDIFRUTA/INSTITUTO FRUTAL, 1999, p. 42-58.
- ROE, B.; BRUEMMER, J.H. Changes in pectic substances and enzymes during ripening and storage of "Keitt" mangos. Journal of the Food Science, Chicago, v.46, n.1, p.186-189, 1981.
- SALGADO, F.R.; LÓPEZ, M.; GUTIÉRREZ, A. Manejo post-cosecha de la guanábana (Annona muricata L.). Santa Fé de Bogotá: NRI, SENA, DFID, 1998. 332p. (Serie de paquetes de capacitación sobre manejo post-cosecha de frutas y hortalizas, 8.)
- SAMS, C.E. Preharvest factors affecting postharvest texture. Postharvest Biology and Technology, Amsterdam, v.15, n.3, p.249-254, 1999.
- WILLS, R.; McGLASSON, B.; GRAHAM, D.; JOYCE, D. Postharvest: an introduction to the physiology & handling of fruit, vegetables & ornamentals. 4ª ed. Wallingford: UNSW Press, 1998. 262p.
- WORREL, D.B.; CARRINGTON, C.M.S.; HUBER, D.J. Growth, maturation and ripening of soursop. Scientia Horticulturae, Amsterdam, v.57, n.1, p.7-15, 1994.
- YEMN, E.W.; WILLIS, A.J. The estimation of carbohydrate in plant extracts by anthrone. The Biochemical Journal, London, v.57, p. 504-514, 1954.
Datas de Publicação
-
Publicação nesta coleção
17 Jul 2003 -
Data do Fascículo
Abr 2003
Histórico
-
Aceito
13 Fev 2003 -
Recebido
24 Maio 2002