Resumos
Rosellinia necatrix Prill induz a podridão branca da raiz da macieira, doença que causa severa perda em pomares localizados no sul do Brasil. O manejo da doença é principalmente preventivo e inclui o uso de porta-enxertos resistentes e a fumigação do solo. A proteção das mudas de macieiras antes do plantio com um isolado antagonista de Pantoea agglomerans foi recentemente proposto para reduzir a incidência da doença. Os objetivos desta pesquisa foram caracterizar o relacionamento entre o patógeno e a bactéria antagonista; a produção de metabólitos biológicamente ativos pelo isolado bacteriano e a sua ação sobre o patógeno; o efeito do pH, da temperatura e de carboxi-metil-celulose (CMC) sobre o crescimento do antagonista e do patógeno, isolados ou não e no controle da doença. Os resultados demonstraram que o crescimento de P. agglomerans foi maior em pH 5,5 e 6,0 e nas temperaturas de 20 °C e 30 °C. O crescimento micelial de R. necatrix foi inibido em meio de cultura e previamente colonizado pelo antagonista e com CMC nas concentrações de 0,25 e 0,5 %. A proteção das macieiras da infecção por R. necatrix foi observada quando utilizadas as concentrações de 10(7); 10(8) e 10(9) cel/mL e nas diferentes concentrações de CMC. Maior desenvolvimento radicular das macieiras foi constatado em todas as concentrações de CMC quando usada a concentração de 10(9) cel/mL. A formulação de P. agglomerans com CMC tornará possível a incorporação desta estratégia de controle ao manejo integrado da podridão branca das raízes da macieira no Sul do Brasil.
controle biológico; formulação; podridão de raízes
Rosellinia necatrix Prill induces the white root rot of apple, a disease that causes severe losses in orchards located in Southern Brazil. The management of the disease is mostly preventative and include usage of less susceptible rootstocks and soil fumigation. Protection of nursery apple trees before planting with an antagonistic bacterium Pantoea agglomerans was recently proposed to reduce the disease incidence. The aims of this research were to know about the pathogen-antagonic bacterium relationships and the effect of cell concentrations, pH, temperature and carboximethyl cellulose(CMC) in the culture media on the growth of the pathogen, the antagonic organism and on the disease control. Results showed that growth of P. agglomerans was improved on pH 5.5 to 6.0 , and at temperature of 20 °C and 30 °C. Mycelia of R. necatrix isolates was inhibited when grown in a media previously colonized by the antagonist and the two isolates were inhibited when in this media were added 0.25 % and 0.5 % of CMC in the culture media. Protection of apple plants from R. necatrix infection was observed with 10(7), 10(8) and 10(9) ufc/mL and different CMC concentrations. Root development was improved when cell concentration was 10(9) ufc/mL and when CMC was used. Formulation of P. agglomerans will make possible the use of biocontrol in the integrated management of the disease.
biological control; formulation; root rot
ARTIGOS
Desenvolvimento de Pantoea agglomerans em diversas temperaturas, pH e concentrações de carboxi-metil-celulose e o seu impacto no controle de Rosellinia necatrix
Development of Pantoea agglomerans under different temperatures, pH and carboximethyl cellulose rates and their effect on Rosellinia necatrix control
Valdirene Camatti-SartoriI; Rosa Maria Valdebenito-SanhuezaII; Rute Terezinha da Silva RibeiroI
ILaboratório de Controle Biológico de Doenças de Plantas, Instituto de Biotecnologia, Universidade de Caxias do Sul, CP 1352, 95001-970 – Caxias do Sul, Rio Grande do Sul
IIEmbrapa Uva e Vinho, CP130, 95700-000, Bento Gonçalves, Rio Grande do Sul
RESUMO
Rosellinia necatrix Prill induz a podridão branca da raiz da macieira, doença que causa severa perda em pomares localizados no sul do Brasil. O manejo da doença é principalmente preventivo e inclui o uso de porta-enxertos resistentes e a fumigação do solo. A proteção das mudas de macieiras antes do plantio com um isolado antagonista de Pantoea agglomerans foi recentemente proposto para reduzir a incidência da doença. Os objetivos desta pesquisa foram caracterizar o relacionamento entre o patógeno e a bactéria antagonista; a produção de metabólitos biológicamente ativos pelo isolado bacteriano e a sua ação sobre o patógeno; o efeito do pH, da temperatura e de carboxi-metil-celulose (CMC) sobre o crescimento do antagonista e do patógeno, isolados ou não e no controle da doença. Os resultados demonstraram que o crescimento de P. agglomerans foi maior em pH 5,5 e 6,0 e nas temperaturas de 20 °C e 30 °C. O crescimento micelial de R. necatrix foi inibido em meio de cultura e previamente colonizado pelo antagonista e com CMC nas concentrações de 0,25 e 0,5 %. A proteção das macieiras da infecção por R. necatrix foi observada quando utilizadas as concentrações de 107; 108 e 109 cel/mL e nas diferentes concentrações de CMC. Maior desenvolvimento radicular das macieiras foi constatado em todas as concentrações de CMC quando usada a concentração de 109 cel/mL. A formulação de P. agglomerans com CMC tornará possível a incorporação desta estratégia de controle ao manejo integrado da podridão branca das raízes da macieira no Sul do Brasil.
Palavras chave: controle biológico, formulação, podridão de raízes.
ABSTRACT
Rosellinia necatrix Prill induces the white root rot of apple, a disease that causes severe losses in orchards located in Southern Brazil. The management of the disease is mostly preventative and include usage of less susceptible rootstocks and soil fumigation. Protection of nursery apple trees before planting with an antagonistic bacterium Pantoea agglomerans was recently proposed to reduce the disease incidence. The aims of this research were to know about the pathogen-antagonic bacterium relationships and the effect of cell concentrations, pH, temperature and carboximethyl cellulose(CMC) in the culture media on the growth of the pathogen, the antagonic organism and on the disease control. Results showed that growth of P. agglomerans was improved on pH 5.5 to 6.0 , and at temperature of 20 °C and 30 °C. Mycelia of R. necatrix isolates was inhibited when grown in a media previously colonized by the antagonist and the two isolates were inhibited when in this media were added 0.25 % and 0.5 % of CMC in the culture media. Protection of apple plants from R. necatrix infection was observed with 107, 108 and 109 ufc/mL and different CMC concentrations. Root development was improved when cell concentration was 109 ufc/mL and when CMC was used. Formulation of P. agglomerans will make possible the use of biocontrol in the integrated management of the disease.
Additional keywords: biological control, formulation, root rot.
Estudos feitos no Rio Grande do Sul mostram que as perdas de plantas de macieira variam de 0,5 a 12 % ao ano, em decorrência de infecções causadas pelas podridões de raízes. A podridão causada por R. necatrix é de difícil controle, uma vez que as práticas disponíveis são pouco eficazes (10). Esta situação compromete a rentabilidade das propriedades. Poucos são os trabalhos de estudo das condições propícias para o desenvolvimento de R. necatrix, que No estudo deste patógeno Agarwala & Sharma (1) é favorecido por temperaturas de 14 a 17 °C e pH 6 .
As recomendações de controle da podridão branca da raiz da macieira incluem: o uso de mudas de um porta-enxerto resistente, a aplicação do brometo de metila no replantio e o tratamento das raízes das plantas com um fungicida do grupo dos benzimidazóis, medidas que têm apresentado eficiência variável. Nas áreas de replantio de macieiras, tratadas com os métodos hoje recomendados, somente é possível retardar o surgimento da doença, fato que torna o replantio das macieiras economicamente inviável. O controle biológico de R. necatrix foi obtido com a utilização de estirpes de Trichoderma spp. incorporadas ao solo contaminado, associando-se esta prática à solarização (4, 17). Outra estratégia de controle da doença recentemente proposto foi o tratamento das raízes das mudas com um isolado de Pantoea agglomerans com 1 % carboxi-methil-celulose (22, 23).
Estudos sobre P. agglomerans mostraram que isolados desta espécie utilizam diversas fontes de carbono, destacando-se a glicose, a sacarose e produzem exopolissacarídeos em meio de cultura e sideróforos, na presença de sacarose, ácido nicotínico, citrato férrico (1, 5). Não foram encontrados na bibliografia, resultados sobre o uso de carboxi-metil-celulose (CMC) no desenvolvimento de P. agglomerans.
O presente trabalho objetivou verificar os efeitos do pH, da temperatura, das diferentes concentrações de células de P. agglomerans e de CMC, em Rosellinia necatrix, na relação antagonista-patógeno e no controle da podridão branca das macieiras.
MATERIAL E MÉTODOS
Microganismos
Foram utilizados dois isolados de R. necatrix obtidos de raízes de macieira, sendo o isolado FC proveniente de um pomar do Município de Flores da Cunha, RS e o isolado LV de um pomar do Município de Lagoa Vermelha, RS. O isolado antagonista de Pantoea agglomerans(10.2), foi obtido de raízes sadias de macieira. A manutenção das culturas dos microrganismos foi feita no meio BDA (batata - 200 g., dextrose - 20g; ágar, 20g e água destilada -1 L) A bactéria antagonista foi multiplicada em vidros com o meio BD (batata - 200 g; dextrose – 20 g e água destilada – 1 L) submetidos à agitação (150 rpm). Todos os microrganismos utilizados pertencem á coleção da Embrapa Uva e Vinho (CNPUV - Bento Gonçalves, RS).
Efeito da temperatura sobre o desenvolvimento de P. agglomerans.
Foram realizados dois experimentos. No primeiro, vidros com o caldo BD e a bactéria foram incubados por 24 h na temperatura de 10, 20, 30 e 40 °C. No segundo, aos tratamentos antes citados se acrescentou a incubação nas temperaturas a 30 ºC e 37 ºC. Ao final dos experimentos, alíquotas das culturas foram transferidas para placas de Petri com BDA e após a incubação foi determinado o número de colônias desenvolvidas. Cada tratamento constou de três repetições constituídas por uma placa.
Efeito do pH sobre o desenvolvimento do antagonista e do patógeno.
O antagonista foi cultivado em frascos com meio BD com pH : 5,0; 5,5 e 6,0, corrigido com HCl. Os cultivos foram mantidos, por 24 h a 27 °C e a avaliação, feita pela medida da absorbância em 570 nm. Cada tratamento foi feito com 3 repetições. No caso de R. necatrix, um disco de 5 mm de diâmetro de ágar, colonizado com os isolados FC ou LV foi cultivado em placas de Petri com meio BDA com pH 5,0; 5,5 ou 6,0 e incubado durante 72 h à 25 °C. O experimento teve cinco repetições e na avaliação registrou-se o diâmetro das colônias.
Fracionamento e efeito de metabólitos liberados por P. agglomerans sobre Rosellinia necatrix.
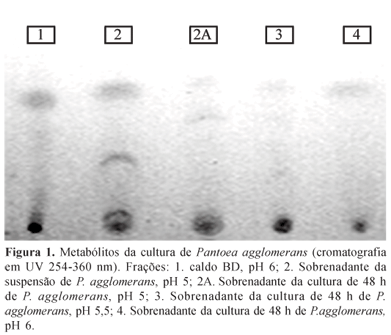
O processo foi feito a partir do sobrenadante de culturas de P. agglomerans. Foi usado caldo de cultura BD com pH 5,0; 5,5 ou 6,0 e o tempo de incubação variou de 0 a 48 h. A testemunha foi apenas caldo BD. As culturas foram centrifugadas a 15.000 g por 30 minutos e a biomassa foi descartada. Os sobrenadantes foram esterilizados por filtração à vácuo em membrana Millipore 0,22 µm e colocados no extrator contínuo com solvente diclorometano por 6 h. A seguir cada extrato foi concentrado em rotavapor rotativo e submetido a cromatografia de camada delgada de fluxo contínuo, realizada em cromatofolhas de alumínio contendo sílica gel 60 GF254. Para a corrida em placa de cromatografia foram utilizados os solventes hexano e diclorometano na proporção de 1:1. A visualização das manchas foi feita sob lâmpada UV 254-360 nm.
Cada mancha cromatográfica foi recortada e colocada próxima a um disco de ágar de 0,5 cm de diâmetro colonizado por um dos isolados de R. necatrix, em placa de Petri com BDA. Após 72 h foi medido o diâmetro das colônias do fitopatógeno. Para cada isolado de R. necatrix e mancha cromatográfica foram feitas três repetições.
Efeito in vitro do caldo de cultura de Pantoea agglomerans sobre Rosellinia necatrix.
A bactéria antagônica foi cultivada em caldo BD com concentrações crescentes de CMC (0,0; 0,25, 0,5; 1 e 1,5 %), em frascos agitados por 24 h a 150 rpm e 27 °C. A seguir, as culturas foram centrifugadas por 30 min a 15.000 g. Os pellets foram descartados e os sobrenadantes foram esterilizados por filtração à vácuo com papel Whatman n° 1 e membrana bacteriológica Millipore de 0,22 µm. A cada um dos filtrados foi acrescentado ágar fundente (40 °C) e em seguida, a mistura foi distribuída em placas de Petri. Na superfície dos meios de cultivo foram colocados 4 discos de 0,7 cm de diâmetro dos isolados FC e LV de R. necatrix . As placas foram mantidas por 3 dias na temperatura de 25 °C. Cada tratamento constou de três repetições e, na avaliação, foi registrado o tamanho das colônias. O controle foi constituído por placas de Petri com caldo BD sem colonizar.
Efeito das concentrações de CMC e de Pantoea agglomerans no desenvolvimento de Rosellinia necatrix.
Foram preparados três conjuntos de suspensões de CMC em caldo BD nas concentrações 0,0; 0,25; 0,5; 1 e 1,5 %. O primeiro deles foi mantido sem P. agglomerans; ao segundo foi transferida uma suspensão de células de P. agglomerans e, ao último foi acrescentado uma cultura de P. agglomerans com 24 h de incubação a 24 °C. Alíquotas destes meios foram distribuídas sobre meio BDA contido em placas de Petri. Quatro discos de 0,7 cm de meio BDA colonizado pelos isolados LV ou LF de R. necatrix, foram colocados na superfície dos meios em posições eqüidistantes. Foram utilizadas duas repetições para cada isolado. Após 72 h a 25 °C e na ausência de luz, foi medido o diâmetro das colônias do fitopatógeno.
Controle biológico de Rosellinia necatrix nas macieiras
A bactéria P. agglomerans foi multiplicada em caldo BD e pH 6,0. A concentração das suspensões bacterianas foi feita pela determinação de absorbância a 570 nm, sendo os valores previamente definidos por uma curva de concentração e absorbância. O patógeno foi multiplicado em segmentos de ramos de macieira (1 x 1 cm) autoclavados por 1 h, dois dias consecutivos e, posteriormente colonizados com discos de BDA (5 mm) com R. necatrix (19). O plantio das mudas do porta-enxerto MM 106 foi feito em caixas de madeira (25 x 30 x 17 cm) com solo sem esterilizar. Um segmento de ramo, colonizado pelo patógeno, foi disposto em 4 covas de 15 cm de profundidade, 15 dias antes do plantio das mudas. As raízes das plantas foram imersas durante 30 min nas suspensões com diferentes concentrações de propágulos do isolado antagônico de P. agglomerans (107, 108 e 109 ufc/mL) sem e com CMC nas concentrações 0,0; 0,25; 0,5; 1,0 e 1,5 %. A testemunha recebeu apenas o fitopatógeno. As plantas foram mantidas em casa de vegetação por 15 dias e posteriormente transferidas para um telado.
O delineamento experimental foi inteiramente casualizado e os tratamentos constaram das combinações entre a infestação do solo com os dois isolados de R. necatrix, nas diferentes concentrações da bactéria antagônica e as de CMC. Cada tratamento foi feito com duas repetições, cada uma constituída por uma caixa com quatro plantas. A avaliação foi feita após 270 dias e as variáveis avaliadas foram: matéria seca da raiz; diâmetro do colo a 2 cm acima do nível do solo e número de plantas sadias e mortas. Foi aplicado o fungicida difenoconazole para controle de oídio, nas doses recomendadas pelo fabricante. Os dados obtidos foram submetidos a análise de variância e à comparação das médias foi feita com o teste de Duncan (p<0,05).
RESULTADOS E DISCUSSÃO
Efeito da temperatura sobre o desenvolvimento de P. agglomerans.
O desenvolvimento de P agglomerans nas temperaturas de 10 °C; 20 °C; 30 °C; 35 °C, medido pela absorbância das suspensões foi de 0,03; 0,217; 0,289; 0,161 e de 0,018 nm, respectivamente. O maior estímulo das temperaturas no crescimento bacteriano, citados em ordem decrescente ocorreu sob 30 °C, 20 °C e 35 °C(p<0,05%). Nas temperaturas de 10 °C e 40 °C detectaram-se as absorbâncias igualmente menores e a absorvância obtida à 40°C foi igual à da suspensão inicial da cultura. Estes resultados são semelhantes aos obtidos por outros autores (5, 8, 26) que obtiveram bom crescimento de P agglomerans entre 18 e 37 °C. Contudo, neste trabalho quando a cultura do isolado antagonista ora avaliado foi feita em meio sólido, após 24 h de incubação a 37 °C, não foi verificado nenhum crescimento da bactéria.
Efeito do pH sobre o desenvolvimento do patógeno e do antagonista.
O desenvolvimento de colônias de P. agglomerans foi igualmente maior (p<0,05%) nos pHs 5,5 e 6,0 (0,34 e 0,36 cm, respectivamente), sendo constatado o menor tamanho de colônia no pH 5,0. O impacto dos três valores de pH sobre R. necatrix foi diferente dependendo do isolado avaliado. Assim, enquanto o isolado FC não foi afetado por nenhum dos valores de pH testados, o isolado LV apresentou colônias de maior tamanho em pH 5,5 atingindo 2,24 cm, o menor no pH 5,0 (1,86 cm) e colônias de tamanho intermediário(2,0 cm) no pH 6,0 (p<0,05%). Outros trabalhos relataram que o maior desenvolvimento deste patógeno ocorre em meio de cultura com valores de pH que oscilam entre 6,0 e 7,0 (4, 11, 19). Não foram encontrados relatos de comparações do efeito do pH do meio de cultura no desenvolvimento de R. necatrix.
Na literatura, cultivos em meios com pH em torno de 5,0 e 7,0 são citados nos experimentos com P. agglomerans, não havendo contudo trabalhos de comparações do efeito de diferentes valores deste fator na bactéria (7, 8, 12, 14, 26). Visto que as condições favoráveis para o desenvolvimento P. agglomerans foram pH 5,5 e 6,0, se sugere que, maior vantagem competitiva para P. agglomerans atuar como antagonista ocorrerá em solos de pomares com pH corrigido, como é atualmente recomendado para a cultura da macieira, (3 , 19).
Efeito in vitro das frações dos metabólitos obtidos do caldo de cultura de P. agglomerans e do CMC sobre o desenvolvimento de R. necatrix.
Foram constatadas 5 frações no caldo de cultura de P. agglomerans e nenhuma delas inibiu o tamanho das colônias do isolado LV de R. necatrix o que apresentou colônias com tamanho de 1,56 a 1,63 cm de diâmetro (Figura 1). No isolado FC deste patógeno, somente pela fração 2 A teve efeito inibitório do tamanho das colônias (p<0,05) a que atingiu 1,66 cm de diâmetro. Não se observou efeito das frações 3 e 1 (1,8 e 1,83 cm, respectivamente) e as frações 2 e 4, apresentaram efeito intermediário nas colônias (1,76 cm de diâmetro).
Os resultados obtidos mostraram a variação dos impactos que a produção de metabólitos pode ter em diferentes estirpes do mesmo patógeno o que confirma a necessidade de antagonistas competitivos apresentarem uma diversidade de mecanismos de ação para se tornarem confiáveis como ferramenta de controle de fitopatógenos.
Não foi constatada variação do tamanho das colônias de R. necatrix. na ausência de CMC, tanto na presença de culturas do antagonista recém preparadas como nas com 24 h de incubação (Tabela 1). No entanto na presença de 0,25 % de CMC e células da bactéria antagonista, ocorreu forte inibição, que foi ainda maior quando foi utilizada a cultura bacteriana com 24 h de idade (Duncan p<0,05). Na concentração 0,5 % de CMC o menor desenvolvimento de R. necatrix ocorreu com a cultura com 24 h de incubação. Finalmente, na presença de 1,0 e 1,5 % de CMC, não foi observado nenhum efeito da presença da bactéria antagonista, confirmando-se que nas concentrações menores da CMC são mais adequadas para estimular a ação antagônica de P. agglomerans a que deverá atuar como suporte para o antagonista sem se constituir em substrato para o patógeno.
Na avaliação do biocontrole de R. necatrix, em casa de vegetação, foi observado quando inoculada uma suspensão de 10 9cel/mL de P. agglomerans as plantas desenvolveram maior massa de raízes em todos os tratamentos com CMC o que não ocorreu com as concentrações menores das suspensões com o antagonista((Tabela 2).
Os dados obtidos sugerem que P. agglomerans deve ter vários mecanismos de ação no biocontrole de R.necatrix e na redução da doença, atuando como antagonista com ação direta de inibição sobre o patógeno, e, na presença de CMC estimulando os mecanismos de defesa no cultivar MM106 de porta-enxerto de macieira pela compensação de massa radicular e provávelmente, competindo com R. necatrix por nutrientes e/ou por sítio de infecção.
Controle biológico de R. necatrix por formulações de P. agglomerans
Todas as concentrações da bactéria antagônica reduziram a mortalidade das macieiras por R. necatrix e promoveram aumento do diâmetro do caule das plantas (Tabela 3). Adicionalmente, no tratamento com 1x108 ufc/mL do antagonista foi detectado aumento da matéria seca das raízes. Linhagens de P. agglomerans, estudadas por outros autores têm apresentado capacidade de formar associações endofíticas e epifíticas com suas hospedeiras, protegendo-as. Este efeito foi relacionado à atividade fitohormonal e habilidade de estirpes desta espécie em reduzir o nitrogênio (16). A promoção de crescimento das plantas por diversas espécies de rizobactérias nas quais P. agglomerans se inclui, é um efeito já conhecido sendo este grupo de organismos citados como PGPR (2, 18).
Os benefícios da formulação da bactéria antagônica com 0,25 a 0,5 % de CMC serão provavelmente os de assegurar a homogeneização da suspensão de células do antagonista, estabelecer uma barreira física entre o hospedeiro e o patógeno e estimular o tamanho do sistema radicular das macieiras melhorando a resistência do porta-enxerto MM 106. A bioformulação com CMC poderá conferir ao isolado antagônico de P. agglomerans, uma vantagem competitiva sobre a microflora do solo por permitir maior adesão e atividade da bactéria junto às raízes se constituindo num fator importante para a otimização do biocontrole de R. necatrix na macieira
Desta forma espera-se que o tratamento das plantas com o isolado de P. agglomerans, agente de biocontrole de R necatrix, poderá representar uma alternativa para complementar o manejo integrado da podridão branca das raízes da macieira na região Sul do Brasil.
Data de chegada:20/06/2005.
Aceito para publicação em: 07/05/2007
Parte da Dissertação de Mestrado do primeiro autor
Autor para correspondência: Rosa Maria Valdebenito Sanhueza
- 1.Agarwala, R. K.; Sharma, V. C. White root rot disease of apple in Himalachal Pradesh. Indian Phytopathology, New Delhi, v. 19, n. 1, p. 82-86, 1966.
- 2. Amellal, N.; Burtin, G.; Bartoli, F.; Heulin, T. Colonization of wheat roots by an exopolysaccharide producing by a Pantoea agglomerans strain and its effect on rhizosphere soil aggregation. Applied and Environmental Microbiology, Washington, v. 64, n. 10, p. 3740-3747, 1998.
- 3. Bleicher, J.; Melzer, R.; Berton, O.; Bonetti, J. I. S.; Driessen, C. Doenças da macieira. In: Empresa Catarinense Agropecuária S. A. Manual de Cultura da Macieira Florianópolis, 1989.
- 4. Feistner, G. J.; Ishimaru, C. Proferrioxamine profiles of Erwinia herbicola and related bacteria. BioMetals, London, v. 9, n. 4, p. 337-344, 1996.
- 5. Freeman, S.; Sztejnberg, A. Rosellinia In: Singleton, L. L.; Mihail, J. D.; Rush, C. M. Methods for research on soilborne phytopathogenic fungi Saint Paul: APS, 1992. p.71-73.
- 6. Gavini, F.; Mergaert, J. Beji. A, Mielcarek, C.; Izard, D.; Kersters, K. Transfer of Enterobacter agglomerans (Beijerinck 1888) Ewing and Fife 1972 to Pantoea new genus as Pantoea agglomerans new combination and description of Pantoea dispersa International Journal of Systematic Microbiology, Reading, v. 39, n. 3, p. 337-345, 1989.
- 7. Kempf, H. J.; Bauer, P. H.; Schroth, M. N. Herbicolin A associated with crown and roots of wheat after seed treatment with Erwinia herbicola B247. Phytopathology, Saint Paul, v. 83, n. 2, p.213-216, 1993.
- 8. Klopmeyer, M. J.; Ries, S. M. Motilily and chemotaxis of Erwinia herbicola and its effect on Erwinia amylovora Phytopathology, Saint Paul, v. 77, n. 2, p. 909-914, 1987.
- 9. Loon Van, L. C.; Bakker, P. A. H. M.; Pieterse, C. M. J. Systemic resistance induced by rhizosphere bacteria. Annual Review of Phytopathology, Palo Alto, v. 36, p. 453-83, 1998.
- 10. Mc Cormack, P. J.; Wildman, H. G.; Jeffries, P. Production of antibacterial compounds by phylloplane inhabiting yeasts and yeastlike fungi. Applied and Environmental Microbiology, Washington, v. 60, n. 3, p. 927-931, 1994
- 11. Melo, E .M. P.; Ferraz, J. F. P. Influência de várias fontes de carbono no crescimento de Rosellinia necatrix Revista de Ciências Agrárias, Belém, v. 1, n. 3/ 4, p. 1-4, 1990.
- 12. Montesinos, E.; Bonaterra, A.; Ophir Y. ; Beer, S. V. Antagonism of selected bacterial strains to Stemphylium vesicarium and biological control of brown spot of pear under controlled environment conditions. Phytopathology, Saint Paul, v. 86, n. 8, p. 856-863, 1996.
- 13. Nicholson, S. L.; Sigee, D. C.; Epton, H. A. S. Biological control of fire blight of perry pear: comparative evaluation of antagonists on immature fruit slices, micropropagated schoots and orchard blossom. Acta Horticulturae, The Hague, n. 273, p. 397, 1990.
- 14. Parker, L. W.; Rathnum, L. M.; Wells, Jr. J. S. SQ 27.860, a simple carbapenem by species of Serratia and Erwinia Journal of antibiotics, Tokyo, v. 35, n. 6, p. 653-661, 1982.
- 15. Ruppel, S.; Buchholz, C. H.; Remus, R.; Ortmann, U.; Schmelzer, R. Settlement of the diazotrophic, phytoeffective bacterial strain Pantoea agglomerans on and within winter wheat: An investigation using ELISA and transmission eletron microscopy. Plant and Soil, The Hague, v. 145, n. 2, p. 261-273, 1992.
- 16. Scholzseidel, C.; Ruppel, S. Nitrogenase and phytohormone activities of Pantoea agglomerans in culture and their reflection in combination with wheat plants. Zentralblatt fur Mikrobiologie, Jena, v. 147, n. 5, p. 319-328, 1992.
- 17. Sztejnberg, A.; Freeman, S.; Chet, I.; Katan, J. Control of Rosellinia necatrix in soil in apple orchard by solarization and Trichoderma harzianum Plant disease, Saint Paul, v. 71, n. , p. 365-369. 1987.
- 18. Sturz, A. V.; Christie, B. R.; Matheson, B. G. Nowak, K. Biodiversity of endophytic bacteria which colonize red clover nodules, roots, stems and foliage and influence on host growth. Biology and Fertility of Soils, Berlin, v. 25, n. 1, p. 13-19, 1997.
- 19. Tourvieille de Labrouhe, D. Pénétration de Rosellinia necatrix (Hart.) Berl. dans les racines du pommier en conditions artificielle. Agronomie, Paris, v. 2, n. 6, p. 553-560, 1982.
- 20. Valdebenito-Sanhueza, R. M. Podridão de raízes da macieira. Pelotas: EMBRAPA-CNPFT, 1988. 16 p. (EMBRAPA-CNPFT. Documentos, 32).
- 21. Valdebenito-Sanhueza, R. M. Podridão das raízes das fruteiras temperadas por Rosellinia necatrix (Dematophora necatrix) Bento Gonçalves: EMBRAPA-CNPUV, 1997. 12 p. (Circular Técnica, 22).
- 22. Valdebenito-Sanhueza, R. M. Controle integrado de Rosellinia necatrix em macieira Bento Gonçalves: EMBRAPA-CNPUV, 1998. Não paginado. Sub-Projeto de Pesquisa – 05.0.94.001.11.
- 23. Valdebenito- Sanhueza, R.M. Biocontrole de Rosellinia necatrix com rizobactérias em macieiras. Summa Phytopathologica, Jaboticabal.v. 27, p. 43-49, 2001.
- 24. Vanneste, J. L. Honey bees and epiphytic bacteria to control fire blight, a bacterial disease of apple and pear. Biocontrol News and Information, London, v. 17, n. 4 (67N-78N), 1996.
- 25. Weller, D. M. Biological control of soiborne plant pathogens in the rhizosphere with bacteria. Annual Review of Phytopathology, Palo Alto, v. 26, p.379-407, 1988.
- 26. Winkelmann, G.; Lupp, R.; Jung, G. Herbicolins – New peptide antibiotics from Erwinia herbicola Journal of antibiotics, Tokyo, v. 33, n. 4., p.353-358, 1980.
Datas de Publicação
-
Publicação nesta coleção
26 Maio 2008 -
Data do Fascículo
Fev 2008
Histórico
-
Recebido
20 Jun 2005 -
Aceito
07 Maio 2007