Resumos
O gênero Calliandra Benth. (Leguminosae) possui 134 espécies exclusivamente neotropicais, das quais 42 ocorrem na Chapada Diamantina, sendo 32 endêmicas da área. O objetivo deste trabalho foi avaliar o efeito da luz e substrato na germinação e desenvolvimento inicial de C. viscidula e C. hygrophila. As sementes foram submetidas a três tratamentos em laboratório: luz, escuro e luz/escuro (12 h/12 h). Em viveiro foram utilizados três substratos: (I) areia; (II) terra vegetal + areia; (III) vermiculita + terra vegetal + areia, mantidos a 100%; e 70% de luminosidade. As avaliações da germinação foram diárias durante 21 dias, utilizando-se os parâmetros: germinabilidade, índice de velocidade de germinação (índice de velocidade de emergência para viveiro), tempo médio de germinação e frequência relativa. Os parâmetros analisados no desenvolvimento inicial foram: comprimentos da parte aérea e raiz, número de folhas, massa seca, relação parte aérea/sistema radicular e razão do peso da folha. C. viscidula e C. hygrophila foram classificadas como fotoblásticas neutras e apresentaram melhor desenvolvimento a 70% de luminosidade nos substratos III e I e II , respectivamente.
Propagação; Fotoblastismo e Crescimento inicial
The genus Calliandra Benth. (Leguminosae) has 134 exclusively Neotropical species; 42 species are distributed in the Chapada Diamantina and 32 are endemics to this area. The aim of this work was to evaluate the effect of light and growing media on germination and initial seedling development of the Calliandra viscidula and C. hygrophila. Seeds were placed under three different light conditions in laboratory conditions: light, dark, and light/dark (12h/12h). In nursery conditions, seeds were germinated in three different growing medias: (I) sand; (II) soil + sand; (III) vermiculite + soil + sand and they were submitted to 100% and 70% of natural light conditions. Evaluations were daily for 21 days for germinability, speed germination index (speed emergency index to nursery), average germination time and relative frequency. The variables evaluated during initial seedling development were: shoot and root lengths, leaf numbers, dry weight, shoot/root system, and leaf weight ratio. C. viscidula and C. hygrophila are photoblastically neutral and have better results to the 130 days under 70% light condition. The best development was achieved when seeds were grown in growing media III for C. viscidula and I and II for C. hygrophila.
Propagation; Photoblastism and Early growth1
Influência da luz e substrato na germinação e desenvolvimento inicial de duas espécies de Calliandra Benth. (Mimosoideae - Leguminosae) endêmicas da Chapada Diamantina, Bahia
Influence of the light and growing media on germination and initial seedling development of two species of the Calliandra Benth. (Mimosoideae - Leguminosae) endemics of the Chapada Diamantina, Bahia
Sheila Vitória Resende; Iara Cândido Crepaldi; Claudinéia Regina Pelacani; Alone Lima Brito
Universidade Estadual de Feira de Santana, UEFS, Brasil. E-mail: <svresende@yahoo.com.br>, <iaracrepaldi@ig.com.br>, <pelacani@uefs.br> e <lima_brito@yahoo.com.br>
RESUMO
O gênero Calliandra Benth. (Leguminosae) possui 134 espécies exclusivamente neotropicais, das quais 42 ocorrem na Chapada Diamantina, sendo 32 endêmicas da área. O objetivo deste trabalho foi avaliar o efeito da luz e substrato na germinação e desenvolvimento inicial de C. viscidula e C. hygrophila. As sementes foram submetidas a três tratamentos em laboratório: luz, escuro e luz/escuro (12 h/12 h). Em viveiro foram utilizados três substratos: (I) areia; (II) terra vegetal + areia; (III) vermiculita + terra vegetal + areia, mantidos a 100%; e 70% de luminosidade. As avaliações da germinação foram diárias durante 21 dias, utilizando-se os parâmetros: germinabilidade, índice de velocidade de germinação (índice de velocidade de emergência para viveiro), tempo médio de germinação e frequência relativa. Os parâmetros analisados no desenvolvimento inicial foram: comprimentos da parte aérea e raiz, número de folhas, massa seca, relação parte aérea/sistema radicular e razão do peso da folha. C. viscidula e C. hygrophila foram classificadas como fotoblásticas neutras e apresentaram melhor desenvolvimento a 70% de luminosidade nos substratos III e I e II , respectivamente.
Palavras-chave: Propagação, Fotoblastismo e Crescimento inicial.
ABSTRACT
The genus Calliandra Benth. (Leguminosae) has 134 exclusively Neotropical species; 42 species are distributed in the Chapada Diamantina and 32 are endemics to this area. The aim of this work was to evaluate the effect of light and growing media on germination and initial seedling development of the Calliandra viscidula and C. hygrophila. Seeds were placed under three different light conditions in laboratory conditions: light, dark, and light/dark (12h/12h). In nursery conditions, seeds were germinated in three different growing medias: (I) sand; (II) soil + sand; (III) vermiculite + soil + sand and they were submitted to 100% and 70% of natural light conditions. Evaluations were daily for 21 days for germinability, speed germination index (speed emergency index to nursery), average germination time and relative frequency. The variables evaluated during initial seedling development were: shoot and root lengths, leaf numbers, dry weight, shoot/root system, and leaf weight ratio. C. viscidula and C. hygrophila are photoblastically neutral and have better results to the 130 days under 70% light condition. The best development was achieved when seeds were grown in growing media III for C. viscidula and I and II for C. hygrophila.
Keywords: Propagation, Photoblastism and Early growth1.
INTRODUÇÃO
O gênero Calliandra Benth. (Mimosoideae - Leguminosae) possui espécies exclusivamente neotropicais de arbustos ou arvoretas (BARNEBY, 1998), muitas das quais utilizadas com fins ornamentais. Seu rápido crescimento somado à densa folhagem e ao sistema radicular profundo indicam que essas espécies sejam adequadas para uso no controle da erosão e na restauração e melhoria do solo de ambientes degradados, favorecendo o estabelecimento de outras espécies vegetais (CHAMBERLAIN, 2000; LORENZI e SOUZA, 2001).
A Chapada Diamantina é um dos principais centros de diversidade de Calliandra; das 134 espécies existentes no gênero, 42 ocorrem nessa região, sendo 32 espécies endêmicas da área (SOUZA, 2001). A espécie Calliandra viscidula Benth possui ampla distribuição por toda essa região, enquanto Calliandra hygrophila Mackinder e Lewis é restrita à serra do Sincorá, situada na região central do Estado da Bahia, ocorrendo nas margens de rios (SOUZA, 2001).
O uso de espécies nativas com enfoque na recuperação de áreas degradadas vem sendo intensificado, visto que essas espécies supostamente são bem mais adaptadas às condições edafoclimáticas, o que facilitaria o restabelecimento do equilíbrio entre a fauna e a flora local (MORAES NETO et al., 2000; ALVARENGA et al., 2003).
Apesar do aumento considerável dos estudos sobre a análise de sementes de espécies nativas, muitas ainda carecem de informações básicas referentes às condições ideais de germinação e crescimento inicial, principalmente quanto ao efeito da luz no controle desse processo (ANDRADE et al., 2000; BORGHETTI, 2000; FONSECA et al., 2006). O conhecimento dos principais processos envolvidos na germinação de sementes de espécies nativas é de grande importância na preservação e utilização dessas em programas de recomposição de áreas degradadas (SMIDERLE e SOUSA, 2003; LIMA et al., 2006).
Alguns fatores como a luz e as condições edáficas desempenham importante papel no controle da germinação das sementes e do desenvolvimento inicial. Apesar de não atuar isoladamente sobre as plantas, a luz desempenha papel fundamental no controle desses processos. (PEDROSO e VARELA, 1995; SILVA et al., 1997; UCHIDA e CAMPOS, 2000; PORTELA et al., 2001; ALVARENGA et al., 2003).
Diferentes níveis de luminosidade causam mudanças fisiológicas e morfológicas na planta, e o seu sucesso depende da sua adaptação a esses diferentes níveis, o que está relacionado com as características genéticas e sua interação com o ambiente (MORAES NETO et al., 2000; SCALON et al., 2002). As informações sobre as respostas morfofisiológicas das plantas a condições distintas de luz podem ser críticas para determinar o seu potencial de crescimento e ocorrência e avaliar a sua capacidade competitiva em diferentes condições ambientais (DIAS-FILHO, 1997).
A composição do substrato é outro fator que desempenha papel importante na germinação e no sucesso do crescimento da planta. Segundo Carvalho Filho et al. (2002), o substrato exerce influência na arquitetura do sistema radicular e no estado nutricional das plantas. A depender da estrutura do substrato, da qualidade de aeração, da capacidade de retenção de água e do seu grau de infestação por patógenos, haverá ou não sucesso na germinação das sementes e no desenvolvimento das plantas (POPINIGIS, 1985).
A análise de crescimento, além de ser instrumento eficaz na avaliação do efeito de fatores como temperatura, água e substrato, é um importante indicador do grau de tolerância das espécies à luz e, ou, à sombra, sendo meio preciso para avaliar o crescimento e inferir a contribuição de diferentes processos fisiológicos sobre o comportamento vegetal (CASTRO et al., 1996; BENINCASA, 2003). Segundo Vieira et al. (1998), o conhecimento com relação ao crescimento e incorporação de biomassa das espécies é imprescindível quando se pretende utilizá-las para fins comerciais ou em programas de recuperação de áreas ambientalmente deterioradas. A análise de crescimento baseia-se no fato de que 90%, em média, da matéria seca acumulada pelas plantas ao longo do seu crescimento resulta da atividade fotossintética (BENINCASA, 2003).
Com relação às espécies do gênero Calliandra da região de Mucugê, contempladas neste estudo, existe apenas conhecimento quanto à sua taxonomia e distribuição (SOUZA, 2001), sendo inexistentes estudos fisiológicos. O objetivo deste trabalho foi estabelecer a melhor condição de propagação por meio de estudos sobre a influência da luz e do substrato na germinação e desenvolvimento inicial de C. viscidula e C. hygrophila.
2. MATERIAL E MÉTODOS
Os frutos de Calliandra viscidula Benth. e Calliandra hygrophila Mackinder e Lewis foram coletados no Parque Municipal de Mucugê, Bahia (13º S; 41º 22' W; 984 m Alt.), em março de 2004 e acondicionados em saco de papel. As sementes foram retiradas dos frutos e selecionadas, descartando-se as murchas e predadas.
Os experimentos foram desenvolvidos na Unidade Experimental Horto Florestal da Universidade Estadual de Feira de Santana (Feira de Santana, Bahia).
Para avaliar o efeito da luz na germinação de C. hygrophila e C. viscidula, as sementes foram desinfestadas em hipoclorito de sódio 0,5% por 10 min e lavadas em água destilada. A semeadura ocorreu em placas de Petri (100 X 20 mm) forradas com duas camadas de papel germitest esterilizados a 105 ºC por 4 h e umedecidas em 8 mL de água destilada. Em seguida, as sementes foram submetidas a três diferentes luminosidades: luz/escuro (12 h/12 h), escuro e luz e mantidas a 25 ºC em câmara de germinação do tipo BOD. No tratamento de escuro, as placas de Petri foram envolvidas com duas folhas de papel-alumínio.
A germinação das sementes nos diferentes tratamentos foi avaliada diariamente durante 21 dias após a semeadura. A semente foi considerada germinada quando houve a emissão de 2 a 5 mm da raiz. No tratamento de escuro, observou-se a germinação em sala escura sob luz verde de segurança.
Os parâmetros avaliados da germinação foram baseados em Maguire (1962), Borghetti (2000) e Santana e Ranal (2000) e consistiram de: Germinabilidade (%G), Índice de Velocidade de Germinação (IVG), Tempo Médio de Germinação (Tm) e Frequência Relativa da Germinação (Fr).
Para análise da interação de diferentes níveis de luminosidade e substrato na germinação e desenvolvimento inicial, a semeadura foi feita a 1 cm de profundidade, em embalagens de polietileno (10 X 18 X 5 cm) contendo três diferentes substratos: areia, terra vegetal + areia (1:1) e vermiculita de granulação média + terra vegetal + areia (2:1:1). O experimento foi mantido a 100% (pleno sol) e em viveiro a 70% de luminosidade, obtida mediante o uso de tela sombrite. A irrigação foi feita diariamente, no início da manhã e no final da tarde.
A avaliação da germinação sob condições de viveiro foi realizada considerando a emergência do cotilédone e utilizando a porcentagem de emergência (%E) e índice de velocidade de emergência (IVE) (MAGUIRE, 1962), além do tempo médio e da frequência relativa da emergência.
A resposta fisiológica das plântulas aos diferentes tratamentos foi avaliada aos 130 dias pós-semeadura, quando se avaliaram o comprimento da parte aérea (cm), a medida da superfície do solo até a inserção da última folha, o comprimento da raiz principal (cm) e o número de folhas (BENINCASA, 2003). Esses parâmetros foram avaliados com o auxílio de régua milimetrada e de paquímetro de precisão (150 mm). As plântulas foram separadas em folhas, caule e raízes, sendo a massa seca (g) determinada após a secagem em estufa com ventilação forçada a 60 ºC, por 72 h. Com o resultado da massa seca foram estimadas a Relação Parte Aérea/Sistema Radicular (PA/SR) e a Razão do Peso da Folha (RPF), calculadas a partir da divisão do peso de matéria seca da folha pelo peso de matéria seca total (UCHIDA e CAMPOS, 2000; BENINCASA, 2003).
Nos ensaios de germinação, utilizou-se o delineamento inteiramente casualizado, sendo cada parcela composta por 20 sementes, com cinco repetições. Os resultados expressos em porcentagem foram transformados em arcosseno (%G /100)0,5, para a normalização da sua distribuição.
Para os ensaios de germinação e desenvolvimento inicial em viveiro, considerou-se o delineamento inteiramente casualizado em arranjo fatorial 2 X 3 (dois níveis de luz e três substratos). Cada parcela foi composta por uma plântula e 10 repetições.
Os dados foram submetidos à análise de variância e as médias, comparadas pelo teste de Scott-Knott a 5% de probabilidade. Utilizou-se o software Sisvar 4.3 (FERREIRA, 2003).
Ao sexto mês da semeadura foi avaliada a porcentagem de sobrevivência das plantas de C. viscidula e C. hygrophila.
3. RESULTADOS
Após 21 dias da semeadura, as sementes de C. viscidula e C. hygrophila apresentaram valores para germinabilidade entre 80 e 84% para C. hygrophila, enquanto C. viscidula obteve entre 67 e 71% nos três diferentes tratamentos de luz (Tabela 1).
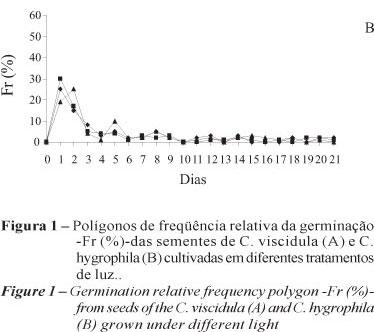
Os polígonos de frequência relativa da germinação das sementes de C. viscidula demonstraram maior pico no primeiro dia da semeadura, com valores de germinabilidade de 40 a 50% (Figura 1A). Em C. hygrophila, o pico de germinação estendeu-se até o segundo dia da semeadura, obtendo índice de 20-30% (Figura 1B). Esses resultados demonstraram a diferença na cinética da germinação entre as espécies estudadas e refletiram nos valores do IVG maiores em C. viscidula, em todos os tratamentos (Tabela 1).

A cinética da germinação das espécies nos três diferentes substratos e nas duas luminosidades foi semelhante às observadas em laboratório, onde inicialmente ocorreu pico de germinação nos primeiros dois dias, reduzindo no decorrer dos 21 dias de avaliação. Esse pico de germinação, diferente do ensaio em câmara de germinação, ocorreu no terceiro dia de semeadura, já que nesse caso foi considerada a emergência do cotilédone e não a protusão da raiz primária como semente germinada.
Em ambas as espécies, as médias da germinabilidade não diferiram nas luminosidades e substratos testados (p<0,05), alcançando valores entre 62 e 75% em C. viscidula e entre 71 e 89% em C. hygrophila (Tabela 2).
Os valores de IVE não diferiram estatisticamente nos dois níveis de luminosidade, nas duas espécies estudadas, com exceção do substrato areia + terra vegetal, em que se apresentaram inferiores aos demais a 70% de luminosidade em C. viscidula e a pleno sol em C. hygrophila.
Em C. viscidula, o maior valor de Tm ocorreu no substrato areia + terra vegetal a 70% de luminosidade (6,61). A diferença foi significativa entre as luminosidades, em C. hygrophila, no substrato areia + terra vegetal, com valor significativamente alto a 100% de luminosidade (10,06).
Em geral houve tendência a melhores valores para o comportamento germinativo no substrato areia em ambas as espécies, a 100% de luminosidade em C. viscidula e a 70% em C. hygrophila (Tabela 2).
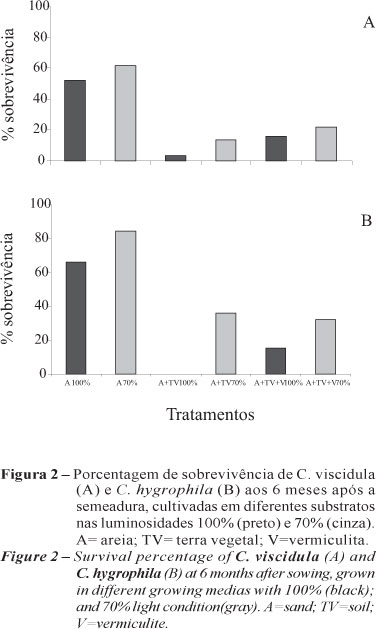
Na análise da resposta fisiológica aos diferentes tratamentos de luz e substrato, em C. viscidula o maior acúmulo da massa seca ocorreu na mistura areia + terra vegetal + vermiculita a 70% de luminosidade (0,191 g), enquanto em C. hygrophila os maiores valores foram encontrados nos substratos areia (0,121 g) e areia + terra vegetal (0,122 g) (Tabela 3). Apesar desses resultados, a porcentagem de sobrevivência foi melhor no substrato contendo somente areia nos dois níveis de luminosidade, em ambas as espécies. Os maiores valores foram obtidos nesse substrato a 70% de luminosidade em C. viscidula (61,82%) e C. hygrophila (84,06%) (Figura 2).
Com relação ao comprimento da parte aérea, as duas espécies de Calliandra alcançaram valores significativamente maiores no ambiente sombreado, com exceção do substrato areia + terra vegetal em C. viscidula e areia + terra vegetal + vermiculita em C. hygrophila (Tabela 3).
Em C. hygrophila, os maiores valores quanto a número de folhas ocorreu nos substratos areia (4,1) e areia + terra vegetal (4,7) a 70% de luminosidade, enquanto em C. viscidula não houve diferença entre os tratamentos (Tabela 3).
Os dados obtidos de comprimento da raiz principal em C. viscidula não diferiram nas luminosidades e substratos testados (Tabela 3). Em C. hygrophila houve diferença entre as luminosidades no substrato areia + terra vegetal, em que a pleno sol o maior comprimento da raiz ocorreu no substrato contendo somente areia (13,97cm). Esse resultado se repetiu na menor luminosidade, e, além do substrato areia (14,94 cm), o substrato areia + terra vegetal possibilitou maior crescimento (14,57 cm) (Tabela 3).
Na Relação Parte Aérea/Sistema Radicular, o maior valor foi obtido no substrato areia a 70% de luminosidade em C. viscidula (1,439). Em C. hygrophila, valor significativamente maior foi alcançado no substrato areia + terra vegetal a 70% de luminosidade (1,956) (Tabela 3).
Os mesmos resultados da Relação Parte Aérea/Sistema Radicular ocorreram na Relação Peso da Folha. Essa relação expressa a fração de massa seca não exportada das folhas no resto da planta (BENINCASA, 2003), indicando que a 70% de luminosidade houve maior desenvolvimento da parte aérea com exceção de C. viscidula no substrato areia + terra vegetal, em que não houve diferença significativa entre as luminosidades.
Apesar da variação ocorrida nos diferentes parâmetros, em geral aos 130 dias de idade houve tendência a melhores resultados para o desenvolvimento inicial no ambiente a 70% de luminosidade na mistura areia + terra vegetal + vermiculita em C. viscidula e em C. hygrophila nos substratos areia e areia + terra vegetal (Tabela 3).
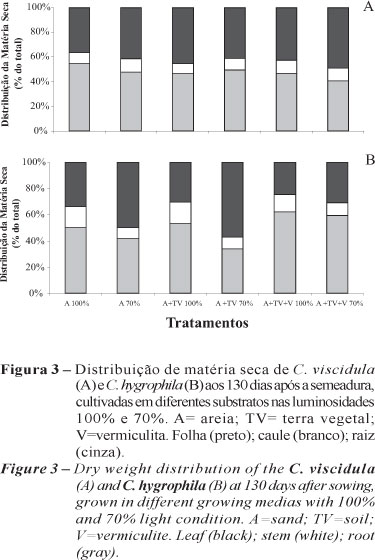
A distribuição de massa seca em porcentagem do total nos diferentes órgãos (folha, caule e raiz) mostrou que em C. viscidula, com exceção do substrato areia a 100% de luminosidade, a parte aérea contribuiu mais do que as raízes na composição da planta (Figura 3). Em C. hygrophila, o sistema radicular teve maior papel na contribuição da massa seca total a pleno sol em todos os substratos testados e a 70% de luminosidade somente no substrato areia + terra vegetal + vermiculita (Figura 3).
4. DISCUSSÃO
Os resultados da avaliação do efeito da luz na germinação indicam que, diferentemente de muitas leguminosas, C. viscidula e C. hygrophila não apresentaram dormência. As espécies tiveram valores de germinabilidade considerados altos, em comparação com os de outras espécies de leguminosas, como Caesalpinia peltophoroides Benth. (43,2%) (SCALON et al., 2002).
As diferenças na germinabilidade entre as espécies, além de serem inerentes às características intrínsecas, podem estar relacionadas com o processo de deterioração das sementes iniciado aos oito dias após a semeadura de C. viscidula, o que não foi observado em C. hygrophila, que manteve sementes germinando até o final do ensaio. Essa maior viabilidade das sementes de C. hygrophila pode ser atribuída à sua melhor adequação a ambientes úmidos, por serem espécies de ocorrência em margens de rios.
Ao contrário do que afirmaram Maluf e Wizentier (1998), as germinações baixas e tardias influenciaram no valor do IVG em C. hygrophila, resultando em valores do Tm significativamente mais altos, em comparação com C. viscidula. Segundo Borghetti e Ferreira (2004), a partir dos dados de tempo médio de germinação é possível inferir que a rápida germinação das sementes é característica de espécies cuja estratégia é estabelecer no ambiente rapidamente, quando este lhe oferece condições propícias para o desenvolvimento.
A análise estatística dos parâmetros de germinação demonstrou que ambas as espécies se comportaram como sementes fotoblásticas neutras, não diferindo nos diferentes tratamentos de luz. Segundo Aguiar et al. (2005), essa plasticidade das sementes é de grande importância ecológica, por não requerer condições específicas de luminosidade.
Resultados cujas médias de germinabilidade não diferiram nos diferentes substratos e luminosidades, como em C. viscidula e C. hygrophila, foram encontrados em sementes de Cassia grandis L. e Caesalpinia echinata Lam., nas quais a germinação não apresentou diferença significativa nos diferentes níveis de luminosidade, que variaram entre 30 e 100% e 20 e 100%, respectivamente, demonstrando que são espécies indiferentes à luz (CARVALHO FILHO et al., 2002; AGUIAR et al., 2005). Entretanto, Fonseca et al. (1994), estudando o efeito de diferentes profundidades e de dois níveis de luminosidades (100% e 50%) na germinação de Dipteryx alata Vog., obtiveram os melhores índices de germinabilidade a 100% de luminosidade.
Em geral, os valores de IVE em C. viscidula e C. hygrophila não diferiram estatisticamente nos dois níveis de luminosidade e foram considerados altos em comparação com os de outras espécies, como Caesalpinia peltophoroides Benth, que alcançou valores de IVE de 1,28 (SCALON et al., 2002). A maior umidade dos substratos no ambiente a 70% de luminosidade, observado no decorrer do experimento, pode ter sido fator determinante para a germinação mais rápida em C. hygrophila, que é encontrada em margens de rios. Segundo Fonseca et al. (1994), na condição de sombreamento parcial o processo de dessecação das sementes é mais lento e moderado do que nas condições a pleno sol. Com relação às sementes de C. viscidula, a tendência a maiores valores de IVE a pleno sol, sendo significativo no substrato areia + terra vegetal, pode indicar que é espécie sensível à intensidade luminosa. O comportamento de melhor germinação em ambientes com alta intensidade de luz demonstra a adequação da espécie em fitofisionomias mais abertas, o que é característico dos locais de ocorrência de C. viscidula (FONSECA et al., 1994).
Com relação ao substrato, a areia não possibilita uniformidade na capacidade de retenção de água devido à evaporação e ao rápido escoamento. Isso pode acarretar maior dificuldade no processo de embebição contínua da semente, o que pode retardar o início do processo de germinação. Apesar dessas características, o maior IVE nesse substrato para ambas as espécies pode ser atribuído à sua composição estrutural, que facilitou a emergência dos cotilédones. (FONSECA et al., 1994; ANDRADE et al., 2000).
O melhor desenvolvimento inicial no ambiente a 70% de luminosidade das espécies estudadas aos 130 dias de idade pode estar relacionado a uma adaptação dessas espécies a baixas intensidades luminosas, que é característica genética que resulta em modificações anatômicas das folhas e propriedades fisiológicas (SCALON et al., 2002). A capacidade de crescer no ambiente a 70% de luminosidade é mecanismo importante de adaptação da espécie, o que constitui uma valiosa estratégia para escapar às condições de baixa intensidade luminosa (MORAES NETO et al., 2000; SCALON et al., 2002). Com relação ao substrato, apesar da variação ocorrida nas diferentes variáveis, os resultados indicaram que a presença da matéria orgânica no substrato pode ter favorecido melhor desenvolvimento inicial das plantas.
As espécies estudadas apresentaram tendência à maior acúmulo de massa seca total a 70% de luminosidade, independentemente do substrato, demonstrando que, quanto menor a luminosidade, maior a eficiência na produção de biomassa.
Estudos indicam que a maior produção de massa seca em ambientes sombreados foi encontrada em Coffea arabica L., Genipa americana L., Muntingia calabura L., Ceiba pentandra (L.) Gaertn e Dinizia excelsa Ducke (VARELA e SANTOS, 1992; PEDROSO e VARELA, 1995; CASTRO et al., 1996; MORAES NETO et al., 2000; PAIVA et al., 2003). Entretanto, em mudas de Dipteryx odorata (Aubl.) Willd. foi verificada tendência à redução da massa seca total com o aumento do sombreamento (UCHIDA e CAMPOS, 2000). Espécies tolerantes ao sombreamento possuem baixo ponto de compensação luminosa, sendo capazes de produzir fotoassimilados em baixas intensidades luminosas, o que sugere que essas espécies possuam mecanismos morfológicos que possibilitam melhor uso da radiação fotossinteticamente ativa (MORAES NETO et al., 2000; ALVARENGA et al., 2003). Além disso, o melhor desempenho no ambiente sombreado pode ter sido favorecido pela menor temperatura, que reduz a evaporação e, consequentemente, aumenta a disponibilidade de água, o que pode ter possibilitado melhor desenvolvimento das plantas. Segundo Fonseca et al. (2002), a diminuição da radiação solar com telas sombrite pode amenizar a temperatura dentro do viveiro em até cinco graus Celsius.
O sombreamento pode ter favorecido a sobrevivência nesse ambiente, assim como em Euterpe edulis Mart., em que o melhor percentual de sobrevivência ocorreu a 50% de luminosidade nos três diferentes níveis de sombreamento (0%, 18%, 50%) estudados (NODARI et al., 1999).
Os resultados da distribuição da massa seca em porcentagem do total nos diferentes órgãos (folha, caule e raiz) demonstraram maior alocação de biomassa no sistema radicular a 100% e na parte aérea a 70%, o que concorda com a afirmação de que as plantas sombreadas tendem a concentrar menos massa seca na raiz que plantas em ambientes não sombreados (ALVARENGA et al., 2003). Dados semelhantes foram encontrados por Castro et al. (1996), que encontraram maior direcionamento da massa seca nas raízes com o aumento da irradiância, ao investigarem a influência de diferentes níveis de luminosidade (100%, 70% e 50%) no crescimento de Muntingia calabura L.
A menor distribuição da massa seca do sistema radicular no ambiente sombreado pode estar relacionada à redução na translocação dos assimilados dos órgãos fotossintéticos para esse órgão, devido à menor intensidade luminosa nesse ambiente, já que a luz exerce influência positiva nesse processo (VARELA e SANTOS, 1992; SOUZA et al., 1999).
Com relação ao comprimento da parte aérea, resultados semelhantes a de C. viscidula e C. hygrophila foram encontrados em Clitoria fairchildiana Howard, Cassia grandis L. e Muntingia calabura L., em que o sombreamento favoreceu o crescimento no comprimento da parte aérea das plantas (CASTRO et al., 1996; PORTELA et al., 2001; CARVALHO FILHO et al., 2002).
Scalon et al. (2002), investigando a influência de diferentes níveis de sombreamentos (0%, 30% e 50%), encontraram maiores valores de comprimento da parte aérea a pleno sol em Caesalpinia peltophoroides Benth. Os diferentes níveis de luminosidade não afetaram o comprimento da parte aérea de Caesalpinia echinata Lam., Ceiba pentandra (L.) Gaertn e Dipteryx odorata (Aubl.) Willd. (UCHIDA e CAMPOS, 2000; PAIVA et al., 2003; AGUIAR et al., 2005).
Devido à estreita relação entre comprimento da parte aérea e massa seca e o número de folhas (CARVALHO FILHO et al., 2002), é possível concluir que o maior crescimento em comprimento da parte aérea das espécies de Calliandra sob as condições de sombreamento foi em função do desenvolvimento e não do fenômeno de estiolamento.
Assim como em C. viscidula e C. hygrophila, foi encontrado em Cassia grandis L., em estudos com diferentes substratos, tamanho de recipientes e diferentes luminosidades (100% e 50%), maior número de folhas no ambiente sombreado, o mesmo ocorrendo em Coffea arabica L. (CARVALHO FILHO et al., 2002; PAIVA et al., 2003).
Uchida e Campos (2000) não verificaram diferença no número de folhas em Dipteryx odorata (Aubl.) Willd., ao investigarem a influência de diferentes sombreamentos (0%, 30%, 50% e 70%), assim como Nodari et al. (1999) estudando plantas de Euterpe edulis Mart. nos níveis de sombreamento de 0%, 18% e 50%; e Souza et al. (1999), investigando a influência de três níveis de irradiância (100%, 70% e 50%) em Raphanus sativus L.
Resultados semelhantes de comprimento da raiz principal em C. viscidula foram encontrados em Amburana cearensis (Arr. Cam.) A.C. Smith e Clitoria fairchildiana Howard (UCHIDA e CAMPOS, 2000; PORTELA et al., 2001). Em C. hygrophila, apesar de ocorrer em beira de rio e ter pouco desenvolvimento radicular, os resultados indicaram que essa espécie apresenta maior plasticidade morfológica ao obter bons resultados no substrato areia, que se caracteriza pela não retenção da água. O maior desenvolvimento no sistema radicular nesse substrato pode evidenciar que essa espécie possua mecanismos de resistência à seca (VIDAL et al., 1999).
A diferença significativa na Relação Parte Aérea/Sistema Radicular indicou que as mudas apresentaram padrão de distribuição da massa seca entre os dois órgãos, dependente do sombreamento durante o início do desenvolvimento. Resultados semelhantes com relação ao nível de luz foram encontrados em Dinizia excelsa Ducke (VARELA e SANTOS, 1992). Em contraposição, maiores valores da Relação Parte Aérea/Sistema Radicular foram encontrados a 100% de luminosidade em Muntingia calabura L., quando se estudaram diferentes níveis de sombreamento (0%,50%, 70%) no desenvolvimento dessa espécie (CASTRO et al., 1996).
5. CONCLUSÃO
Calliandra viscidula e C. hygrophila são fotoblásticas neutras e apresentaram melhor crescimento aos 130 dias, a 70% de luminosidade, independente do substrato.
6. AGRADECIMENTOS
Aos funcionários do Projeto Sempre - Viva (Mucugê - Bahia), pelo auxílio nas coletas das sementes; e à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pela concessão da bolsa.
7. REFERÊNCIAS
Recebido em 02.07.2008 e aceito para publicação em 25.08.2010.
- AGUIAR, F. F. A. et al. Germinação de sementes e formação de mudas de Caesalpinia echinata Lam. (pau-brasil): efeito de sombreamento. Revista Árvore, v.29, n.6, p.871-875, 2005.
- ALVARENGA, A. A. et al. Effects of different light levels on the initial growth and photosynthesis of Croton urucurana Baill in southeastern Brazil. Revista Árvore, v.27, n.1, p.53-57, 2003.
- ANDRADE, A. C. S. et al. Germinação de sementes de jenipapo: temperatura, substrato e morfologia do desenvolvimento pós-seminal. Pesquisa Agropecuária Brasileira, v.35, n.3, p.609-615, 2000.
- BARNEBY, R. C. Silk tree, guanacaste, monkey's earring: A generic system for the synandrous Mimosaceae of the Americas. Memoire New York Botanic Gardner, v.74, n.3, p.1-223, 1998.
- BENINCASA, M. M. P. Análise de crescimento de plantas (noções básicas). Jaboticabal: FUNEP, 2003. 41p.
- BORGHETTI, F. Ecofisiologia da germinação de sementes. Universa, v.8, n.1, p.149-180, 2000.
- BORGHETTI, F.; FERREIRA, A. G. Interpretações de resultados de germinação. In: FERREIRA, A. G.; BORGHETTI, F. Germinação: do básico ao aplicado. Porto Alegre: Artmed., 2004. 323p.
- CARVALHO FILHO, J. L. S. et al. Produção de mudas de Cassia grandis L. em diferentes ambientes, recipientes e misturas de substratos. Revista Ceres, v.40, n.284, p.341-352, 2002.
- CASTRO, E. M.; ALVARENGA, A. A.; GOMIDE, M. B. Crescimento e distribuição de matéria seca de mudas de calabura (Muntingia calabura L.) submetidas a três níveis de irradiância Ciência e Agrotecnologia, v.20, n.3, p.357-365, 1996.
- CHAMBERLAIN, J. R. Improving Seed Production in Calliandra calothyrsus: A Field Manual for Researchers and Extension Workers Oxford: Oxford Forestry Institute, 2000. 37p. (Miscellaneous Paper).
- DIAS-FILHO, M. B. Physiological responses of Solanum crinitum LAM. to contrasting light environments. Pesquisa Agropecuária Brasileira, v.32, n.8, p.789-796, 1997.
- FERREIRA, D. F. SISVAR Sistema de análises estatísticas, v 4.3. Lavras: Universidade Federal de Lavras, 2003.
- FONSECA, C. E. L.; FIGUEIREDO, S. A.; DA SILVA, J. A. Influência da profundidade de semeadura e da luminosidade na germinação de sementes de Baru (Dipteryx alata Vog.) Pesquisa Agropecuária Brasileira, v.29, n.4, p.653-659, 1994.
- FONSECA, E. P. et al. Padrão de qualidade de mudas de Trema micrantha (L) Blume, produzidas sob diferentes períodos de sombreamento. Revista Árvore, v.26, n.4, p.515-523, 2002.
- FONSECA, M. G.; LEÃO, N. V. M.; SANTOS, F. A. M. Germinação e crescimento inicial de plântulas de Pseudopiptadenia psilostachya (DC.) G.P. Lewis & M.P. Lima (Leguminosae) em diferentes ambientes de luz. Revista Árvore, v.30, n.6, p.885-891, 2006.
- LIMA, J. D. et al. Efeito da temperatura e do substrato na germinação de sementes de Caesalpinia férrea Mart. ex. Tul. (Leguminosae- Caesalpinoideae). Revista Árvore, v.30, n.4, p.513-518, 2006.
- LORENZI, H. C.; SOUZA, H. M. Plantas Ornamentais no Brasil: arbustos, herbáceas, e trepadeiras. 3.ed. Nova Odessa: Instituto Plantarum, 2001. 1088p.
- MAGUIRE, J. D. Speed of germination-aid in selection and evolution for seedling emergence vigor. Crop Science, v.2, p.176-177, 1962.
- MALUF, A. M.; WIZENTIER, B. Aspectos fenológicos e germinação de sementes de quatro populações de Eupatorium vauthierianum DC. (Asteraceae) Revista Brasileira de Botânica, v.21, n.3, p.247-251, 1998.
- MORAES NETO, S. P. et al. Crescimento de mudas de algumas espécies arbóreas que ocorrem na mata atlântica, em função do nível de luminosidade. Revista Árvore, v.24, n.1, p.35-45, 2000.
- NODARI, R. B. et al. Crescimento de mudas de palmiteiro (Euterpe edulis Mart.) em diferentes condições de sombreamento e densidade. Revista Árvore, v.23, n.3, p.285-292, 1999.
- PAIVA, L. C.; GUIMARÃES. R. J.; SOUZA, C. A. S. Influência de diferentes níveis de sombreamento sobre o crescimento de mudas de cafeeiro (Coffea arabica). Ciência e Agrotecnologia, v.27, n.1, p.134-140, 2003.
- PEDROSO, S. G.; VARELA, V. P. Efeito do sombreamento no crescimento de mudas de sumaúma (Ceiba pentandra (L.) Gaertin). Revista Brasileira de Sementes, v.17, n.1, p.47-51, 1995.
- POPINIGIS, F. Fisiologia da semente 2.ed. Brasília: AGIPLAN, 1985. 289p.
- PORTELA, R. C. Q.; SILVA, I. L.; PIÑA-RODRIGUES, F. C. M. Crescimento inicial de mudas de Clitoria fairchildiana Howard e Peltophorum dubium (Sprenge) Taub. em diferentes condições de sombreamento. Revista Ciência Florestal, v.11, n.1, p.163-170, 2001.
- SANTANA, D. G.; RANAL, M. A. Análise estatística na germinação. Revista Brasileira de Fisiologia Vegetal, v.12, Edição Especial, p.205-237, 2000.
- SCALON, S. P. Q. et al. Crescimento inicial de mudas de espécies florestais nativas sob diferentes níveis de sombreamento. Revista Árvore, v.26, n.1, p.1-5, 2002.
- SILVA, A. et al. Interação de luz e temperatura na germinação de sementes de Esenbeckia leiocarpa Engl. (Guarantá). Revista do Instituto Florestal, v.9, n.1, p.57-64, 1997.
- SMIDERLE, O. J.; SOUSA, R. C. P. Dormência em sementes de paricarana (Bowdichia virgilioides Kunt- Fabaceae - Papilionidae. Revista Brasileira de Sementes, v.25, n.2, p.48-52, 2003.
- SOUZA, E.R. Aspectos taxonômicos e biogeográficos do gênero Calliandra Benth. (Leguminosae - Mimosoideae) na Chapada Diamantina, Bahia, Brasil 2001. 187f. Dissertação (Mestrado em Botânica) - Universidade Estadual de Feira de Santana, Feira de Santana, 2001.
- SOUZA, J. R. P. et al. Sombreamento e o desenvolvimento e produção de rabanete. Scientia Agrícola, v.56, n.4, p.987-992, 1999.
- UCHIDA, T.; CAMPOS, M. A. A. Influência do sombreamento no crescimento de mudas de cumaru (Dipteryx odorata (AUBL.) WILLD. -Fabaceae), cultivdas em viveiro. Acta Amazônica, v.30, n.1, p.107-114, 2000.
- VARELA, V. P.; SANTOS, J. Influência do sombreamento na produção de mudas de angelim pedra (Dinizia excelsa Ducke) Acta Amazônica, v.22, n.3, p.407-411, 1992.
- VIDAL, M. C.; SERAPHIN- STACCIARINI, E.; CÂMARA, H. H. L. L. Crescimento de plântulas de Solanum lycocarpum St. Hill. (Lobeira) em casa de vegetação. Acta Botanica Brasílica, v.13, n.3, p.271-274, 1999.
- VIEIRA, F. et al. Schizolobium parahybum (Vell.) Blake - Uma análise de desenvolvimento em três ambientes, na fase de viveiro. Floresta e Ambiente, v.5, n.1, p.118-123, 1998.
Datas de Publicação
-
Publicação nesta coleção
11 Abr 2011 -
Data do Fascículo
Fev 2011
Histórico
-
Aceito
25 Ago 2010 -
Recebido
02 Jul 2008