Resumos
Objetivou-se analisar o comportamento do epitélio branquial de tilápias, cultivadas em tanques posicionados em diferentes altitudes e interconectados por tubos de PVC. Filamentos branquiais de quatro espécimes de quatro tanques (T1, T2, T3 e T4) interconectados seqüencialmente foram submetidos à rotina histológica, para obtenção de cortes de 5µm de espessura, os quais foram corados com Hematoxilina-Eosina, ou submetidos à técnica histoquímica para glicoconjugados: PAS + solução de diástase ou Alcian Blue pH 2,5 ou Alcian Blue pH 1,0. Considerando as regiões basal, intermediária e apical dos filamentos, mensurou-se a área lamelar e contou-se o número de células mucosas em cada uma dessas regiões, o que correspondia a 0,56 mm². Verificou-se que a concentração de oxigênio, pH e a temperatura se reduziam progressivamente com a passagem da água de um tanque para outro. Em função disso, constatou-se um aumento abrupto do número de células mucosas e da área lamelar no T2, e uma redução progressiva destas medidas nos tanques que recebiam água do T2. Além disso, observou-se nos animais do T2, T3 e T4, descolamento do epitélio branquial, hiperplasia celular no espaço interlamelar e telangectasias. Conclui-se que o ambiente aquático de tanques interconectados seqüencialmente por tubos de PVC se altera ao passar de um tanque para o outro, e que estas flutuações físico-químicas se refletem no comportamento do epitélio branquial através de variações da área lamelar e do número de células mucosas.
Tilápias nilóticas; cultivo intensivo; brânquia; morfometria; células mucosas e lamelas
The behavior of the gill epithelium of tilapias cultured in tanks at different altitudes and interconnected with PVC pipes was analyzed. Gill filaments of four specimens from four tanks (T1, T2, T3 e T4) sequentially interconnected were submitted to histological routine to obtain 5-mm-thick cuts that were stained with HE or submitted to histochemistry reactions PAS + diastase solution or Alcian Blue pH 2.5 or Alcian Blue pH 1.0. Considering the intermediary, apical and basal regions of the filaments, the lamellar area was measured and the amount of mucous cells was counted. It was verified that oxygen, pH, and temperature decreased progressively as the water flew from one tank to another. Thus, an increase was realized of the amount of mucous cells and the lamellar area in T2, as well as a progressive decrease of these measures on the tanks which received water from T2. Moreover, detachment of the gill epithelium, cellular hyperplasia in the interlamellar space, and telangectasias were observed in the fishes from T2, T3 e T4. It was concluded that the aquatic environment in tanks sequentially interconnected with PVC pipes suffers alterations from one tank to another, as that physico-chemical fluctuations reflect on the behavior of the gill epithelium through variations of the lamellar area and the amount of mucous cells.
Nile tilapia; intensive culture system; gill; morphometry; mucous cells; secundary lamellar cells
Alterações do epitélio branquial e das lamelas de tilápias (Oreochromis niloticus) causadas por mudanças do ambiente aquático em tanques de cultivo intensivo1 1 Esta pesquisa recebeu apoio financeiro da Universidade Paranaense (Unipar) e refere-se ao Trabalho de Conclusão de Curso de Graduação em Enfermagem pela Universidade do primeiro autor.
The influence of the aquatic environment in tanks sequetially interconnected with PVC pipes on the gill epithelium and lamellas of tilapia (Oreochromis niloticus)
Anderson Brunetti ReisI; Débora de Mello Gonçales Sant'AnaII; Jorge Fernandes de AzevedoIII; Luiz Sérgio MerliniII; Eduardo José de Almeida AraújoII, * * Autor para correspondência: eduardoaraujo@unipar.br
IEnfermeiro pela Unipar, Rua Paraíba 228, Centro, Cruzeiro do Oeste, PR 87400-000 Brasil
IILaboratório de Neurogastroenterologia Experimental, Unipar, Praça Mascarenhas de Moraes 4286, Umuarama, PR 87502-210, Brasil
IIIAcadêmico do Mestrando em Ciência Animal da Unipar, Umuarama, PR, Brasil
RESUMO
Objetivou-se analisar o comportamento do epitélio branquial de tilápias, cultivadas em tanques posicionados em diferentes altitudes e interconectados por tubos de PVC. Filamentos branquiais de quatro espécimes de quatro tanques (T1, T2, T3 e T4) interconectados seqüencialmente foram submetidos à rotina histológica, para obtenção de cortes de 5µm de espessura, os quais foram corados com Hematoxilina-Eosina, ou submetidos à técnica histoquímica para glicoconjugados: PAS + solução de diástase ou Alcian Blue pH 2,5 ou Alcian Blue pH 1,0. Considerando as regiões basal, intermediária e apical dos filamentos, mensurou-se a área lamelar e contou-se o número de células mucosas em cada uma dessas regiões, o que correspondia a 0,56 mm2. Verificou-se que a concentração de oxigênio, pH e a temperatura se reduziam progressivamente com a passagem da água de um tanque para outro. Em função disso, constatou-se um aumento abrupto do número de células mucosas e da área lamelar no T2, e uma redução progressiva destas medidas nos tanques que recebiam água do T2. Além disso, observou-se nos animais do T2, T3 e T4, descolamento do epitélio branquial, hiperplasia celular no espaço interlamelar e telangectasias. Conclui-se que o ambiente aquático de tanques interconectados seqüencialmente por tubos de PVC se altera ao passar de um tanque para o outro, e que estas flutuações físico-químicas se refletem no comportamento do epitélio branquial através de variações da área lamelar e do número de células mucosas.
Termos de indexação: Tilápias nilóticas, cultivo intensivo, brânquia, morfometria, células mucosas e lamelas.
ABSTRACT
The behavior of the gill epithelium of tilapias cultured in tanks at different altitudes and interconnected with PVC pipes was analyzed. Gill filaments of four specimens from four tanks (T1, T2, T3 e T4) sequentially interconnected were submitted to histological routine to obtain 5-mm-thick cuts that were stained with HE or submitted to histochemistry reactions PAS + diastase solution or Alcian Blue pH 2.5 or Alcian Blue pH 1.0. Considering the intermediary, apical and basal regions of the filaments, the lamellar area was measured and the amount of mucous cells was counted. It was verified that oxygen, pH, and temperature decreased progressively as the water flew from one tank to another. Thus, an increase was realized of the amount of mucous cells and the lamellar area in T2, as well as a progressive decrease of these measures on the tanks which received water from T2. Moreover, detachment of the gill epithelium, cellular hyperplasia in the interlamellar space, and telangectasias were observed in the fishes from T2, T3 e T4. It was concluded that the aquatic environment in tanks sequentially interconnected with PVC pipes suffers alterations from one tank to another, as that physico-chemical fluctuations reflect on the behavior of the gill epithelium through variations of the lamellar area and the amount of mucous cells.
Index terms: Nile tilapia, intensive culture system, gill; morphometry, mucous cells, secundary lamellar cells.
INTRODUÇÃO
Os recursos hídricos sempre determinaram a existência da vida, estando a saúde definitivamente relacionada com a existência desses recursos, e sua relação positiva com o meio ambiente (Maciel-Filho et al. 2003). A baixa qualidade da água pode estar associada ao aumento de doenças infecto-contagiosas, pois esta constitui veículo direto ou indireto de doenças entre os seres vivos, quando contaminada por agentes químicos e/ou biológicos como bactérias, fungos, vírus e parasitas. Alguns desses microorganismos podem afetar os peixes que estejam expostos a condições de estresse, tais adversidades podem provocar o surgimento de patologias. Essas doenças geram um ciclo de causa-efeito de difícil solução (Moraes & Jordão 2002).
As alterações do meio aquático também interferem significativamente na saúde humana, e contribuem para a elevação dos custos empregados no tratamento de doenças advindas desse meio, por isso, a busca por soluções para os problemas ambientais tornou-se prioridade no mundo. Para a Organização Mundial da Saúde, o saneamento do meio "é a ciência e a arte de promover, proteger e recuperar a saúde, através de medidas de alcance coletivo e de motivação da população". Desta maneira, o controle de todos os fatores do meio ambiente, que interagem com os seres humanos e que exercem ou podem exercer efeito nocivo sobre o bem-estar físico, mental e social do homem, deverão ser observados como a primeira ação da saúde pública (Carvalho & Oliveira 1997).
Considerando o campo da saúde pública, podemos encontrar água que não apresente nenhuma das mudanças ecológicas características da poluição, mas a mesma pode estar contaminada de organismos patogênicos, que acometem o homem e animais domésticos por intermédio de seu consumo ou pela ingestão de alimentos provenientes desse meio (Mellanby 1982, Moraes & Jordão 2002).
Em nutrição humana, o peixe constitui uma excelente fonte de proteínas de alto valor biológico, ainda apresenta quantidades significativas de fósforo, ferro, e outros sais minerais, bem como diversas vitaminas, em especial as do complexo B e vitaminas A e D. Em muitos países, principalmente da Europa e da Ásia, é a proteína de origem animal mais consumida (Hermanns & Bellé 2006). Além disso, a carne do pescado contém ácidos graxos altamente insaturados, os quais detêm benefícios nutricionais e medicinais como prevenção e tratamento de doenças cardiovasculares, hipertensão, inflamações em geral e outros tipos de doença (Suarez-Mahecha et al. 2002).
Os órgãos como a pele, tubo digestório e as brânquias estão em contato direto com a água e com as alterações físico-químicas ambientais. Devido a isso, eles podem sofrer alterações morfológicas, o que poderá se refletir como perturbações reversíveis ou irreversíveis à fisiologia destes órgãos. Vários são os estudos que utilizam as brânquias de peixes como bioindicadores de mudanças do meio aquático, visto serem estruturas vitais para a saúde desses animais, pois além de serem o principal local de trocas gasosas, também estão envolvidas nos processos de osmoregulação, equilíbrio ácido-básico, excreção de compostos nitrogenados e desempenham ainda a função de órgão sensorial da gustação (Araújo et al. 1997, 1999, 2000, 2001, Leonardo 1999, Sabóia-Morais et al. 1999, Winkaler et al. 2001, Lopes 2006, Figueiredo-Fernandes et al. 2007, Garcia-Santos et al. 2007, Fontaínhas-Fernandes et al. 2008). Qualquer comprometimento nos filamentos e lamelas branquiais, que interfiram em sua função certamente comprometerá a sobrevivência desses animais.
Certas condições podem comprometer a integridade estrutural das brânquias, levando a alterações histológicas, como edema e hiperplasia epitelial das lamelas secundárias, infiltração de células epiteliais, fusão lamelar, assim como a morte de células mucosas, devido a longos períodos de hipersecreção de muco, alterações estas, conseqüentes a uma resposta defensiva crônica a infecções parasitárias, bacterianas ou irritantes químicos (Leonardo 1999, Cavichiolo 2000, Figueiredo-Fernandes et al. 2007, Garcia-Santos et al. 2007, Fontaínhas-Fernandes et al. 2008).
Considerando o exposto, realizou-se este estudo no intuito de avaliar densidade populacional de células mucosas do epitélio branquial, assim como a área lamelar de brânquias de tilápias nilóticas cultivadas em tanques posicionados em diferentes altitudes e interconectados seqüencialmente através de tubos de PVC, visando à movimentação do ambiente aquático.
MATERIAL E MÉTODOS
O protocolo deste estudo foi previamente aprovado pelo Comitê de Ética em Pesquisa Envolvendo Experimentação Animal (CEPEEA) da Universidade Paranaense (UNIPAR) (protocolo 30/04, aprovado em 13/09/2004).
O sistema de piscicultura do fornecedor de animais. Este estudo foi realizado com tilápias nilóticas (Oreochromis niloticus) adultas cultivadas num Pesque-Pague da região Noroeste do Paraná, que utiliza o sistema de piscicultura intensivo, por meio de tanques conectados entre si através de tubulações de PVC. Dos vários tanques existentes no estabelecimento comercial, foram escolhidos quatro que estavam interconectados seqüencialmente para trânsito da água, devido a estarem posicionados em diferentes altitudes, sendo que o mais alto era o que recebia água direto de uma nascente. Desta forma, a água do tanque 1 (T1) passava para o tanque 2 (T2), que em seguida atravessava para o tanque 3 (T3), e por último para o tanque 4 (T4).
O primeiro tanque (o mais alto), considerado como tanque controle, tinha 2.700 m² (com aproximadamente 14.850 tilápias). Este recebia água de fonte natural, a qual se movia para os demais tanques por energia mecânica (sobretudo pelo seu componente gravitacional). O segundo tanque tinha 900 m² (~4.000 tilápias) e o terceiro 3.000 m² (~15.000 tilápias). O quarto tanque, de nível de altitude mais baixa, tinha 9.000m² (~40.000 tilápias).
Os animais deste pesque-pague foram adquiridos de fornecedores externos, quando tinham de 200-300g e foram alimentados com ração comercial SUPLA®.
Minutos antes da coleta dos animais, mensurou-se a temperatura, o pH e a concentração de oxigênio dissolvido na água, no intuito de tentar identificar se esses fatores poderiam estar relacionados à morfologia do epitélio branquial.
Obtenção dos animais e coleta das holobrânquias. Utilizando-se redes de pesca, quatro tilápias jovens de cada tanque com peso médio de 455,9g foram capturadas, as quais foram anestesiadas com 3g de benzocaína diluída em 20mL de álcool 99ºGL, para 30 litros de água. Os animais ficavam completamente sedados em 3-5 minutos, porém foram deixados neste meio por 10 minutos para garantir que fossem submetidos à eutanásia por aprofundamento anestésico. Em seguida, foi realizada pesagem dos animais, medidos o comprimento total (da cabeça até a extremidade da cauda), comprimento padrão (da cabeça até a inserção da cauda), altura (distância entre o ventre e dorso) e, cirurgicamente foi removida a segunda holobrânquia esquerda.
Análise histológica. Após 24 horas de fixação, por imersão em paraformaldeído a 4% em tampão fosfato de sódio 0,01M pH 7,4, os filamentos branquiais foram lavados com o mesmo tampão do fixador, desidratados em concentrações crescentes de etanol, diafanizados em xilol e incluídos em parafina. De cada filamento foram feitos oito cortes longitudinais de 5µm de espessura, sendo que dois foram corados com Hematoxilina-Eosina (HE), dois com Periodic Acid-Schff (PAS) + solução de diástase (para detecção de glicoconjugados neutros), dois com Alcian Blue pH 2,5 (para detecção de glicoconjugados ácidos: sialomucinas e sulfomucinas), e os dois remanescentes com Alcian Blue pH 1,0 (para detecção de glicoconjugados ácidos sulfatados: sulfomucinas).
Análise morfométrica das lamelas branquiais e densidade populacional de células mucosas. Nas lâminas histológicas, os filamentos branquiais foram medidos, utilizando régua milimetrada e um estereomicroscópio, e divididos em três partes iguais, as quais foram denominadas como Basal [B] (Região proximal ao rastelo), Intermediária [I] e Apical [A] (Região mais distal ao rastelo).
A área de 30 lamelas branquiais (de cada região do filamento: B, I e A) dos animais de cada tanque foi medida utilizando os cortes corados com HE. As imagens lamelares foram capturadas com auxílio de uma câmera digital Moticam 2000, 2.0m pixel USB 2.0 acoplada a um microscópio fotônico Motic B5, e as análises foram feitas utilizando-se o sistema computacional de análise de imagens Motic Images Plus 2.0.
Para avaliar a densidade populacional de células mucosas, contou-se o número total dessas células numa área total de 0,56mm² (o que correspondia a praticamente toda a extensão do filamento segmentado) por região dos filamentos branquiais coletados (B, I e A), utilizando uma ampliação de 400x através de um microscópio fotônico Motic BL. Também se avaliou a proporção do número de células mucosas produtoras de mucinas ácidas (AB pH 2,5) em relação ao número de células mucosas produtoras de mucinas neutras (P.A.S. + solução de diástase) tanto para o filamento branquial inteiro como para suas regiões acima denominadas.
Análise estatística. Todos os dados foram inicialmente submetidos ao teste de Kolmogorov-Smirnov, para verificação do tipo de distribuição. Dados com distribuição normal foram expressos como média ± desvio-padrão, e os com distribuição livre como mediana (P25; P75). Utilizou-se ANOVA, seguida pelo teste de Tukey, para fazer comparações entre os tanques de dados que tinham distribuição normal. Já os dados com distribuição livre foram comparados através do teste de Qui-Quadrado. Em todos os testes considerou-se 5% como nível de significância.
RESULTADOS
O Quadro 1 apresenta os resultados referentes aos parâmetros físico-químicos observados no momento da coleta dos animais de cada um dos tanques. Apesar das alterações físico-químicas observadas na água destes tanques, não se constatou nenhuma alteração significativa quanto ao peso, comprimento total, comprimento padrão e altura dos animais neles mantidos (Quadro 2). A densidade populacional de células mucosas em todo filamento branquial de todos os animais de cada tanque são apresentadas no Quadro 3.
A área de lamelas por região do filamento branquial observada nos animais de cada tanque está apresentada no Quadro 4. A densidade populacional de células mucosas por região do filamento branquial está apresentada no Quadro 5. A proporção de células mucosas produtoras de mucinas ácidas em relação às produtoras de mucinas neutras está apresentada no Quadro 6.
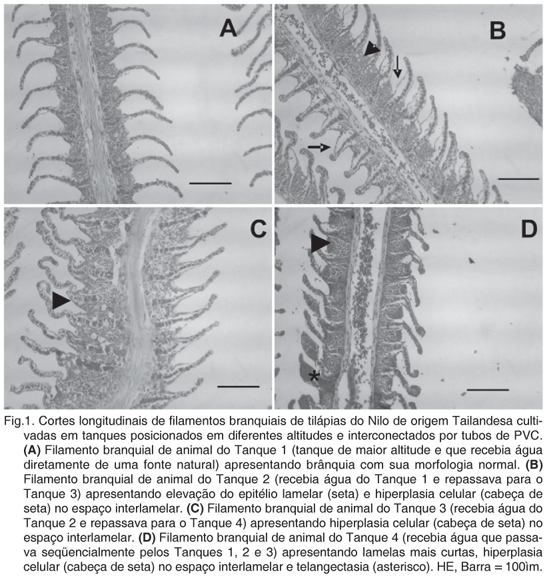
Pela análise morfológica foram evidenciadas alterações histológicas nas brânquias como elevação epitelial, hiperplasia, fusão lamelar e telangectasia (Fig.1).
DISCUSSÃO
Como os tanques observados neste estudo foram construídos em diferentes altitudes, mantendo-se conectados por tubulação de PVC, a mesma água passava por todos os tanques, sob propulsão de energia mecânica (com predominância do seu componente gravitacional). Quando se avaliou os parâmetros físico-químicos (temperatura, pH e concentração de oxigênio dissolvido) do ambiente aquático de cada tanque, percebeu-se que os valores decresciam à medida que passava de um tanque para o outro. Desta forma, considerou-se que os animais do tanque de maior altitude (T1) e que recebiam água de uma fonte natural (nascente) eram mantidos num ambiente aquático mais próximo das condições ideais para o cultivo de tilápias nilóticas. Estes animais foram utilizados, portanto, como parâmetro de comparação do que é normal para esta espécie.
Apesar das flutuações físico-químicas observadas, nenhuma alteração externa foi evidenciada nos animais coletados dos diferentes tanques, ou seja, estes fatores aparentemente não interferiram no desenvolvimento dos animais que estavam sendo mantidos em cultivo intensivo, visto que, não houve diferença significativa entre o peso, comprimento total e padrão e a altura dos animais entre os diferentes tanques (p>0,05). Considerando que os parâmetros ideais para o cultivo de tilápias são: temperatura 26-28°C, concentração de oxigênio dissolvido 3-6mg/l e pH 7,0-8,6 (Hein & Brianese 2004), observou-se que os tanques deste estudo estavam dentro destes limites, inclusive o tanque 4 (o que recebia a água depois de passada por todos os demais), apesar de ter uma baixa concentração de oxigênio dissolvida na água (2,8mg/l). Corrobora-se aos achados do presente trabalho, um estudo realizado para avaliar os parâmetros para a criação de tilápias nilóticas em gaiolas, no qual se observou que as variações encontradas quanto ao nível de oxigênio dissolvido (5,8±1,9mg/l), pH (6,6±0,4) e temperatura (27,52,8ºC) da água não tiveram influência significativa no desempenho dos peixes (Bozano et al. 1999). É importante ainda lembrar que, a densidade de estocagem tem efeito significativo sobre a taxa de crescimento, consumo de alimento e oxigênio (Zuanon et al. 2004), e se recomenda que esta não ultrapasse cinco peixes por m2 (Proença & Bittencourt 1994), o que também foi observado neste estudo.
Mesmo não sendo possível observar sinais de estresse pelas medidas corpóreas dos peixes, sabe-se que alterações anatômicas só ocorrem quando animais são submetidos a um grau importante de estresse em longo prazo. Antes de se observar uma lesão macroscopicamente, alterações na biologia das células e tecidos acontecem no intuito de manter a homeostasia do animal e/ou prepará-lo, para se defender de uma situação que o ameace (Mallatt 1985).
Neste sentido, indutores de irritação branquial têm sido investigados com muita freqüência, através de análises microscópicas, tais como baixa qualidade da água com pH e temperaturas extremas (Mallatt 1985), agentes tóxicos e irritantes como metais pesados e pesticidas (Schwaiger et al. 1997, Pane et al. 2004, Figueiredo-Fernandes et al. 2007, Garcia-Santos et al. 2007, Fontaínhas-Fernandes et al. 2008), agentes infecciosos e presença de parasitas (Pavanelli et al. 2002).
A análise morfométrica de lamelas branquiais permite avaliar de forma direta a relação do peixe com o ambiente aquático no que tange sua homeostasia, visto que é esperado que a área lamelar seja maior quando há condições favoráveis para o que o animal tenha maior interação com o ambiente aquático, e o oposto ocorre quando o animal está num ambiente mais hostil, seja pela presença de agentes químicos ou biológicos irritantes, como também variações de características físicas. Essa propriedade das lamelas branquiais têm sido utilizada em estudos de farmacognosia no intuito de se avaliar a toxicidade de compostos fitoquímicos sobre células de origem animal (Falkenberg et al. 2003).
Neste sentido, os resultados quanto à área lamelar deste estudo, independente das regiões do filamento, demonstraram um aumento progressivo da extensão desta estrutura nos animais do T1 ao T3, e um pequeno declínio nos animais do T4. Isto pode significar um aumento da superfície de contato do epitélio branquial com o meio aquático no intuito de captar mais gás oxigênio, já que a concentração desta molécula reduzia de um tanque para o outro. Porém no T4, embora tivesse pouco oxigênio dissolvido, a redução da área lamelar pode significar um mecanismo de contenção do animal frente a agentes irritantes presentes no ambiente aquático, como por exemplo metais pesados ou outros elementos que por lixiviação a partir de propriedades vizinhas poderiam ter sido escoados para o tanque (já que era o tanque de menor altitude). Resultados semelhantes foram encontrados por Pane et al. (2004) quando utilizaram peixes expostos a níquel, no qual observaram alterações significativas na ultraestrutura das lamelas, que induziram um aumento em número e extensão.
Contudo, quando se observou as lamelas considerando as diferentes regiões do filamento branquial, notou-se nos animais do T2 que a área lamelar aumentou progressivamente, da região mais proximal do arco branquial em sentido à ponta do filamento (sem diferença significativa para a região basal). Praticamente o oposto ocorreu nos animais do T4, que tinham lamelas de maior área na região basal. Esses resultados se distanciam dos achados por Silva et al. (2002), que utilizaram peixes expostos a frações da folha e da casca do caule do pequi (Caryocar brasilienses), e não encontraram diferenças significativas entre as três regiões quando comparadas ao grupo controle, sendo provável que as concentrações usadas no experimento não foram suficientes para provocar alterações na morfometria lamelar.
As lamelas branquiais são os locais onde ocorre a troca efetiva entre elementos químicos do sangue e da água, sobretudo, promovendo a hematose (Leonardo et al. 2001). Considerando que a concentração de oxigênio dissolvido era cada vez menor com a passagem da água de um tanque para o outro, é razoável considerar que uma ampliação da área das lamelas seja necessária para manter o nível de O2 circulante no sangue dos animais. Por outro lado, era o T4 que tinha o menor nível de O2 dissolvido na água e, contraditoriamente, os animais deste tanque tinham lamelas com área semelhante aos do T1, exceto às que estavam na região basal do filamento. Sugere-se que isso provavelmente ocorreu em função da possível existência de outros componentes do ambiente aquático, que eram irritantes para os animais deste tanque, e que desta maneira os forçavam a manter uma área lamelar mais reduzida. Por outro lado, já que as lamelas da região basal estão posicionadas mais distalmente do ambiente aquático externo e de forma mais proximal da água que entra pela faringe do animal, possivelmente sua maior extensão nos animais do T4 seja explicada pelo fato de terem uma maior responsabilidade na captação do pouco O2 existente na água logo que era ingerida. Isso é corroborado pelo fato de que o fluxo de água, através das diferentes porções do filamento (basal, intermediária e apical) varia de acordo com o volume ventilatório do animal, e sob ventilação moderada existe uma grande resistência para o fluxo na ponta do filamento (Severi et al. 2000). Por isso as lamelas localizadas perto da ponta do filamento tendem a ser mais curtas que aquelas da base, fato este que explica o encontrado nos animais do T4.
A superfície das brânquias de peixes é coberta por um muco, que contribui para proteção do animal quanto às flutuações dos parâmetros físicos, químicos e biológicos do ambiente aquático. E esta forma de proteção pode ser avaliada através do número das células produtoras das glicoproteínas (mucinas) que compõem este muco, assim como, pela análise das características químicas de seus componentes (Fernandes & Perna-Martins 2001, Breseguelo et al. 2004). Neste sentido, calculou-se a densidade populacional de células mucosas em todo o filamento branquial, e observou-se que houve aumento expressivo do número de células produtoras de mucinas neutras nos animais do T3 em relação ao T2 e T1 (p<0,05), contudo não houve diferença no número destas células nos animais do T4 (p>0,05). Quanto às células produtoras de mucinas ácidas (sialomucinas e sulfomucinas), observou-se um aumento em sua densidade populacional nos animais do T2, e uma redução progressiva nos peixes do T3 (que também demonstram um aumento em relação aos animais do T1) e T4 (p<0,05). Já o número de células produtoras de mucinas ácidas sulfatadas não foi alterado (p>0,05). Quando se avaliou a proporção de células produtoras de mucinas ácidas em relação ao número de células produtoras de mucinas neutras, constatou-se que nos animais do T2 esta proporção aumentou em relação aos animais do T1 e T3 (p<0,05), contudo nenhuma variação desta razão foi observada nos animais do T4 (p>0,05). Mucinas ácidas são mais solúveis do que as neutras quando entram em contato com o meio aquático, visto que possuem maior quantidade de cargas elétricas residuais (Myers et al., 2008). Desta forma, sugere-se que os animais do T3 tinham um maior volume de muco sobre o epitélio branquial, contudo, sem diferença na composição química em relação aos peixes do T1. Por outro lado, é importante destacar o aumento expressivo da proporção de mucinas ácidas em relação às neutras observado nos animais do T2, fato que indica que nestes animais o muco que recobria as brânquias estava mais fluido em relação ao dos animais dos outros tanques.
Mas quando se comparou quantitativamente o número de células mucosas, considerando as regiões do filamento branquial (basal, intermediária e apical), entre os animais dos diferentes tanques, encontrou-se hiperplasia de células produtoras de mucinas ácidas (sialomucinas + sulfomucinas) somente na região apical dos animais do T2 e hiperplasia de células produtoras de mucinas neutras somente na região apical dos animais do T3. Contudo um aumento da proporção entre mucinas ácidas e neutras por região do filamento branquial foi evidenciado tanto na região apical como basal dos animais do T2 (p<0,05), o que de fato indica que o muco nestas duas regiões estava mais fluido nestes animais, o que é indicativo de que a fisiologia dos peixes do T2 permitia, nesta situação, uma maior interação do epitélio branquial e do ambiente aquático. Como o T4 tinha o menor nível de oxigênio dissolvido na água, esperava-se observar alterações morfológicas nos animais deste tanque, que corroborassem com uma maior interação entre o epitélio branquial lamelar e o meio aquático, o que pode ser observado através da extensão da área desta estrutura na região basal dos filamentos.
Sabe-se que a proliferação e hipersecreção das células mucosas nas brânquias frente a situações de estresse, podem ser compreendidas como um mecanismo de defesa em função de uma resposta defensiva crônica, entretanto, podem comprometer a função branquial dependendo da severidade do processo (Mallatt 1985, Leonardo et al. 2001, Fracácio et al. 2003). Neste sentido, Breseghelo et al. (2004) expuseram peixes a flúor, e observaram hiperplasia das células mucosas com maior intensidade de glicoconjugados neutros no grupo controle e maior intensidade de glicoconjugados ácidos carboxilados no grupo experimental. Fracácio et al. (2003) analisaram a toxicidade de amostras de sedimentos coletados do rio Tietê (São Paulo, Brasil), para brânquias de Danio rerio e observaram hipertrofia das células mucosas. Leonardo et al. (2001) alimentaram larvas de tilápias com diferentes níveis de vitamina C, e observaram hipertrofia e hiperplasia das células mucosas nos filamentos branquiais das larvas que não ingeriram vitamina C. Berntssen et al. (1997) expuseram salmão atlântico (Salmo salar) em ambientes aquáticos com diferentes concentrações de alumínio e observaram um aumento do número de células mucosas contendo mucinas ácidas na brânquia. Martins et al. (2001) analisaram brânquias de várias espécies como tambacu híbrido, pacu, tambaqui, piauçu, tilápia e curimbatá infectado por parasitas (dinoflagelados), e constataram aumento da produção de muco em todas as espécies. Resultado semelhante quanto ao número de células mucosas foi encontrado por Dezfuli et al. (2003), utilizando a espécie Abramis brama infectada por ectoparasitas (Ergasilus sieboldi). Desta forma, a literatura mostra que as alterações na morfometria e no número de células mucosas dependem do agente, cujo os peixes são expostos. No presente estudo, considera-se que outros fatores (além dos físico-químicos observados) podem ter influenciado nos achados apresentados quanto ao número destas mucosas, já que não se pode desconsiderar a possibilidade da presença de parasitas, metais pesados contidos nos sedimentos, além do fato de que a região do pesque-pague alvo desta pesquisa é margeada por fazendas com atividade pecuária.
Na análise qualitativa, constatou-se que o ambiente aquático influenciou na morfologia tecidual das brânquias entre os diferentes tanques. Nos animais do T1, que tinham um ambiente aquático dentro do ideal para o cultivo de tilápias nilóticas, constatou-se o aspecto normal da morfologia de brânquias de teleósteos: quatro arcos branquiais funcionais, contendo vários filamentos dos quais partiam perpendicularmente as lamelas. Nos animais dos outros tanques, notaram-se alterações morfológicas, cada vez mais intensas em relação às alterações físico-químicas do meio aquático, ou seja, progressivamente havia maior descolamento do epitélio branquial, hiperplasia celular no espaço interlamelar, e nos animais do T4 observou-se com freqüência telangectasias. Resultados semelhantes são observados quando peixes são submetidos a estresse. Córdova et al. (1996) observaram hiperplasia na ponta dos filamentos e nas lamelas em tilápias confinadas em um ambiente com espaço reduzido. Schwaiger et al. (1997) expuseram juvenis de truta marrom a metais pesados e pesticidas e constataram proliferação de células epiteliais dos filamentos e lamelas branquiais, fusão lamelar e hiperplasia de células mucosas. Fracácio et al. (2003) observaram hiperplasia e fusão lamelar em larvas de Danio rerio expostas a sedimentos tóxicos do rio Tietê (São Paulo, Brasil). Leonardo et al. (2001) trabalhando com larvas de tilápias alimentadas com diferentes níveis de vitamina C, também evidenciaram elevação epitelial, hiperplasia das células dos filamentos, telangectasia e fusão lamelar. Os resultados observados neste estudo confrontados com a literatura demonstram que não é possível concluir que estes achados se devam especificamente às variações físico-químicas observadas no meio aquático dos tanques avaliados, visto que a literatura descreve o surgimento destas alterações morfológicas quando peixes são expostos ao estresse, metais pesados, sedimentos tóxicos e elementos da dieta.
CONCLUSÕES
O declínio da temperatura, do pH e da concentração de oxigênio dissolvido observado na água de tanques de cultivo intensivo das tilápias avaliadas neste estudo provocou inicialmente uma extensão da área lamelar e do número de células mucosas branquiais.
A contínua queda destes parâmetros físico-químicos induziu uma contenção tanto da área lamelar quanto do número de células mucosas.
Houve um progressivo aumento de alterações morfológicas do epitélio branquial (hiperplasia interlamelar, fusão lamelar, descolamento epitelial e telangectasias) quando foram observados os peixes cultivados em tanques com condições menos favoráveis.
Os achados deste estudo também podem estar relacionados à presença de parasitas ou elementos químicos tóxicos presentes no ambiente aquático, os quais não foram investigados.
Agradecimentos.- Os autores agradecem à mestranda do Programa em Ciência Animal da Unipar Catchia Hermes, pelo auxílio na análise estatística, e á Universidade Paranaense pelo financiamento integral da pesquisa.
Recebido em 21 de abril de 2008.
Aceito para publicação em 7 de janeiro de 2009.
- Araújo E.J.A., Morais J.O.R., Souza P.R. & Sabóia-Morais S.M.T. 2001. Efeito de poluentes químicos cumulativos e mutagênicos durante o desenvolvimento ontogenético de Poecilia vivipara (Cyprinodontiformes, Poeciliidae). Acta. Sci. 23:391-399.
- Araújo E.J.A., Sabóia-Morais S.M.T., Morais J.O.R. & Mata, J.R. 1997. Effects of irritants agents on surface ultrastucture of gill rays and filaments of guppy (Poecilia vivipara). Acta Microsc. 6:700-701.
- Araújo E.J.A., Nascimento O.R., Pires D.R., Carneiro C.E.A., Silva L.D., Ferreira R.N., Souza P.R., Morais J.O.R. & Sabóia-Morais S.M.T. 1999. Toxic action of heavy metals on the brachial epithelium of guppies (Poecilia vivipara). Acta Microsc. 8:355-356.
- Araújo E.J.A., Carneiro C.E.A. & Sabóia-Morais S.M.T. 2000. Reação das células transportadoras de íons de brânquias de peixes adultos após exposição a acetato de chumbo. Braz. J. Morph. Sci. 17:94-94.
- Berntssen M.H.G., Kroglund F., Rosseland B.O. & Wendelaar Bonga, S.E. 1997. Responses of skin mucous cells to aluminium exposure at low pH in Atlantic salmon (Salmo salar) smolts. Can. J. Fish. Aquat. Sci. 54:1039-1045.
- Bozano G.L.N., Rodrigues S.R.M., Caseiro A.C. & Cyrino J.E.P. 1999. Desempenho da tilápia nilótica Oreochromis niloticus (L.) em gaiolas de pequeno volume. Sci. Agric. 56:819-825.
- Breseghelo L., Cardoso M.P., Borges-de-Oliveira R., Costa M.F., Barreto J.C.B., Sabóia-Morais S.M.T. & Yamada A.T. 2004. Efeitos do fluoreto de sódio no epitélio da brânquia do peixe guaru (Poecilia vivipara). Braz. J. Vet. Res. Anim. Sci. 46:685-695.
- Carvalho A.R. & Oliveira M.V.C. 1997. Princípios Básicos do Saneamento do Meio. Senac, São Paulo. 132p.
- Cavichiolo F. 2000. Efeito da vitamina C (ácido ascórbico) na ocorrência de ectoparasitas e nas alterações histológicas em alevinos de tilápia do Nilo (Oreochromis niloticus). Dissertação de Mestrado em Zootecnia, Universidade Estadual de Maringá, Paraná. 61p.
- Córdova M.S., Ocampo A.A & Argüero N.B. 1996. Lesiones histopatológicas producidas en la tilapia (Oreochromis sp.) por el confinamiento experimental en acuario. Vet. Mex. 27:143-148.
- Dezfuli B.S., Giari L., Konecny R., Jaeger P. & Manera M. 2003. Immunohistochemistry, ultrastructure and pathology of gills of Abramis brama from lake Mondsee, Austria, infected with Ergasilus sieboldi (Copepoda). Dis. Aquat. Org. 53:257-262.
- Falkenberg M.B., Santos R.I. & Simões C.M.O. 2003. Introdução à análise fitoquímica, p.229-245. In: Simões C.M.O. (Ed.), Farmacognosia: da Planta ao Medicamento. 5Ş ed. UFRGS/UFSC, Porto Alegre.
- Fernandes M.N. & Perna-Martins S.A. 2001. Epithelial gill cells in the armored catfish, Hypostomus cf. plecostomus (Loricariidae). Revta Bras. Biol. 61:69-78.
- Figueiredo-Fernandes A., Ferreira-Cardoso J.V., Garcia-Santos S., Monteiro S.M., Carrola J., Matos P. & Fontainhas-Fernandes A. 2007. Histopathological changes in liver and Gill epithelium of Nile tilapia, Oreochromis niloticus, exposed to waterborne copper. Pesq. Vet. Bras. 27:103-109.
- Fontainhas-Fernandes A., Luzio A., Garcia-Santos S., Carrola J. & Monteiro S. 2008. Gill histopathological alterations in Nile tilápia, Oreochromis niloticus, exposed to treated sewage water. Braz. Arch. Biol. Technol. 51:1057-1063.
- Fracácio R., Verani N.F., Espíndola E.L.G., Rocha O., Rigolin-Sá O. & Andrade C.A. 2003. Alterations on growth and gill morphology of Danio rerio (Piscis, ciprinidae) exposed to the toxic sediments. Braz. Arch. Biol. Technol. 46:685-695.
- Garcia-Santos S., Monteiro S.M., Carrola J. & Fontainhas-Fernandes A. 2007. Alterações histológicas em brânquias de tilápias nilóticas Oreochromis niloticus causadas pelo cádmio. Arq. Bras. Med. Vet. Zootec. 59:376-381.
- Hein G. & Brianese R.H. 2004. Modelo Emater de produção de tilápia. Disponível em <http://www.emater.pr.gov.br/arquivos/File/Comunicacao/Premio_ Extensao_Rural/1_Premio_2005/ModeloEmaterProd_Tilapia.pdf >. Acesso em 2 de abril de 2008.
- Hermanns G. & Bellé T.L. 2006. Desenvolvimento e aceitação de formulações à base de peixe cultivado. Revta Hig. Aliment. 20:48-52.
- Leonardo J.M.L.O. 1999. Efeito da Vitamina C em larvas de tilápia do Nilo (Oreochromis niloticus) de origem tailandesa. Dissertação de Mestrado em Zootecnia, Universidade Estadual de Maringá, Paraná. 55p.
- Leonardo J.M.L.O., Vargas L., Ribeiro R.P., Moreira H.L.M., Natali M.R.M., Volski T. & Cavichiolo F. 2001. Histologia das Brânquias de larvas de tilápia do Nilo, Oreochromis niloticus (L.) de origem tailandesa, submetidas a diferentes níveis de vitamina C. Acta Sci. 23:863-870.
- Lopes F.F. 2006. Monitoramento ambiental da bacia hidrográfica do lago Guaíba-RS, Brasil, através da utilização de diferentes metodologias aplicadas a taxocenoses de peixes. Tese de Doutorado em Biologia Animal, Universidade Federal do Rio Grande do Sul, Rio Grande do Sul. 234p.
- Maciel-Filho A.A., Góes-Júnior C.D., Câncio J.A., Heller L., Moares L.R.S., Carneiro M.L. & Costa S.S. 2003. Interfaces da gestão de recursos hídricos e saúde pública. Disponível em: <http://portal.saude.gov.br/portal/arquivos/PDF/interfaces.pdf>. Acesso em 2 de abril de 2008.
- Mallatt J. 1985. Fish gill structural changes induced by toxicants and other irritantes: a statistical Review. Can. J. Fish. Aquat. Sci. 42:630-648.
- Martins M.L., Moraes J.R.E., Andrade P.M., Schalch S.H.C. & Moares F.R. 2001. Piscinoodinium pillulare (Schaperclaus, 1954) Lom, 1981 (Dinoflagellida) infection in cultivated freshwater fish from the northeast region of São Paulo State, Brazil: Parasitological and pathological aspects. Braz. J. Biol. 61:639-644.
- Mellanby K. 1982. Biologia da Poluição. Vol.28. EPU, São Paulo. 89p.
- Moraes D.S.L. & Jordão B.Q. 2002. Degradação de recursos hídricos e seus efeitos sobre a saúde humana. Rev. Saúde Pública 36:370-374.
- Myers B.M., Fredenburgh J.L. & Grizzle W.E. 2008. Theory and practice of histological techniques, p.161-187. In: Bancroft J.D. & Gamble M. (Eds), Carbohydrates. 6th ed. Elselvier, Philadelphia.
- Pane E.F., Haque A., Goss G.G. & Wood C.M. 2004. The Physiological consequences of exposure to chronic, sublethal waterborne nickel in rainbow trout (Oncorhynchus mykiss): exercise vs resting physiology. J. Exp. Biol. 207:1249-1261.
- Pavanelli G.C., Takemoto R. & Eiras J. 2002. Doença de Peixes: profilaxia, diagnóstico e tratamento. EDUEM, Maringá, Paraná. 305p.
- Proença M.C.E. & Bittencourt L.P.R. 1994. Manual de Pscicultura Tropical. Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis. IBAMA, Brasil. 196p.
- Sabóia-Morais S.M.T., Santos S.C., Carneiro C.E.A., Silva R.M., Pires D.R., Araújo E.J.A., Morais J.O.R. 1999. Evaluation of the effect of extracts of cerrado plants of gill cells of guppies (Poecilia vivipara) used as biomonitors. Acta. Microsc. 8:357-358.
- Schwaiger J., Wanke R., Adam S., Pawert M., Honnen W. & Triebskorn R. 1997. The use of histopathological indicators to evaluate contaminant-related stress in fish. J. Aquat. Eco. Stress Recov. 6:75-86.
- Severi W., Rantin F.T. & Fernandes M.N. 2000. Structural and morphological features of Piaractus mesopotamicus (Holmerg, 1887) gills. Revta Bras. Biol. 60:493-501.
- Silva L.D., Rosa E.V., Santos S.C. & Sabóia-Morais S.M.T. 2002. Análise morfométrica do epitélio de revestimento do filamento branquial do guaru (Poecilia vivipara) exposto a frações do extrato da folha e casca do caule de pequi (Caryocar brasiliensis). Arq. Ciênc. Saúde Unipar 6:101-106.
- Suarez-Mahecha H., Francisco A., Beirão L.H., Block J.M., Saccol A. & Pardo-Carrasco S. 2002. Importância de ácidos graxos poliinsaturados presentes em peixes de cultivo e de ambiente natural para a nutrição humana. Bolm Inst. Pesca 28:48-52.
- Winkaler E.U., Silva A.G., Galindo H.C. & Martinez C.B.R. 2001. Biomarcadores histológicos e fisiológicos para o monitoramento da saúde de peixes de ribeirões de Londrina, estado do Paraná. Acta Sci. 23:507-514.
- Zuanon J.A.S., Assano M. & Fernandes J.B.K. 2004. Desempenho de tricogaster (Trichogaster trichopterus) submetido a diferentes níveis de arroçamento e densidades de estocagem. Revta Bras. Zootec. 33:1639-1645.
Datas de Publicação
-
Publicação nesta coleção
08 Jul 2009 -
Data do Fascículo
Abr 2009
Histórico
-
Recebido
21 Abr 2008 -
Aceito
07 Jan 2009