Resumos
O timo é um órgão linfático primário que desenvolve sua atividade em organismos jovens. Apesar de sua função ser responsável por mecanismos fundamentais na aquisição das defesas e conseqüentes respostas orgânicas, ela ainda não está totalmente esclarecida, nem tampouco as bases morfológicas que respondem por tais funções, como o processo de desenvolvimento e involução do órgão. Objetivou-se analisar e caracterizar os aspectos morfológicos do timo, tais como seu tamanho e volume, e aspectos histológicos do timo em gatos, correlacionando o sexo e o desenvolvimento etário. Doze timos provenientes de fetos de gatos domésticos (Felis domesticus) sem raça definida (SRD), machos e fêmeas, separados em três grupos etários. O timo apresentou-se com uma coloração rosa-pálida e com duas porções, a torácica e a cervical, sendo que cada uma delas possuía um lobo direito e um lobo esquerdo em sua maioria. A porção torácica localizava-se em região de mediastino cranial, entre os pulmões e à base do coração. E a porção cervical estendia-se além das costelas em sentido cranial, estando localizada ventralmente à traqueia. A estrutura celular do timo demonstrou-se organizada com a presença de agregados concêntricos, os chamados corpúsculos tímicos, formados por células epiteliais, sustentada por uma cápsula de tecido conjuntivo de onde partiam septos que ao penetrar no órgão dividia-o em lóbulos. Ocorreram variações significativas quanto à lobação e as dimensões do timo entre indivíduos da mesma faixa etária, e entre sexos diferentes. Os valores relativos ao comprimento, espessura e largura, de maneira geral, apresentaram aumento, em conformidade ao desenvolvimento dos animais, mas com diferenças entre os sexos.
Timo; morfometria; órgão linfóide; linfócitos T; felinos
The thymus is a primary lymphatic organ that develops its activity in young organisms. But despite its function is fundamental mechanisms responsible for the acquisition and subsequent body defenses, it is not yet fully understood, neither the morphological basis accounting for such functions as the process of development and the organ involution. The objective was to analyze and characterize the morphology of the thymus, such as its size and volume, and histological aspects of the thymus in cats, correlating sex and age development. Twelve foetus of mongrel domestic cats (Felis domesticus), males and females, divided into three age groups. The thymus presented two parts with a pale pink color, the thoracic and cervical portion; each has a right lobe and a left lobe mostly. The thoracic portion was located in the region of cranial mediastinum, between the lungs and the heart base. And the cervical portion extended beyond the ribs cranially and is located ventral to the trachea. The thymus cellular structure was shown by the presence of organized concentric aggregates, named Hassall's corpuscles, formed by epithelial cells, supported by a connective tissue capsule that penetrated the parenchyma dividing it into lobules. Significant changes with number of lobes and thymus size between individuals of the same age, and between sex. The values for length, thickness and width, in general, showed an increase, according to the development of animals, with differences between sex.
Thymus; morphometry; linfoid organ; T lymphocyte; Felis domesticus cats
MORFOFISIOLOGIA
Aspectos morfométricos do timo em gatos domésticos (Felis domesticus)
Morphometric aspects of the thymus in domestic cats (Felis domesticus)
Camila E. BarrosoI,* * Autor para correspondência: camilaercolini@gmail.com ; Fernanda R. AgresteII; Luana C.S. SilvaIII; Pedro P. BombonatoI; Bruno CogliatiII
IDepartamento de Cirurgia, Faculdade de Medicina Veterinária e Zootecnia (FMVZ), Universidade de São Paulo (USP), Av. Prof. Orlando Marques de Paiva 87, Cidade Universitária, São Paulo, SP 05508-270, Brasil
IIDepartamento de Patologia Experimental e Comparada, Faculdade de Medicina Veterinária e Zootecnia (FMVZ), Universidade de São Paulo (USP), Av. Prof. Orlando Marques de Paiva 87, Cidade Universitária, São Paulo, SP 05508-270, Brasil
IIIMédica Veterinária, Prefeitura de Pomerode, Rua 15 de novembro 525, Pomerode, SC 89107-000, Brasil
RESUMO
O timo é um órgão linfático primário que desenvolve sua atividade em organismos jovens. Apesar de sua função ser responsável por mecanismos fundamentais na aquisição das defesas e conseqüentes respostas orgânicas, ela ainda não está totalmente esclarecida, nem tampouco as bases morfológicas que respondem por tais funções, como o processo de desenvolvimento e involução do órgão. Objetivou-se analisar e caracterizar os aspectos morfológicos do timo, tais como seu tamanho e volume, e aspectos histológicos do timo em gatos, correlacionando o sexo e o desenvolvimento etário. Doze timos provenientes de fetos de gatos domésticos (Felis domesticus) sem raça definida (SRD), machos e fêmeas, separados em três grupos etários. O timo apresentou-se com uma coloração rosa-pálida e com duas porções, a torácica e a cervical, sendo que cada uma delas possuía um lobo direito e um lobo esquerdo em sua maioria. A porção torácica localizava-se em região de mediastino cranial, entre os pulmões e à base do coração. E a porção cervical estendia-se além das costelas em sentido cranial, estando localizada ventralmente à traqueia. A estrutura celular do timo demonstrou-se organizada com a presença de agregados concêntricos, os chamados corpúsculos tímicos, formados por células epiteliais, sustentada por uma cápsula de tecido conjuntivo de onde partiam septos que ao penetrar no órgão dividia-o em lóbulos. Ocorreram variações significativas quanto à lobação e as dimensões do timo entre indivíduos da mesma faixa etária, e entre sexos diferentes. Os valores relativos ao comprimento, espessura e largura, de maneira geral, apresentaram aumento, em conformidade ao desenvolvimento dos animais, mas com diferenças entre os sexos.
Termos de indexação: Timo, morfometria, órgão linfóide, linfócitos T, felinos.
ABSTRACT
The thymus is a primary lymphatic organ that develops its activity in young organisms. But despite its function is fundamental mechanisms responsible for the acquisition and subsequent body defenses, it is not yet fully understood, neither the morphological basis accounting for such functions as the process of development and the organ involution. The objective was to analyze and characterize the morphology of the thymus, such as its size and volume, and histological aspects of the thymus in cats, correlating sex and age development. Twelve foetus of mongrel domestic cats (Felis domesticus), males and females, divided into three age groups. The thymus presented two parts with a pale pink color, the thoracic and cervical portion; each has a right lobe and a left lobe mostly. The thoracic portion was located in the region of cranial mediastinum, between the lungs and the heart base. And the cervical portion extended beyond the ribs cranially and is located ventral to the trachea. The thymus cellular structure was shown by the presence of organized concentric aggregates, named Hassall's corpuscles, formed by epithelial cells, supported by a connective tissue capsule that penetrated the parenchyma dividing it into lobules. Significant changes with number of lobes and thymus size between individuals of the same age, and between sex. The values for length, thickness and width, in general, showed an increase, according to the development of animals, with differences between sex.
Index terms: Thymus, morphometry, linfoid organ, T lymphocyte, Felis domesticus cats.
INTRODUÇÃO
O timo constitui-se num órgão linfático primário que desenvolve sua atividade em organismos jovens (König & Liebich 2004). Suas células T iniciam seu desenvolvimento como precursoras na medula óssea e migram para o timo, aonde sofrem divisão, diferenciação e maturação em células T funcionais (Bar-Dayan et al. 1999). E a migração para os órgãos linfáticos secundários faz com que adquiram novas características e se transformem em células imunocompetentes (Silva et al. 2001). Diversos experimentos têm revelado que tanto o desenvolvimento normal quanto a manutenção de várias estruturas linfóides são dependentes de um órgão linfóide específico, ou seja, o timo. Assim as respostas imunológicas naturais ou induzidas, que estão de diferentes formas relacionadas às estruturas do sistema linfático, muitas vezes são intermediadas e coordenadas pelo timo (Bombonato 1997, Appolinário 1998).
O desenvolvimento do timo ocorre desde o terceiro arco faríngeo, de forma par e ao longo da traquéia em direção caudal, até o pericárdio (König & Liebich 2004). Advindo da terceira e quarta bolsa faríngea e acompanhando os derivados paratireoideanos dessas bolsas. O broto tímico então se separa desses derivados, vindo a ocupar parte do mediastino cranial, desde a entrada do tórax à região cervical ventral (Banks 1992). Ainda na vida fetal e no período neonatal o timo detém uma alta importância imunológica e, anatomicamente, é o maior órgão linfático. Dentre todos os órgãos linfóides, ele é o que possui a maior atividade linfopoiética (Drummond 1996) e demonstrando também relações com o sistema endócrino, principalmente com a função gonadal (Drummond 1996, Appolinário 1998).
Segundo Silva et al. (2001) a topografia do timo nos felinos demonstrou a presença de duas porções, a torácica e a cervical, em que esta última, quando presente apresentava-se fina e estreita, representando não mais que um quinto a um sexto do tecido tímico total, e pronunciou-se cranialmente ao primeiro par de costelas e ventral à traqueia. A forma deste órgão varia individualmente, mesmo entre irmãos e irmãs, e também conforme a idade do felino, chegando a atingir o seu tamanho absoluto na puberdade (Daschinger 1983). Macroscopicamente este órgão no gato doméstico recém-nascido possui um aspecto visivelmente lobular e de coloração rosa-pálida, que se estende cranialmente do manúbrio do esterno pela abertura torácica cranial e ventralmente preenche dois terços do mediastino cranial (Daschinger 1983). E em filhotes, se estende cranialmente à região torácica e caudalmente entre as veias braquiocefálica e entre a quinta e sexta cartilagem intercostal, onde repousa na superfície ventral do pericárdio. Tendo a região caudal do órgão com o lóbulo direito menor que o esquerdo (Hudson & Hamilton 1993).
Daschinger (1983) descreveu que o suprimento arterial de todo o timo de gatos recém-nascidos provinha da A. torácica interna direita que se ramifica dando origem a um ramo caudal em direção ao timo torácico e um ramo craniomedial ao timo cervical.
Quando o animal adquire sua maturidade sexual o timo sofre uma involução significativa, a qual será distinta conforme a espécie animal (Banks 1992, König & Liebich 2004). Essa involução se dá de forma gradual e contínua, estando caracterizada pela diminuição do peso do órgão, pela perda dos linfócitos corticais, infiltração pelas células adiposas e o aumento do número dos corpúsculos tímicos. Que por fim culminam com a infiltração de células adiposas em substituição completa do órgão (Banks 1992). Desta forma este órgão é submetido à senescência de forma prematura, ou seja, é submetido aos danos que ocorrem com diversos outros órgãos quando do envelhecimento animal (George & Ritter 1996).
Apesar de a função tímica ser responsabilizada por mecanismos fundamentais na aquisição das defesas e consequentes respostas orgânicas, ela ainda não está totalmente esclarecida, e tampouco as bases morfológicas que respondem por tais funções, como o processo de desenvolvimento e involução (Bombonato 1997). Mas sabe-se que o desenvolvimento da imunocompetência animal é depende da diferenciação dos linfócitos do timo, os quais são capazes de reativar células imunes mediadas, em adição à diferenciação em outros locais de células que sintetizam e secretam anticorpos (Bockman 1997). Além disso, o papel significativo do timo na imunidade é facilmente demonstrado, em algumas espécies, quando da realização da timectomia neonatal, o que ocasiona o enfraquecimento da reação de hipersensibilidade retardada e a diminuição na habilidade para produzir respostas mediadas por anticorpos (Banks 1992).
Recentemente diversos morfologistas têm se beneficiado do uso de técnicas genéticas e moleculares que ajudam no esclarecimento de questões acerca dos órgãos linfóides e consequentemente melhorando assim as pesquisas biológicas e biomédicas. No entanto, assuntos concernentes a alterações de tecidos, células ou organelas celulares (as quais frequentemente aparecem em períodos de adaptação, evolução ou doença de um organismo), assim como uma melhor correlação entre a morfologia e função, necessitam ainda de uma análise quantitativa para um melhor entendimento (Mayhew 1992, Andersen & Pakkenberg 2003, Mandarim 2003).
Diante desses aspectos, objetivou-se analisar e caracterizar os aspectos morfológicos do timo em gatos, tais como seu tamanho e volume, e seus aspectos histológicos da vascularização, correlacionando o sexo e o desenvolvimento etário.
MATERIAL E MÉTODOS
Foram coletados 12 timos provenientes de fetos de gatos domésticos (Felis domesticus) sem raça definida (SRD), machos e fêmeas em igual número, com 35, 45 e 55 dias, separados em três grupos etários designados de Grupo I, II, III, respectivamente. Os timos removidos foram mensurados, com um paquímetro digital, em seu eixo longitudinal e transversal (região cranial e caudal), obedecendo ao sentido craniocaudal do mesmo, resultando nas medidas de comprimento, espessura e largura do órgão. O volume do órgão ou volume referência (V(ref)) foi estimado conforme o método de Cavalieri (Gundersen & Jensen 1987, Mayhew 1992). Em seguida foram fixados em solução de Karnovsky modificada por um período de 24 horas. Após a fixação estes foram seccionados transversalmente originando planos macroscópicos paralelos e seriados com distância média de 3 mm como descrito no trabalho de Mayhew (1992) e Wulfsohn et al. (2004). As fatias escolhidas aleatoriamente obedeceram ao método orientator modificado (Mattfeldt et al. 1990) e assim obteve-se cortes AUI (Aleatórios e Uniformemente Isotrópicos) ou IUR (Isotropic Uniform Random). Posteriormente tais fatias foram processadas para emblocagem com resina araldite (502 Polyscience Inc.®), cortados em cortes semi-finos no ultra-micrótomo MT-XL®, com 1 µm de espessura e corados com azul de toluidina para a análise histológica.
RESULTADOS
O timo nos gatos dos diferentes grupos etários analisados se apresentou com uma coloração rosa-pálida, localização no mediastino cranial entre os pulmões e dorsalmente à base do coração. Algumas particularidades foram encontradas em alguns fetos irmãos, em que houve variação da lobação do órgão entre estes, e entre faixas etárias diferentes, ocorrendo diferenciações quanto à morfologia e ao número de lobos tímicos.
No Grupo I (35 dias) um feto macho apresentou macroscopicamente os lobos torácicos divididos em direito e esquerdo em região de tórax e no final da porção cervical. Dois fetos machos apresentaram macroscopicamente uma única massa sem divisão com vestígio do lobo direito em região torácica. E o timo de uma fêmea apresentou-se macroscopicamente com os lobos torácicos totalmente divididos, em direito e esquerdo, unidos por um fino tecido conjuntivo. Neste caso, o lobo torácico direito estava dividido em porção cranial e caudal, enquanto que o esquerdo não possuía divisão, e assim aparecia como uma massa única. O lobo intermediário se apresentou emitindo tecido para a divisão em região cervical direita e esquerda. Duas fêmeas deste mesmo grupo apresentaram macroscopicamente uma massa única em região torácica totalmente dividida em lobo direito e esquerdo, indicando ser lobo torácico, sem vestígios ainda de lobo cervical.
No Grupo II (45 dias) não houve variação quanto à lobação entre os indivíduos.
No Grupo III (55 dias) o timo de uma fêmea apresentou macroscopicamente extensão do lobo torácico direito além do primeiro par de costelas em região cervical, com direção ao antímero direito. Neste mesmo grupo, dois timos de fêmeas, apresentaram também uma extensão que ultrapassava o 1º par de costelas, mas que se ramificava delgadamente ao antímero esquerdo, entre o esôfago e a traqueia cervical.
A porção cervical do timo relacionou-se ventralmente com o terço distal da traqueia cervical, aparecendo com uma forma fina e estreita. Foi possível observar a presença de sulcos em região de transição entre a porção cervical e a porção torácica, dividindo estes em lobos, e nestes sulcos notou-se a confluência da artéria e veia torácica interna. A porção esquerda do lobo tímico torácico, de maior tamanho quando comparado com o direito, se apresentava em região de 1º a 5º costela esquerda. Estando relacionada caudalmente com a aurícula direita do coração, dorsalmente com o esôfago torácico, e lateralmente, em região de incisura tímica entre os lobos cervicais e torácicos, observou-se adentrando à incisura a artéria torácica interna esquerda. Esta porção esquerda ainda relacionou-se ventral e medialmente com o lobo cranial do pulmão esquerdo. Já a porção tímica torácica direita relacionou-se caudalmente com o coração e medial e ventralmente ao lobo pulmonar cranial direito.
O timo encontrava-se revestido por uma cápsula de tecido conjuntivo que emitia invaginações formando septos que dividiam parcialmente o lobo formando em uma estrutura lobular. Os vasos sanguineos que adentravam os lobos juntamente com os septos se ramificavam no percurso dos septos interlobulares alcançando o parênquima tímico. O parênquima do órgão se apresentou bem heterogêneo em algumas regiões, devido a presença de células em diferentes estágios de maturação, e em outras regiões encontrava-se de forma homogênea. Os corpúsculos tímicos foram evidenciados em todos os grupos, no entanto no grupo III houve uma maior freqüência de aparecimento.
Nos animais dos Grupos I e II o timo apresentou-se de forma heterogênea com uma população de células se reorganizando, quando comparados com o grupo III, o qual se apresentou de forma mais organizada e homogênea. Os arranjos celulares do grupo I, II e III foram semelhantes entre si, sendo caracterizados por células mais agrupadas e com os vasos aumentando de calibre conforme o crescimento do animal (Fig.1). Não foram observadas diferenças morfológicas entre machos e fêmeas.
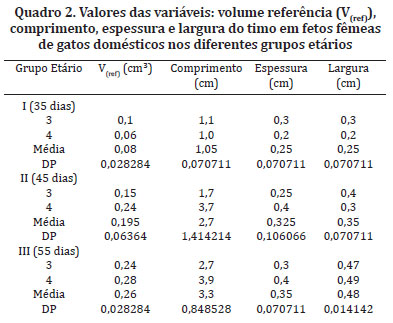
De forma geral o comprimento, a espessura e a largura do timo apresentaram um aumento gradativo conforme o avanço da faixa etária e desenvolvimento dos fetos analisados (Quadros 1 e 2).
O volume do órgão ou volume referência (Vref) estimado pelo método de Cavalieri apresentou valores que aumentavam gradualmente de acordo com o desenvolvimento dos fetos, sendo maior nas fêmeas dos Grupos I e III quando analisada a mesma faixa etária (Quadros 1 e 2). Houve diferença estatisticamente significante (p < 0,05) entre macho e fêmea do Grupo II (Mann-Whitney e Wilcoxon), e quando comparado às faixas etárias entre os Grupos I e II, I e III (Teste de Wilcoxon).
DISCUSSÃO
Escassos são os estudos referentes à histologia e à microvascularização especificos do timo de gatos, fato este que nos fez considerar outras espécies animais nas análises comparativas, considerando suas particularidades.
Quanto aos achados macroscópicos estes foram semelhantes aos relatados por Daschinger (1983) o qual descreveu que o timo possui uma coloração rosea-pálida e aspecto visivelmente lobular. A lobação do órgão observada no presente trabalho foi semelhante aos relatos de Daschinger (1983) e Silva et al. (2001) os quais relataram que o órgão possui duas porções, a torácica e a cervical, estando esta última de forma fina e estreita.
A localização do órgão foi observada em região do mediastino, estando cranial ao coração e ventral à traqueia, corroborando com a descrição feita por Daschinger (1983).
A forma deste órgão variou individualmente mesmo entre fetos da mesma ninhada, concordando assim com Daschinger (1983), e houve a observação de que o lobo direito é menor que o lobo esquerdo, achado este similar ao descrito por Hudson & Hamilton (1993).
Quanto à sintopia nossos achados foram semelhantes aos de Daschinger (1983), em que pôde-se notar que o nervo vago localizava-se dorsalmente à porção torácica, e que a transição dos lobos torácicos para os lobos cervicais encontrava-se demarcada por sulcos onde nos quais confluíam a artéria e veia torácica interna.
Kato (1997) e Junqueira & Carneiro (1999) relataram que durante o desenvolvimento do timo, ramos vasculares penetram no interior do parênquima acompanhando o tecido conjuntivo, formando um espaço de tecido conjuntivo perivascular, fato este que também foi observado em nosso trabalho. As arteríolas ramificavam-se e aprofundavam-se seguindo os septos conjuntivos, formando então capilares que penetravam na cortical, e depois se dirigiam para a região medular onde desembocavam em vênulas, as quais em seguida confluíam para formar veias e penetrar nos septos conjuntivos, saindo do timo pela cápsula.
No tocante ao suprimento sanguíneo do timo de gatos, Daschinger (1983) descreve que o mesmo é provido pela artéria torácica interna, confirmando o que foi observado em nosso trabalho. Já Silva et al. (2001) propuseram que a maior contribuição vascular para o timo de gatos decorre através dos ramos fornecidos pela artéria torácica interna esquerda e do tronco braquicefálico, sendo os ramos diretos destinados ao timo provenientes das artérias torácicas internas e carótidas comuns direita e esquerda e dos troncos bicarotídeos.
Quanto à histologia do órgão concordamos com Melo & Lage (1987) e Banks (1992) os quais descreveram que tanto a cápsula quanto o septo do timo são constituídos por tecido conjuntivo frouxo e tecido adiposo. Os corpúsculos tímicos denominados de corpúsculos de Hassals foram localizados em todas as faixas etárias estudadas, tendo células epiteliais dispostas concentricamente, como já relatado por Getty (1986) e Banks (1992).
Vizi et al. (1995), Leposavić et al. (2000), Savino & Dardene (2000) e Solarović et al. (2004) descreveram que a inervação dos órgãos linfóides, as interações entre os microambientes do timo e o desenvolvimento de células T, o qual é coordenado pelo sistema nervoso autônomo (SNA) e neuroendócrino, embora não tenham sido o objeto de estudo direto deste trabalho pode ser que possíveis alterações métricas do órgão, quando comparamos machos e fêmeas, podem ter sido ocasionados por influência do sistema neuroendócrino.
As mudanças no tamanho do timo quando da puberdade podem estar correlacionados com as alterações hormonais (Vizi et al. 1995, Olsen & Kovacs 1996, Kurz et al. 1997), e estes achados corroboram com a hipótese de Marchetti et al. (1990) os quais relataram que o sistema nervoso simpático está envolvido com a resposta da célula tímica à ação hormonal por mudanças de receptores dentro da glândula, confirmando assim nossos achados os quais indicaram variação de tamanho das células em relação à idade. E devido aos hormônios esteroides sexuais influenciarem na expressão de receptores nos timócitos houve a observação de diferença de tamanho entre machos e fêmeas no presente trabalho.
O volume do timo deteve um aumento em conformidade com o desenvolvimento do animal, apresentando um aumento brusco no Grupo II (45 dias). Uma explicação para tal fato decorre devido ao crescimento do órgão que ao nascimento se apresenta totalmente desenvolvido. Já na literatura há relatos de que este desenvolvimento ocorre no período pré-natal (aproximadamente 60 dias) e no início de vida pós-natal.
As variações de comprimento, espessura e largura de maneira geral apresentaram aumento conforme o desenvolvimento gradual dos animais, corroborando desta maneira com o trabalho de Agreste (2005).
Sugere-se que as diferenças nos valores encontrados nas variações estereológicas podem estar relacionadas com a variação da forma macroscópica do órgão, que por sua vez pode determinar variações de dimensão deste, resultando em alterações nos valores estereológicos, porquanto em muitas equações são utilizados os elementos métricos de dimensão para os cálculos da estereologia.
CONCLUSÕES
O timo apresentou uma coloração rósea-pálida e com duas porções, a torácica e a cervical, sendo que cada uma delas possuía um lobo direito e um lobo esquerdo em sua maioria. A porção torácica localizava-se em região de mediastino cranial, entre os pulmões e à base do coração. E a porção cervical estendia-se além das costelas em sentido cranial, estando localizada ventralmente à traquéia.
A estrutura celular do timo demonstrou-se organizada com a presença de agregados concêntricos, os chamados corpúsculos tímicos, formados por células epiteliais mortas, sustentada por uma cápsula de tecido conjuntivo de onde partiam septos que ao penetrar no órgão dividiam-o em lóbulos.
Ocorreram variações significativas quanto à lobação e as dimensões do timo entre indivíduos da mesma faixa etária, e entre sexos diferentes. Os valores relativos ao comprimento, espessura e largura, de maneira geral, apresentaram aumento, em conformidade ao desenvolvimento dos animais, mas com diferenças entre os sexos.
Agradecimentos.- À Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), pelo apoio financeiro recebido (2010/52480-8) e pela bolsa de doutorado de Camila E. Barroso (2008/58096-5).
Recebido em 30 de agosto de 2012.
Aceito para publicação em 8 de outubro de 2012.
- Agreste F.R. 2005. Quantificação da vascularização do timo em cães. Dissertação de Mestrado em Anatomia dos Animais Domésticos e Silvestres, Faculdade de Medicina Veterinária e Zootecnia, USP, São Paulo, SP. 104p.
- Andersen B.B. & Pakkenberg B. 2003. Stereological quantitation in cerebella from people with schizopherenia. Braz. J. Psycol. 182:354-361.
- Appolinário A.V.M. 1998. Vascularização arterial do timo de coelhos (Oryctolagus cuniculus Linnaeus, 1758) da raça Nova Zelândia Branco. Dissertação de Mestrado em Anatomia dos Animais Domésticos e Silvestres, Faculdade de Medicina Veterinária e Zootecnia, USP, São Paulo, SP. 66p.
- Banks W.J. 1992. Sistema linfático e imunitário, p.370-390. In: Banks W.J. (Ed.), Histologia Veterinária Aplicada. 2Ş ed. Manole, São Paulo. 658p.
- Bar-Dayan Y., Akek A., Goldberg I. & Kopolovic J. 1999. Proliferation, apoptosis and thymic involution. Tissue Cell 31(4):391-396.
- Bockman D.E. 1997. Development of the thymus. Microsc. Res. Tech. 38:209-215.
- Bombonato P.P. 1997. Aspectos da morfologia, topografia e vascularização arterial do timo em fetos de búfalo. Tese de Livre Docência em Anatomia dos Animais Domésticos e Silvestres, Faculdade de Medicina Veterinária e Zootecnia, USP, São Paulo, SP. 88p.
- Daschinger V.E.K. 1983. Zur Topographie und Blutgefäßversorgung des Thymus der neugeborenen Hauskatze (Felis silvestris f. catus). Zeitschr. Vers. Tierkde 25:100-113.
- Drumond S.S. 1996. Aspectos morfométricos e vascularização arterial do timo em suínos da raça Hampshire. Tese de Doutorado em Anatomia dos Animais Domésticos e Silvestres, Faculdade de Medicina Veterinária e Zootecnia, USP, São Paulo, SP. 57p.
- George A.J.T. & Ritter M. 1996. Thymic involution with ageing: obsolence or good housekeeping? Immun. Tod. 17:267-272.
- Getty R. 1986. Sisson/Grossman Anatomia dos Animais Domésticos. Vol.2. 5Ş ed. Guanabara Koogan, Rio de Janeiro. 2000p.
- Gundersen H.J.G. & Jensen E.B. 1987. The efficiency of systematic sampling in stereology and its prediction. J. Microsc. 147:219-223.
- Hudson L.C. & Hamilton W.P. 1993. Atlas of Feline Anatomy for Veterinarians. W.B. Saunders, Philadelphia. 287p.
- Junqueira L.C. & Carneiro J. 1999. Histologia Básica. 9Ş ed. Guanabara Koogan, Rio de Janeiro. 488p.
- Kato S. 1997. Thymic microvascular system. Microsc. Res. Tec. 38(3):287-299.
- König H.E. & Liebich H.G. 2004. Anatomia dos Animais Domésticos: texto e atlas. Vol.2. Artmed, Porto Alegre. 788p.
- Kurz B., Feindt J., Gaudecker V.B., Kranz A., Loppnow H. & Mentlein R. 1997. B-adrenoceptor-mediated effects in rat cultured thymic epithelial cells. Braz. J. Pharm. 120:1401-1408.
- Leposavić G., Plećas B. & Kosec D. 2000. Differential effects of chronic propanolol treatment on the phenotypic profile of thymocytes from immature and adult rats. Immunopharm. 46:79-87.
- Mandarim-de-Lacerda C.A. 2003. Stereological tools in biomedical research. Anais Acad. Bras. Ciênc 75(4):469-489.
- Mattfeldt T., Mall G., Gharehbaghi H. & Moller P. 1990. Estimation of surface area and length with the orientator. J. Microsc. 159:301-317.
- Marchetti B., Morale M.C. & Palletier G. 1990. Sympathetic nervous system control of the rat thymus gland maturation: autoradiographic localization of the B2-adrenergicreceptor in the thymus and presence of sexual dimorphism during ontogeny. Neur. Endocr. Immun. 3:103-114.
- Mayhew T.M. 1992. A review of recent advances in sterology for quantifying neural structure. J. Neurociênc. 21:313-328.
- Melo A.M.A. & Lage M.C.D. 1987. Alguns aspectos da estrutura e funções do timo em Gallus domesticus Repert. Trab. Lab. Nac. Invest. Vet. 19:13-24.
- Olsen N.J. & Kovacs W.J. 1996. Gonadal steroids and immunity. Endoc. Rev. 17(4):369-384.
- Savino W. & Dardenne M. 1995. Immune-neuroendocrine interactions. Immun. Today 16(7):318-322.
- Silva F.O.C., Severino R.S., Santos A.L.Q., Drummond A.S., Silva M., Bombonato P.P., Rezende R.J. & Lima E.M.M. 2001. Suprimento arterial do timo em gatos sem raça definida. Biosci. J. 17(1):61-66.
- Solarović P.B., Lalić L. & Leposavić G. 2004. Age-dependent morphometrical changes in the thymus of male propanolol-treated rats. Ann. Anat. 186:141-147.
- Weibel E.R. 1979. Stereological methods: Practical methods for biological morphometry. Academic Press Inc., London. 400p.
- Wulfsohn D., Gundersen H.J., Vedel-Jensen E.B. & Nyengaard J.R. 2004. Volume estimation from projections. J. Microsc. 215:111-120.
- Vizi E.S., Orsó E., Osipenko O.N., Haskó G. & Elenkov I.J. 1995. Neurochemical, electrophysiological evidence for a noradrenergic link between the sympathetic nervous system and thymocytes. Neurosci. 86(4):1263-1276.
Datas de Publicação
-
Publicação nesta coleção
15 Fev 2013 -
Data do Fascículo
Dez 2012
Histórico
-
Recebido
30 Ago 2012 -
Aceito
08 Out 2012