ABSTRACT:
This study was made to investigate the anatomical features of the white-eared opossum skull, by osteology and radiographic anatomy. For this, five animals were used without sexual distinction. The skull was examined by radiographic and macroscopic characteristics. The skulls were then subjected to maceration. The skull was described macroscopically according to standard views, i.e. dorsal and caudal, lateral, ventral, and midsagittal. The skull can be divided into facial (viscerocranium) and cranial (neurocranium) regions. The facial region was elongated and more developed than neurocranium. The supraorbital foramen was absent. The tympanic bulla is not well developed. The zygomatic arch was formed by zygomatic process of the temporal bone, zygomatic process of the maxilla, and temporal process of the zygomatic bone. There was no significant difference between bones found in this study when compared with those described for others mammals. These findings may contribute to the better understanding of the anatomy and biology of the white-eared opossum.
INDEX TERMS:
Anatomy; radiography; white-eared opossum; Didelphis albiventris; opossum; marsupial; skull; wild animals
RESUMO:
Este estudo objetivou investigar as características anatômicas do crânio do gambá-de-orelha-branca, através da anatomia óssea e radiográfica. Para tanto, cinco animais foram usados sem distinção sexual. O crânio foi estudado através de análises radiográficas e macroscópicas. Para esta última, os crânios foram macerados. O crânio foi descrito macroscopicamente de acordo com as vistas ou normas rotineiras, isto é, dorsal e caudal, lateral, ventral e sagital. O crânio pode ser dividido em regiões facial (viscerocrânio) e cranial (neurocrânio). A região facial se mostrou alongada e mais desenvolvida que o neurocrânio. O forame supraorbital estava ausente. A bula timpânica não era bem desenvolvida. O arco zigomático era formado pelo processo zigomático do osso temporal, processo zigomático da maxila, e pelo processo temporal do osso zigomático. Não houve diferenças significativas entre os ossos estudados quando comparados com os já descritos para outros mamíferos. Estes resultados podem contribuir com um melhor entendimento da anatomia e biologia deste animal.
TERMOS DE INDEXAÇÃO:
Anatomia; radiografia; gambá-de-orelha-branca; Didelphis albiventris; gambá; marsupial; crânio; animais silvestres
Introduction
Mammals of the genus Didelphis are found exclusively in the Americas and Australia. Two species of the genus Didelphis appear in the State of São Paulo, Brazil, Didelphis albiventris and Didelphis aurita. D. albiventris is a medium-sized marsupial species, called white-eared opossum, because can be easily identified by its white ears with a dark base. They have a black body, long tapering nose and naked tail (Emmons & Feer 1997Emmons L.H. & Feer F. 1997. Neotropical rainforest mammals: a field guide. University of Chicago Press, Chicago .).
The white-eared opossum has solitary and nocturnal habit, with an arboreal and a terrestrial behavior. It may be found in open and deciduous forest, and Cerrado, Caatinga, and Chaco environments, and also in areas of anthropogenic change, such as agricultural habitats, deforested areas and suburban environment. The white-eared opossum is very common in the urban region of many Brazilian municipalityies (Cerqueira 1985Cerqueira R. 1985. The distribution of Didelphis in South America (Polyprotodontia, Didelphidae). J. Biogeogr. 12:135-145., Cáceres 2002Cáceres N.C. 2002. Food habitas and seeds dispersal by the white-eared opossum, Didelphis albiventris, in Southern Brazil. Stud. Neotrop. Fauna Environ. 37:97-104., Alessio et al. 2005Alessio F.M., Pontes A.R.M. & Silva V.L. 2005. Feeding by Didelphis albiventris on tree gum in the Northeastern atlantic forest of Brazil. Mastoz. Neotr. 12:53-56., Smith 2007Smith P. 2007. White-eared opossum Didelphis albiventris. Fauna Paraguay Handbook of Mammals of Paraguay, 1:1-19. Acessed November 10, 2015 at < Acessed November 10, 2015 at http://www.faunaparaguay.com/mam1Didelphisalbiventris.pdf
>
http://www.faunaparaguay.com/mam1Didelph...
, Cerqueira & Tribe 2008Cerqueira R. & Tribe C. 2008. Genus Didelphis, p.17-25. In: Gardner A. (Ed.), Mammals of South America. Marsupilas, xenarthrans, shrews, and bats. University of Chicago Press, Chicago., Costa et al. 2008Costa L., Astua de Moraes D., Brito D., Soriano P., Iew D. & Delgado C. 2008. “Didelphis albiventris” (on-line). IUCN Red List of Threatened Species. Accessed May 13, 2015 at < Accessed May 13, 2015 at www.iucnredlist.org
>
www.iucnredlist.org...
, Siciliano Martina 2013Siciliano Martina L. 2013. Didelphis albiventris (on-line). Animal Diversity web. Acessed November 10, 2015 at < Acessed November 10, 2015 at http://animaldiversity.org/accounts/Didelphis_albiventris
/>
http://animaldiversity.org/accounts/Dide...
).
The mammalian skull study is important because the phenotic appearance of the head of animal species depends on the shape of the skull (Kunzel et al. 2003Kunzel W., Breit S. & Oppel M. 2003. Morphometric investigations of breed specific features in feline skulls and considerations on their functional implications. Anat. Histol. Embryol. 32:218-223., Yahaya et al. 2013Yahaya A., Olopade J.O. & Kwari H.D. 2013. Morphological analysis and osteometry of the foramen magnum of the one-humped camel (Camelus dromedarius). Anat. Histol. Embryol. 42:155-159.). Therefore, morphological studies of the skull are important as they contribute to anatomical knowledge (Olopade & Onwuka 2009Olopade J.O. & Onwuka S.K. 2009. Morphometric analysis of the skull of the Sahel goat breed: basic and clinical anatomy. Ital. J. Anat. Embryol. 114:167-178.); the skull morphology has also been used as major skeletal structure to determine taxonomic affiliations (Bruenner et al. 2002Bruenner H., Lugon-Moulin N., Balloux F., Fumagalli L. & Hausser J. 2002. A taxonomical re-evaluation of the Valais chromosome race of the common shrew Sorex araneus (Insectivora: Soricidae). Acta Theriol. 47:245-275., Yahaya et al. 2013Yahaya A., Olopade J.O. & Kwari H.D. 2013. Morphological analysis and osteometry of the foramen magnum of the one-humped camel (Camelus dromedarius). Anat. Histol. Embryol. 42:155-159.).
The genus Didelphis has been used as a model to understand various aspects of evolution of the mammalian skull (Crompton 1989Crompton A.W. 1989. The evolution of mammalian mastication, p.23-40. In: Wake D.B. & Roth J. (Eds), Complex Organismal Function: integration and evolution in vertebrates. John Wiley and Sons Ltd, New York., Abdalla et al. 2001Abdalla F., Flores D.A. & Giannini N.P. 2001. Postweaning ontogeny of the skull of Didlephis albiventris. J. Mammal. 82:190-200.). Several studies on the skull of small marsupials have been performed, specially dealing with allometry and craniometry. Postnatal development of the cranial structure in Didelphis albiventris has been examined by analysis of allometry (Abdalla et al. 2001Abdalla F., Flores D.A. & Giannini N.P. 2001. Postweaning ontogeny of the skull of Didlephis albiventris. J. Mammal. 82:190-200.). On the basis of morphologic and morphometric data of the skull, phenotypic variation in white-eared opossum and its relevance for the taxonomy was analyzed (Lemos & Cerqueira 2002Lemos B. & Cerqueira R. 2002. Morphological differentiation in the white-eared opossum group (Didelphidae, Didelphis). J. Mammal. 83:354-369.). Didelphis has been reviewed systematically by craniometrical analysis of skulls from species of this genus in Venezuela (Ventura et al. 2002Ventura J., Salazar M., Pérez-Hernández R. & López-Fuster M.J. 2002. Morphometrics of the genus Didelphis (Didelphimorpha: Didelphidae) in Venezuela. J. Mammal. 83:1087-1096.). These authors claimed, based on morphometric differences, that Didelphis imperfect is an another specie of the genus Didelphis and is a relatively small-sized Didelphis that lives in the evergreen rainforest of tropical, subtropical, and temperate levels in Venezuela. No description of the gross anatomy and radiography was found in the available literature. Thus, the aim of this study was to describe the gross anatomy and radiographic features of the skull of the adult white-eared opossum inhabiting the Brazilian southeastern region. These findings may contribute to better understanding of the anatomy and biology of the white-eared opossum.
Materials and Methods
This study followed the guidelines for the care and use of laboratory animals and was approved by the Animal Use and Ethics Committee (626/2014) and by the National Environmental and Wildlife Bureau (SISBIO 44768-2).
The skulls of five adult white-eared opossum (Didelphis albiventris) were used in this study. The opossums had died for reasons unrelated to this study. All animals had originated from Center of Medicine and Research in Wildlife (CEMPAS), School of Veterinary Medicine and Animal Science, Unesp, Botucatu/SP, Brazil.
The specimens were collected immediately after the animals had died and stored at -20°C. For radiographic examinations the animals were defrosted, and the images were acquired in dorsoventral, and mediolateral views. A digital radiographic system was used (EZy-Rad Pro X-ray system, Shimadzu; Console Advance, DR-ID 300CL, Fujifilm) with focus-film distance of 100cm and an exposure of 70kV, 200mA, 6.4mAs.
Afterwards, all soft tissues were removed from the skulls by boiling, detaching, and rinsing, so that a clear view of the fine bony structures was achieved. The skulls were left to dry at room temperature before further observations according to He et al. (2002)He T., Friede H. & Kiliaridis S. 2002. Macroscopic and roentgenographic anatomy of the skull of the ferret (Mustela putoris furo). Lab. Anim. 36:86-96.. The bones were identified, described, photodocumented in standard views, i.e. dorsal and caudal, lateral, ventral, and midsagittal. Therefore, the structures studied were named according to Nomina Anatomica Veterinaria (International Committee on Veterinary Gross Anatomical Nomenclature, 2012International Committee on Veterinary Gross Anatomical Nomenclature 2012. Nomina Anatomica Veterinária. 5th ed. World Association on Veterinary Anatomist, Knoxville. 177p.).
Results
The skull of white-eared opossum can be divided into a facial plus palatal region (viscerocranium) and a cranial portion (neurocranium). The facial region of the adult opossum was elongated and more developed than the neurocranium.
Dorsal and caudal view of skull
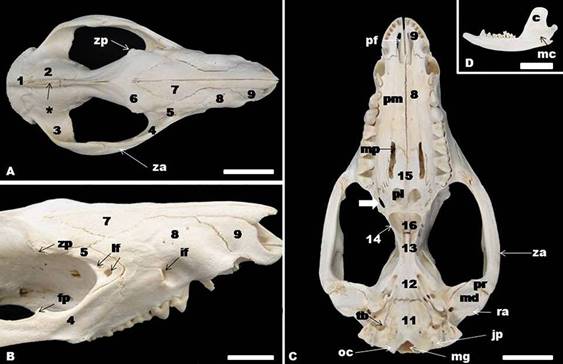
Caudally, the cranial region comprises the interparietal, parietal, frontal, temporal, and part of the sphenoid bones. This part of the sphenoid bone can be called alisphenoid, because consists in the greater wing of the sphenoid bone. The external sagittal crest was the most prominent marking of the dorsum of the cranium. It was a median longitudinal ridge, formed by the interparietal, parietal, and frontal bones (Fig.1A,3A-D). Rostrally, the crest tends to split to form a “V”, which normally extends to the zygomatic processes of frontal bones on each side of the skull (Fig.1A). The nuchal crest marks the boundary between the dorsal and caudal surface of the skull (Fig.3C). The facial region comprises the dorsal surfaces of the nasal, maxilla, incisive, lacrimal, and zygomatic bones. The incisive and nasal bones form the border of the external nasal opening. The zygomatic arch could be seen in this view (Fig.1A-C). The caudal aspect of the white-eared opossum skull was formed by the interparietal and occipital bones, squamous and petrosus parts of the temporal bone (Fig.3C-D).

The white-eared opossum skull. Interparietal (1), parietal (2), temporal (3), zygomatic (4), lacrimal (5), frontal (6), nasal (7), maxilla (8), incisive (9), basioccipital (11), basisphenoid (12), presphenoid (13), pterygoid (14), palatine (15), vomer bones (16). Note also zygomatic process of the frontal bone (zp), zygomatic arch (za), lacrimal foramen (lf), infraorbital foramen (if), frontal process of the zygomatic bone (fp), magnum foramen (mg), occipital condyle (oc), jugular process (jp), tympanic bulla (tb), mandibular fossa (md), retroarticular process (ra), prearticular process (pr), maxillopalatine fenestra (mp), palatine process of the maxilla (pm), palatine fenestra (pl), palatine fissure (pf), mandibular canal (mc), coronoid process of the mandible (c), external sagittal crest (*). Bar=2cm.

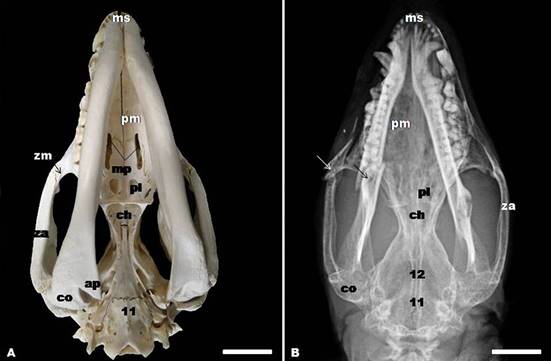
(A) Gross anatomy and (B) dorsoventral radiographic view of the skull showing basioccipital (11), and basisphenoid bones (12), zygomatic arch (za), zygomatic process of the maxilla (zm), angular process (a), condylar process (co), choana (ch), palatine fenestra (pl), maxillopalatine fenestra (mp), palatine process of the maxilla (pm), and mandibular symphysis (ms). The arrows indicated bone fracture probably caused by animal bite. Bar=2cm.
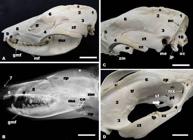
(A-D) Gross anatomy, and (B) mediolateral radiography of the skull. Interparietal (1), parietal (2), temporal (3), zygomatic (4), lacrimal (5), frontal (6), nasal (7), maxilla (8), incisive (9), and occipital bones (10). Showing also external sagittal crest (*), infraorbital foramen (if), lacrimal foramina (lf), zygomatic process of the temporal bone (zt), masseteric fossa (ma), mental foramina (mf), greater mental foramen (gmf), zygomatic process of the frontal bone (zp), external acoustic meatus (me), angular process (ap), condylar process (co), zygomatic process of the maxilla (zm), temporal process of the zygomatic bone (zz), nuchal crest (nc), jugular process (jp), occipital condyle (oc), temporal fossa (tf), frontal process of the zygomatic bone (fp), maxillar foramen (mx), sphenopallatine foramen (sf), caudal palatine foramen (thick arrow). Bar=2cm.
Lateral view of the skull
In this view was evident the temporal, interparietal, parietal, frontal, lacrimal, sphenoid, zygomatic, maxilla, nasal, and incisive bones. The zygomatic arch is best seen in this aspect; it was formed by the zygomatic process of the temporal bone, the zygomatic process of the maxilla, and the temporal process of the zygomatic bone. Therefore, it was observed that the orbit, the frontal process of the zygomatic bone, and the temporal and pterygopalatine fossae. Rostrally, in the pterygopalatine fossa could be noted a rostral group of foramina which comprises the maxillary, sphenopalatine, and caudal palatine foramina (Fig.3D). The infraorbital foramen and external acoustic meatus also were found in this view of the skull. The lacrimal bone formed the anterior part of the orbit and was perforated by two lacrimal foramina on each side. The zygomatic process of the frontal bone was present (Fig.3A-D).
The mandible consists of a pair of bones that articulate rostrally at the mandibular symphysis. Each of part comprises an elongated horizontal body that contains the dental alveoli, an ascending ramus with well-developed coronoid and condylar processes, and a posteroventral angular process. A greater and rostral mental foramen and two small mental foramina, a masseteric fossa, condylar process, coronoid process, mandible notch, and an angular process were found laterally in the mandible. Medially, a great mandibular canal was found.. The dental formula of the opossum is incisors 5/4, canines 1/1, premolars 3/3, and molars 4/4 (50 in total) (Fig.1D, 2A-B, 3A).
Ventral view of skull
In the ventral view, three regions of the skull were distinct: the base of the cranium, the choanal region, and the hard palate. The tympanic bulla could be seen in this aspect. They were two undeveloped protuberances lying on the two sides of the ventrocaudal part of the cranium. The magnum foramen, occipital condyle, and jugular process were observed caudal to the tympanic bulla, in the occipital bone. Medial and rostrally to the tympanic bulla, from caudal to rostral, appeared basioccipital, basisphenoid, presphenoid, pterygoid and vomer bones. The basioccipital bone articulates laterally with the tympanic and petrous parts of the temporal bone and rostrally with the body of the basisphenoid. The carotic canal, alar canal with rostral and caudal alar foramina, stylomastoid foramen, and minor palatine foramen were presented. In the caudal portion of the zygomatic arch, could be observed retroarticular process, mandibular fossa, and prearticular process (Fig.1C, 4A-B).

(A) ventral, (B) lateral, and (C-D) midsagittal aspects of the opossum skull. (2) Parietal (2), (6) frontal (6), (7) nasal (7), (10) occipital (10), (11) basicoccipital (11), and (12) basisphenoid bones (12). Note also magnum foramen (mg), occipital condyle (oc), jugular process (jp), estylomastoid foramen (es), external acoustic meatus (me), carotic canal (cc), caudal alar foramen (c1), rostral alar foramen (c2), transverse canal foramen (tc), spinous process (sp), perpendicular plate of the ethmoid (pp), orbital fissure (fo), internal acoustic meatus (im), hypoglossal canal (hc), rostral cranial fossa (F1), middle cranial fossa (F2), caudal cranial fossa (F3), cribriform plate of the ethmoid (cp), round foramen (rf), oval foramen (of), jugular foramen (jf), esphenopallatine foramen (ef), caudal palatine foramen (thick arrow), conchal crest (arrow head), ethmoidal crest (arrow), and ethmoturbines (et). Bar=2cm.
Palatine, maxilla, and incisive bones formed the facial region of the ventral aspect of the skull. The hard palate comprised the ventral surface of this part and was constituted by the palatine process of the incisive bone, palatine process of the maxilla and palatine bone. The choanae were open at the end of the hard palate. In the incisive ventral part were two palatine fissures separated by the palatine processes of the incisive. Therefore, the maxillopalatine fenestra appeared in the maxilla and palatine bones, and a palatine fenestra and a posterolateral palatal foramen was noted in the palatine bones (Fig.1C, 2A-B).
Midsagittal view of skull
In the midsagittal view of the opossum skull cranial and nasal cavities can be seen. The cranial cavity is caudal, and the base of the cavity is formed by the occipital and sphenoid bones. The caudal wall is formed by the occipital, and the rostral wall is formed by the cribriform plate of the ethmoid. The temporal, parietal, frontal, sphenoid, and occipital bones formed the lateral wall. This cavity comprises three compartments or fossae. The rostral cranial fossa is located caudallly to the cribriform plate and houses the olfactory bulbs and tracts, and the remaining parts of the frontal lobes of the brains. The middle cranial fossa is situated at a lower level than the rostral cranial fossa. This fossa supports the cerebrum, diencephalon and mesencephalon. The digital impressions that are formed by the gyri and sulci of the brains, were not observed in the internal face of this fossa The caudal cranial fossa is located caudally to the middle cranial fossa. The pons, the cerebellum, and the medulla oblongata rest on it. The orbital fissure, round foramen, oval foramen, carotic canal, hypoglossal canal and jugular foramen were observed in this cavity (Fig.4C-D).
The nasal cavity is separated from the cranial cavity by the cribriform plate and is composed of two symmetric halves which are separated from each other by the nasal septum. Each half presents ventral nasal conchae ventrorostrally and the ethmoturbinates dorsocaudally. The nasal septum is formed caudally by the perpendicular plate of the ethmoid and by the vomer. Rostrally, this septum consists of septal cartilage, what was not observed in the macerated skulls. The ventral and dorsal nasal conchae protrude medially from the conchal crest and from the ethmoidal crest, respectively (Figs.4C-D).
Discussion
The available literature suggests that research on the marsupial skull has been developed mainly in regard to skull allometry. The ontogeny of the skull of Didelphis albiventris was studied by Abdalla et al. (2001)Abdalla F., Flores D.A. & Giannini N.P. 2001. Postweaning ontogeny of the skull of Didlephis albiventris. J. Mammal. 82:190-200.. There is no report about anatomical features found in D. albiventris. Thus, the goal of this paper is the description of anatomic features of the South-American opossum skull.
The white-eared opossum skull was divided into the facial and cranial region. The largest part comprises the elongated facial region, and the cranial region, the brain case is relatively short. It is different from what observed for the ferret skull, which showed an elongated flat skull with a shortened facial region (He et al. 2002He T., Friede H. & Kiliaridis S. 2002. Macroscopic and roentgenographic anatomy of the skull of the ferret (Mustela putoris furo). Lab. Anim. 36:86-96., O’Malley 2005O’Malley B. 2005. Clinical anatomy and physiology of exotic species. Elsevier Saunders, St Louis.). There was no significant difference between bones found in the white-eared opossum skull when compared with those described for other mammals, as of porcupine (Yilmaz 1998Yilmaz S. 1998. Macro-anatomical investigation on the skeleton of porcupine (Hystrix cristata). III. Skeleton axiale. Anat. Histol. Embryol. 27:293-296.), ferret (He et al. 2002Emmons L.H. & Feer F. 1997. Neotropical rainforest mammals: a field guide. University of Chicago Press, Chicago .), and dog (Dyce et al. 2010Dyce K.M., Sack W.O. & Wensing C.J.G. 2010. Textbook of Veterinary Anatomy. 4th ed. Elsevier Saunders, Saint Louis, p.59-61.). Voss & Jansa (2009Voss R.S. & Jansa S.A. 2009. Phylogenetic relationships and classification of Didelphid marsupials, an extant radiation of New World Metatherium mammals. B. Amer. Mus. Nat. Hist. 322:1-177.) reported presence of the jugal bone in the marsupial Marmosa murina. However, in this study we have not identified the jugal and mastoid bones. In the cranium of the common European mole, the lacrimal, jugal, and mastoid bones were identified only in the prenatal specimens. In this animal, the jugal and lacrimal bones fuse with the maxilla during prenatal growth (Goswami & Prochel 2007Goswami A. & Prochel J. 2007. Ontogenetic morphology and allometry of the cranium in the common European mole (Talpa europaea). J. Mammal. 88:667-677.). All the skulls analyzed in this study were from adult animals and, perhaps these bones could have fused early, therefore in the Nomina Anatomia Veterinaria (NAV 2012) is not included jugal and mastoid bone.
The external sagittal crest is the most prominent structure in the dorsal view of the skull, such as described in the European badger (Meles meles) (Dinc 2001Dinc G. 2001. Porsuk (Meles meles) iskeltsistemi üzerinde makroanatomik arastirmalr III. Skeleton axiale. F. Ü. Saglik Bil. Derg. 15:175-178.) and ferret (He et al. 2002He T., Friede H. & Kiliaridis S. 2002. Macroscopic and roentgenographic anatomy of the skull of the ferret (Mustela putoris furo). Lab. Anim. 36:86-96.). The supraorbital foramen was absent in the white-eared opossum. In the raccoon, badger, otter, and marten skull, the supraorbital foramen was also absent (Yilmaz et al. 2000Yilmaz S., Dinc G. & Toprak B. 2000. Macro-anatomical investigations on skeletons of otter (Lutra lutra). III. Skeleton axial. Vet. Arhiv 70:191-198., Dinc 2001Dinc G. 2001. Porsuk (Meles meles) iskeltsistemi üzerinde makroanatomik arastirmalr III. Skeleton axiale. F. Ü. Saglik Bil. Derg. 15:175-178., Karan et al. 2006Karan M., Timurkaan S., Özdemir D. & Unsaldi E. 2006. Comparative macroanatomical study of the neurocranium in some carnivore. Anat. Histol. Embryol. 35:53-55.).
The orbit and zygomatic arch are most developed structures in the lateral view. In the macerated skull, the orbit walls are very incomplete as in ferret (He et al. 2002He T., Friede H. & Kiliaridis S. 2002. Macroscopic and roentgenographic anatomy of the skull of the ferret (Mustela putoris furo). Lab. Anim. 36:86-96.) and the dog (Dyce et al. 2010Dyce K.M., Sack W.O. & Wensing C.J.G. 2010. Textbook of Veterinary Anatomy. 4th ed. Elsevier Saunders, Saint Louis, p.59-61.). According to these authors, the orbital rim is completed by a ligament that connects the zygomatic process of the frontal bone to the frontal process of the zygomatic bone. The zygomatic arch is formed by the zygomatic process of the temporal bone, the zygomatic process of the maxilla, and the temporal process of the zygomatic bone. Dyce et al. (2010)Dyce K.M., Sack W.O. & Wensing C.J.G. 2010. Textbook of Veterinary Anatomy. 4th ed. Elsevier Saunders, Saint Louis, p.59-61. postulated that the zygomatic arch in the dog is formed by two bones, the squamous temporal and zygomatic bone. The zygomatic arch found in the ferret skull bends from zygomatic process of the maxilla to the zygomatic process of the temporal bone (He et al. 2002He T., Friede H. & Kiliaridis S. 2002. Macroscopic and roentgenographic anatomy of the skull of the ferret (Mustela putoris furo). Lab. Anim. 36:86-96.).
The tympanic bulla is not well developed in Didelphis albiventris, unlike well developed in porcupine (Yilmaz 1998Yilmaz S. 1998. Macro-anatomical investigation on the skeleton of porcupine (Hystrix cristata). III. Skeleton axiale. Anat. Histol. Embryol. 27:293-296.), ferret (He et al. 2002He T., Friede H. & Kiliaridis S. 2002. Macroscopic and roentgenographic anatomy of the skull of the ferret (Mustela putoris furo). Lab. Anim. 36:86-96.), badger, otter, marten, cat, and dog (Karan et al. 2006Karan M., Timurkaan S., Özdemir D. & Unsaldi E. 2006. Comparative macroanatomical study of the neurocranium in some carnivore. Anat. Histol. Embryol. 35:53-55., Dyce et al. 2010Dyce K.M., Sack W.O. & Wensing C.J.G. 2010. Textbook of Veterinary Anatomy. 4th ed. Elsevier Saunders, Saint Louis, p.59-61.). There is no difference between bone formation found in the hard palate of the white-eared opossum skull and in the dog (Dyce et al. 2010Dyce K.M., Sack W.O. & Wensing C.J.G. 2010. Textbook of Veterinary Anatomy. 4th ed. Elsevier Saunders, Saint Louis, p.59-61.). The hard palate is perforated by foramina and fenestrae. According to Voss & Jansa (2009)Voss R.S. & Jansa S.A. 2009. Phylogenetic relationships and classification of Didelphid marsupials, an extant radiation of New World Metatherium mammals. B. Amer. Mus. Nat. Hist. 322:1-177., these foramina and fenestrae exhibit considerable variation in occurrence, size, and position among marsupials. The maxillopalatine fenestra may also be called major palatine foramen (Wible 2003Wible J.R. 2003. On the cranial osteology of the short-tailed opossum Monodelphis brevicaudata (Didelphidae, Marsupialia). Ann. Carnegie Mus. 72:137-142.), and the other name of the posterolateral palatal foramen could be palatine canal or minor palatine foramen (Wible 2003Smith P. 2007. White-eared opossum Didelphis albiventris. Fauna Paraguay Handbook of Mammals of Paraguay, 1:1-19. Acessed November 10, 2015 at < Acessed November 10, 2015 at http://www.faunaparaguay.com/mam1Didelphisalbiventris.pdf
>
http://www.faunaparaguay.com/mam1Didelph...
). Archer (1976)Archer M. 1976. The basicranial region of marsupicarnivores (Marsupialia), interrelationships of carnivorous marsupials, and affinities of the insectivorous marsupial paramelids. Zool. J. Linn. Soc. Lond. 59:217-322. reported that this foramen transmits the minor palatine artery from the maxillary artery to the ventral surface of the palate. The mandible comprises two parts that are firmly united by the connective tissue of the mandibular symphysis, similar to the ferret (He et al. 2002He T., Friede H. & Kiliaridis S. 2002. Macroscopic and roentgenographic anatomy of the skull of the ferret (Mustela putoris furo). Lab. Anim. 36:86-96.). In our study, the mandible is the largest bone in the skull. The same was described for the guinea pig skull (O’Malley 2005O’Malley B. 2005. Clinical anatomy and physiology of exotic species. Elsevier Saunders, St Louis.).
Knowledge of the skull morphology in mammals is important, because provides baseline anatomic information and may collaborate to veterinarian medical and surgical clinics. In this study, clinically relevant anatomic structures were identified and labeled in the white-eared opossum skull and many of these features are easily palpated in life.
References
- Abdalla F., Flores D.A. & Giannini N.P. 2001. Postweaning ontogeny of the skull of Didlephis albiventris J. Mammal. 82:190-200.
- Alessio F.M., Pontes A.R.M. & Silva V.L. 2005. Feeding by Didelphis albiventris on tree gum in the Northeastern atlantic forest of Brazil. Mastoz. Neotr. 12:53-56.
- Archer M. 1976. The basicranial region of marsupicarnivores (Marsupialia), interrelationships of carnivorous marsupials, and affinities of the insectivorous marsupial paramelids. Zool. J. Linn. Soc. Lond. 59:217-322.
- Bruenner H., Lugon-Moulin N., Balloux F., Fumagalli L. & Hausser J. 2002. A taxonomical re-evaluation of the Valais chromosome race of the common shrew Sorex araneus (Insectivora: Soricidae). Acta Theriol. 47:245-275.
- Cáceres N.C. 2002. Food habitas and seeds dispersal by the white-eared opossum, Didelphis albiventris, in Southern Brazil. Stud. Neotrop. Fauna Environ. 37:97-104.
- Cerqueira R. 1985. The distribution of Didelphis in South America (Polyprotodontia, Didelphidae). J. Biogeogr. 12:135-145.
- Cerqueira R. & Tribe C. 2008. Genus Didelphis, p.17-25. In: Gardner A. (Ed.), Mammals of South America. Marsupilas, xenarthrans, shrews, and bats. University of Chicago Press, Chicago.
- Costa L., Astua de Moraes D., Brito D., Soriano P., Iew D. & Delgado C. 2008. “Didelphis albiventris” (on-line). IUCN Red List of Threatened Species. Accessed May 13, 2015 at < Accessed May 13, 2015 at www.iucnredlist.org >
» www.iucnredlist.org - Crompton A.W. 1989. The evolution of mammalian mastication, p.23-40. In: Wake D.B. & Roth J. (Eds), Complex Organismal Function: integration and evolution in vertebrates. John Wiley and Sons Ltd, New York.
- Dyce K.M., Sack W.O. & Wensing C.J.G. 2010. Textbook of Veterinary Anatomy. 4th ed. Elsevier Saunders, Saint Louis, p.59-61.
- Dinc G. 2001. Porsuk (Meles meles) iskeltsistemi üzerinde makroanatomik arastirmalr III. Skeleton axiale. F. Ü. Saglik Bil. Derg. 15:175-178.
- Emmons L.H. & Feer F. 1997. Neotropical rainforest mammals: a field guide. University of Chicago Press, Chicago .
- Goswami A. & Prochel J. 2007. Ontogenetic morphology and allometry of the cranium in the common European mole (Talpa europaea). J. Mammal. 88:667-677.
- He T., Friede H. & Kiliaridis S. 2002. Macroscopic and roentgenographic anatomy of the skull of the ferret (Mustela putoris furo). Lab. Anim. 36:86-96.
- International Committee on Veterinary Gross Anatomical Nomenclature 2012. Nomina Anatomica Veterinária. 5th ed. World Association on Veterinary Anatomist, Knoxville. 177p.
- Karan M., Timurkaan S., Özdemir D. & Unsaldi E. 2006. Comparative macroanatomical study of the neurocranium in some carnivore. Anat. Histol. Embryol. 35:53-55.
- Kunzel W., Breit S. & Oppel M. 2003. Morphometric investigations of breed specific features in feline skulls and considerations on their functional implications. Anat. Histol. Embryol. 32:218-223.
- Lemos B. & Cerqueira R. 2002. Morphological differentiation in the white-eared opossum group (Didelphidae, Didelphis). J. Mammal. 83:354-369.
- Olopade J.O. & Onwuka S.K. 2009. Morphometric analysis of the skull of the Sahel goat breed: basic and clinical anatomy. Ital. J. Anat. Embryol. 114:167-178.
- O’Malley B. 2005. Clinical anatomy and physiology of exotic species. Elsevier Saunders, St Louis.
- Siciliano Martina L. 2013. Didelphis albiventris (on-line). Animal Diversity web. Acessed November 10, 2015 at < Acessed November 10, 2015 at http://animaldiversity.org/accounts/Didelphis_albiventris />
» http://animaldiversity.org/accounts/Didelphis_albiventris - Smith P. 2007. White-eared opossum Didelphis albiventris Fauna Paraguay Handbook of Mammals of Paraguay, 1:1-19. Acessed November 10, 2015 at < Acessed November 10, 2015 at http://www.faunaparaguay.com/mam1Didelphisalbiventris.pdf >
» http://www.faunaparaguay.com/mam1Didelphisalbiventris.pdf - Ventura J., Salazar M., Pérez-Hernández R. & López-Fuster M.J. 2002. Morphometrics of the genus Didelphis (Didelphimorpha: Didelphidae) in Venezuela. J. Mammal. 83:1087-1096.
- Voss R.S. & Jansa S.A. 2009. Phylogenetic relationships and classification of Didelphid marsupials, an extant radiation of New World Metatherium mammals. B. Amer. Mus. Nat. Hist. 322:1-177.
- Wible J.R. 2003. On the cranial osteology of the short-tailed opossum Monodelphis brevicaudata (Didelphidae, Marsupialia). Ann. Carnegie Mus. 72:137-142.
- Yahaya A., Olopade J.O. & Kwari H.D. 2013. Morphological analysis and osteometry of the foramen magnum of the one-humped camel (Camelus dromedarius). Anat. Histol. Embryol. 42:155-159.
- Yilmaz S. 1998. Macro-anatomical investigation on the skeleton of porcupine (Hystrix cristata). III. Skeleton axiale. Anat. Histol. Embryol. 27:293-296.
- Yilmaz S., Dinc G. & Toprak B. 2000. Macro-anatomical investigations on skeletons of otter (Lutra lutra). III. Skeleton axial. Vet. Arhiv 70:191-198.
Publication Dates
-
Publication in this collection
Nov 2016
History
-
Received
20 Jan 2016 -
Accepted
14 July 2016