
ABSTRACT:
The goal of this work was to characterize morphometrically diaspores from three species of mistletoes (Passovia pyrifolia, Struthanthus marginatus e Phoradendron mucronatum), as well as to study its germination behavior according to temperature and luminosity. In germination tests, the completely randomized design was used, in factor scheme 4 x 2, with 4 replications of 15 diaspores. The factors for the first test were different temperatures and two diaspore conditions (intact and without epicarp). In the second test, factors were different light qualities and two diaspore conditions. The analyzed variables were: germination percentage, germination rate index (GRI) and mean germination time (MGT). Mistletoe’s diaspores present an elliptic shape with viscin layer, an essential characteristic for the dispersion of the species. In addition to this, they present a distinction as for the moisture content when compared to the Loranthaceae and Viscaceae, where the latter presents the highest content. Germination is greater at the alternated temperature of 20-30 oC for P. pyrifolia and P. mucronatum diaspores and 20 oC for S. marginatus. Luminosity interfered in the germination of P. mucronatum and S. marginatus but did not influence P. pyrifolia. The diaspore condition influenced the germination of the three studied species, observing a significant increase in their germination when the epicarp was removed. Temperature and luminosity interfere directly in the GRI and MGT of the three mistletoes species.
Keywords:
mistletoes; temperature; luminosity
RESUMO:
O objetivo deste trabalho foi caracterizar morfometricamente diásporos de três espécies de ervas-de-passarinho (Passovia pyrifolia, Struthanthus marginatus e Phoradendron mucronatum), bem como estudar o seu comportamento germinativo em função de temperatura e luminosidade. Nos ensaios de germinação utilizou-se o delineamento inteiramente casualizado em esquema fatorial 4 x 2, com 4 repetições de 15 diásporos. Os fatores para o primeiro ensaio foram diferentes temperaturas e duas condições do diásporo (intacto e sem epicarpo). No segundo ensaio, os fatores foram diferentes qualidades de luz e duas condições do diásporo. As variáveis analisadas foram: porcentagem de germinação, índice de velocidade de germinação (IVG) e tempo médio de germinação (TMG). Os diásporos de ervas-de-passarinho apresentam formato elíptico com camada de viscina, característica essencial na dispersão das espécies. Além disso, apresentam distinção quanto ao grau de umidade quando se comparam as famílias Loranthaceae e Viscaceae, sendo esta última a que apresenta o maior teor. A germinação é maior à temperatura alternada de 20-30 ºC para diásporos de P. pyrifolia e P. mucronatum e 20 ºC para S. marginatus. A luminosidade interferiu na germinação de P. mucronatum e S. marginatus e não influenciou na de P. pyrifolia. A condição do diásporo influenciou a germinação das três espécies em estudo, observando-se incremento significativo na sua germinação quando houve a remoção do epicarpo. Temperatura e luminosidade interferem diretamente no IVG e no TMG das três espécies de ervas-de-passarinho.
Palavras-chave:
erva-de-passarinho; temperatura; luminosidade
INTRODUCTION
The species popularly known as mistletoes are hemiparasitic plants that grow on tree branches or host shrubs, establishing a connection (via haustorium) with the xylem, in order to get water and nutrients (Okubamichael et al., 2014Okubamichael D.Y. et al. Reciprocal transplant experiment suggests host specificity of the mistletoe Agelanthus natalitius in South Africa. J Trop Ecol. 2014;30:153-63.). In Portuguese, they are called ervas-de-passarinho (bird’s herb grass) due the dispersion of their seeds, since their fruits feed different species of birds which, when defecating, take the seeds to long distances (Cazzeta and Galletti, 2007).
Similarly to what happens with most plants, establishing is the key for limitation or success in the life cycle of mistletoes, where a proper disperser is required, in the presence of a proper host (Okubamichael et al., 2016Okubamichael D.Y. et al. Host specificity in parasitic plants - perspectives from mistletoes. AoB PLANTS. 2016; Sept 22:1-39). Unlike most terrestrial species, mistletoes diaspores cannot simply fall on the ground, germinate and grow.
The relation between mistletoes and birds, its primary disperser, has been widely studied (Soto-Gamboa and Bozinovic, 2002Soto-Gamboa M., Francisco Bozinovic F. Fruit-disperser interaction in a mistletoe-bird system: a comparison of two mechanisms of fruits processing on seed germination. Plant Ecol. 2002;159:171-4.). Birds generally remove some portion of the pericarp, in order to ingest the fruit and eliminate the seed, whose viscin layer, due to its viscosity characteristic, grants it the possibility to adhere to a branch. Even if seeds from many plant parasites germinate only in response to chemical signals from their hosts (Musselman et al., 2001Musselman L.J. et al. Parasitic plants major problem of food crops. Science. 2001;293:1434.), mistletoe’s seeds germinate in almost all situations (Lamont, 1985Lamont B. Host distribution, potassium content, water relations and control of two co-occurring mistletoe species. J R SocWest Aust. 1985; 68:21-5 ). However, narrower branches help the establishment (development of the haustorial connection) and the quick growth of plants (Norton et al., 1998Norton D.A. et al. Mistletoes as parasites: host specificity and speciation. Trends Ecol Evol. 1998;13:101-5.). In addition to this, some parts of a host tree are more proper for the establishment than others, reflecting differences in the dispersers’ behavior (Norton et al., 1998).
Some mistletoes species have economic importance, due to the damages they cause to plantations (Cazzetta and Galletti, 2003Cazzetta E., Galletti M. Frugivoria e especificidade por hospedeiro na erva-de-passarinho Phoradendron rubrun (L). Griseb (Viscaceae). Rev Bras Bot. 2007;30:321-50.). They may affect negatively the physiological, reproductive and ecological processes of their hosts (Press et al., 1999Press M.C. et al. Parasitic plants: physiological e ecological interactions whit their hosts. In: Press M.C. et al., editors Physiological plant ecology. Oxford: Blackwell Scientific, 1999. p.175-97.), interfering in the balance between water and nutrients, reducing photosynthesis and respiration rates and, in some situations, leading them to death (Aukema, 2003Aukema J.E. Vectors, Viscin, and Viscaceae: mistletoes as parasites, mutualists, and resources. Front Ecol Environ. 2003;1:212-9.). Its presence damages trees and shrubs in forests, orchards and gardens. Among the effects caused by these hemiparasites on host trees, it is possible to highlight the reduction of vigor and fruit and seed production, bad functioning of woody tissues, gall production, sparse foliage, apex death, predisposition to the attack of diseases and insects and even premature death (Harris, 1992Harris R.W. Arboriculture: integrated management of landscape trees, shrubs and vines. New Jersey: Prentice-Hall, 1992. 674p.).
In spite of the knowledge about damages, and infestation, little is known about the ecology of the germination of mistletoe’s diaspores. In this context, the goal of this work was to characterize morphometrically diaspores from three species of mistletoes (Passovia pyrifolia (Kunth) Tiegh., Struthanthus marginatus (Desr.) Blume and Phoradendron mucronatum (DC.) Krug & Urb., as well as to observe the germination responses to temperature and light quality variations.
MATERIAL AND METHODS
Collection and selection of the plant material
Diaspores from three mistletoes species were collected in orange lima (acidless) (Citrus sinensis - Rutaceae) and sugar-apple (Annona squamosa L. - Annonaceae) orchards, located in the state of Alagoas, between October 2014 and June 2015. In the laboratory, diaspores were cleaned, and the ones contaminated by pathogens and insects or presenting injuries were eliminated. In order to evaluate germination, the experiments were conducted using recently-harvested and ripeness-homogeneous diaspores.
Morphological characterization
For the morphological characterization (MF), 20 diaspores from each species were used, which were cut longitudinally to visualize their structures with an optical stereoscope. To do so, the differentiation of the diaspore tissues, the color and location (position) of the embryo were analyzed (Córner, 1976Córner E.J.H. The seeds of dicotyledons. Cambridge: University Press, 1976. v.1, 311p.).
Thousand diaspore weight
The weight of a thousand diaspores (TDW) was determined through weighing ten replications with 100 units. In addition, the humidity rate was determined in an oven at 105 ? 3 ºC for 24 hours, in five samples with 10 diaspores (Brazil, 2009Brasil. Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Brasília: Secretaria de Defesa Agropecuária, 2009. 395p. ).
Diaspore biometry
In order to perform the biometry, the length (mm) and thickness (mm) were determined, with the help of a digital caliper, in eight replications with 100 diaspores. The thickness was measured from the median line and the length in the top-bottom direction of the diaspore. For each variable, mean, mode, median, standard deviation, variance, variation coefficient and total amplitude were calculated (Araújo Neto et al., 2014Araújo Neto J.C. et al. Morphometric characterization, germination and conservation of Caesalpinia pulcherrima (L) SW. (Fabaceae: Caesalpinioidea) seeds. Semina: Ci Agr. 2014;35:2287-300. ). In order to indicate the distortion degree, the Pearson’s Coefficient of Skewness (ΔS) was calculated (Lorentz and Nunes, 2013Lorentz L.H., Nunes U.R. Relationship between quality measuring of rice seed batches. Rev Ci Agron. 2013;44:798-804.).
Asepsis for germination tests
Before the implantation of the temperature and luminosity experiments, the asepsis of the diaspores was performed; this consisted in the immersion in solutions of 70% ethanol (v/v) for a minute and 2% sodium hypochlorite for two minutes. For the luminosity experiment, asepsis was made under a green safety light, in order to avoid the effect of luminosity.
Temperature test
Tests were conducted with the completely randomized design in factor scheme 4 x 2 (temperature x diaspore condition), with 4 replications of 15 diaspores each. The used temperatures were the constant ones at 20, 25 and 30 oC and the alternated ones at 20-30 oC. The diaspore was used intact or with the removal of the epicarp, characterizing the two conditions. For each mistletoes species, an individual test was conducted. The used substrate was germitest paper, which was kept hydrated by adding distilled water in a quantity of 2.5 the initial weight of the paper (Brazil, 2009Brasil. Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Brasília: Secretaria de Defesa Agropecuária, 2009. 395p. ). The diaspores presenting the emission of the primary root were considered germinated. Counts were performed daily, and at the end of the experiment, the germination percentage (G (%)), germination rate index (GRI) and mean germination time (MGT) were calculated.
Light quality test
In the second experiment, diaspores (intact or without the pericarp) were incubated under lights (four fluorescent 20 w lamps) with different spectral qualities, for a photoperiod of 12 hours, at the alternated temperature of 20-30 oC for Passovia pyrifolia, Phoradendron mucronatum and at the constant temperature of 20 oC for Struthanthus marginatus, according to the result of the previous experiment. The tested treatments were: (L1) white light, (L2) red light, (L3) far-red light and (L4) dark. The red light was obtained covering the transparent plastic box (gerbox) with red cellophane, and the distant red light was obtained with red cellophane + blue cellophane. The dark one was obtained using black opaque gerboxes. The germination criterion and the analyzed variables were the same previously described. Except for the white light, the evaluation of the germination was made under a green safety life (20 w) (Lessa et al., 2013Lessa B.F.T. et al. Germination of Emilia coccinea (Sims) G. DON as a function of light, temperature, storage and sowing depth. Semina: Ci Agr . 2013;34:3193-3204. ).
Statistical analysis
Data from the germination testes were submitted to ANOVA and, if a significant effect of the treatments was observed, the averages were compared by Tukey’s test (p<0.05).
RESULTS AND DISCUSSION
The ripening of P. pyrifolia diaspores occurs from the distal portion towards the proximal portion, presenting a yellow color; at the end of ripening it presents an orange shade, both on the tip and at the bottom of the fruit (Figure 1A). Diaspores present viscin zone (ZN) between the epicarp and the endocarp. The endosperm (EN) and the embryo (EM), represented by cotyledons leaves with continuous axial position and chlorophyllaceous hypocotyl-radicle axis, are well detached by the contrast with the other structures of the diaspore (Figure 1B).

External vision of a Passovia pyrifolia fruit (A): Internal structure of P. pyrifolia (B): ZN - viscin zone; CE - stylus scar; EN - endosperm; EM - embryo; HT - hypocotyl radicle axis: Vascular bundles (C).
Externally, S. marginatus diaspores are elliptic berry fruit-type containing endosperm. The ripening occurs from the distal portion towards the proximal portion, which initially has a brownish color, progressing to the black color, reaching total black when completely ripe (Figure 2A). At this stage, the epicarp easily comes out when squeezed between the fingers. In the mesocarp there are the viscin zone (ZN), the endosperm (EN) involved by the epicarp is evident, as well as the embryo in the continuous axial position (EM) and the chlorophyllaceous hypocotyl-radicle axis (HT) inside the diaspore (Figure 2B).

Mistletoe fruit, Struthanthus marginatus (A): Internal structure of S. marginatus (B): CE - stylus scar; ZN - viscin zone; EN - endosperm; EM - embryo; HT - hypocotyl radicle axis.
Externally, diaspores of P. mucronatum, from the Santalaceae family, are characterized by an elliptic berry fruit-type format (ovoid), with wrinkly surface, with a single seed (monospermic) involved by a viscous tissue, with an easily removable epicarp when ripe. They present a well evident orange color during the ripening process from the distal portion to the proximal portion of the diaspore (Figure 3A). Recent studies conducted by Polli et al. (2016Polli A. et al. Structural development of the fruits and seeds in three mistletoe species of Phoradendron (Visceae: Santalaceae). Rodriguésia. 2016;67:649-59.) suggest a new terminology for fruits from the Phoradendron genus, denominated viscid-type pomaceous fruit. It presents a false seed, which is constituted by endosperm (EM), embryo (EN) in the axial position in a continuous format and chlorophyllaceous hypocotyl-radicle axis (HT) (Figure 3B). Verrucous fruits (like the previous species, presenting internally non-real seeds) are constituted by endosperm and chlorophyllaceous embryo (Caires and Dettke, 2010Caires C.S., Dettke G.A. Loranthacea. In: Lista de Espécies da Flora do Brasil. Rio de Janeiro: Jardim Botânico do Rio de Janeiro, 2010 http://floradobrasil.jbrj.gov.br/2010/FB000152.
http://floradobrasil.jbrj.gov.br/2010/FB...
).

Top view of Phoradendron mucronatum fruit (A): Side view of P. mucronatum (B): Internal structure of Phoradendron mucronatum (C) EN - chlorophyllaceous endosperm; EM - embryo; HT - hypocotyl-radicle axis: EM - swollen embryo (D).
From the species belonging to the Loranthaceae family, S. marginatus was the one presenting the highest weight, whereas P. mucronatum, belonging to the Santalaceae family, presented the lowest value among the three studied species. Values of thousand diaspore weight (TDW) of P. pyrifolia, S. marginatus and P. mucronatum, when converted to kg, correspond to 21,376, 18,632 and 2,976.19 diaspores per kg, respectively. For this variable, the three species presented a variation coefficient below the one required by the Rules for Seed Analysis (Brazil, 2009), which is 4% maximum (Table 1).
The species from the Loranthaceae family presented moisture content (MC) of the recently-harvested diaspores that were very close among themselves: P. pyrifolia with 62.97% and S. marginatus with 63.82. In P. mucronatum (Santalaceae), the moisture content of diaspores was 71.34% (Table 1). The high moisture content of diaspores is a common and important characteristic in mistletoes species, since they present a viscin layer, which contributes to the initial fixation on branches; therefore, it is a characteristic that is connected to the dispersion of these species.
Ripe diaspores of P. pyrifolia presented low variation in length and thickness measures, seen through the values of the central tendency. According to Pearson’s coefficient of skewness (ΔS), they presented small negative skewness (m0>md>m) for the length and symmetry for the thickness (m0<md=m). The length of S. marginatus diaspores presented moderated skewness according to the (ΔS), whereas the thickness showed a moderate asymmetric response (m0<md=m) according to the scale. P. mucronatum diaspores presented small negative asymmetry (m0<md>m) both for length and thickness, according to the (ΔS) (Table 2).
Descriptive statistics of the length and thickness, in mm, of recently-harvested diaspores of Passovia pyrifolia, Struthanthus marginatus and Phoradendron mucronatum
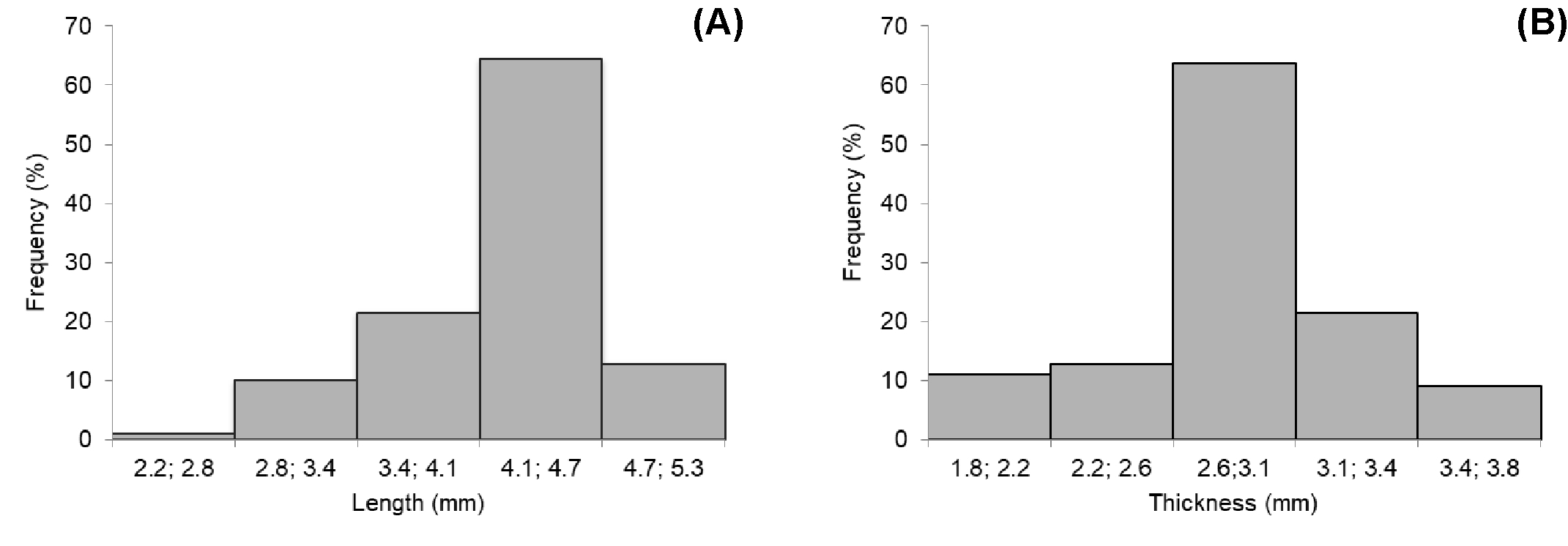
In the frequency histograms corresponding to data about length and thickness of P. pyrifolia diaspores, it is possible to observe that 64.5% of the length is distributed between 4.1 and 4.7 mm and, for the thickness, 63.8% are distributed between 2.69 and 3.09 mm. There was an asymmetric behavior of the curve for the length and a symmetric one for the thickness (Figure 4). For S. marginatus, it was possible to observe that 41% of the length of diaspores is distributed between 7.37 and 7.98 mm and 46% of the thickness between 4.71 and 5.17 mm. The length and thickness of the diaspores were asymmetric as for their distribution (Figure 5). In P. mucronatum diaspores, it was possible to observe that 51.11% of the length is distributed between 3.81 and 4.83 mm and 55.05% of the thickness between 2.39 and 2.92 mm. There was an asymmetric response for these variables (Figure 6).

Distribution of the relative frequencies of length (A) and thickness (B) of Passovia pyrifolia diaspores.

Distribution of the relative frequencies of length (A) and thickness (B) of Struthanthus marginatus.

Distribution of the relative frequencies of length (A) and thickness (B) of Phoradendron mucronatum diaspores.
As for the germination (G (%)) of P. pyrifolia, it was possible to observe the effect of temperature on the intact diaspores, where the alternated temperature of 20-30 oC was the one causing more germination (48.9%). In the diaspores submitted to the removal of the epicarp, which is one of the dispersion ways in the environment, temperature did not influence germination (Table 3). The germination percentage of diaspores that had their epicarp removed was always higher than the highest germination value of intact diaspores. Thus, knowing the dispersion way of P. pyrifolia, it is possible to deduce that the species will have a successful germination in a wide range of temperatures with the removal of the epicarp.
The GRI of P. pyrifolia presented a significant difference for different temperatures when the diaspores were intact, whereas when they were submitted to epicarp removal, there was no difference among the evaluated temperatures (Table 3). The MGT presented a similar response to the one previously observed: the temperature influenced when the diaspore was intact, whereas when the epicarp was removed, temperatures did not influence the mean germination time. The alternated temperature of 20-30 oC presented longer germination time: 4.4 days and 1.0 day in intact and epicarp-free diaspores, respectively (Table 3).
A possible explanation for that is that, according to Brancalion et al. (2005Brancalion P.H.S. Optimal temperature for seed germination of Brazilian tree species. Rev Bras Sementes. 2010;32:15-21.), most tropical species present an excellent germination rate between 20 oC and 30 oC. It is also possible that the requirements for environmental factors of hemiparasites are similar to the ones of their hosts. A consequence for this would be a wide temperature range for its development, in the case of P. pyrifolia, which is found as a parasite on Citrus sinensis.
The G (%) of S. marginatus was also influenced by the conditions of diaspores and by the different temperatures. In intact fruits, at the temperature of 20 oC there was 10% germination, and at the alternated temperature of 20-30 oC this value was 15%. There was no germination at the other tested temperatures. As for fruits without epicarp, G (%) was higher at 20 oC (90%), followed by the alternated temperature 20-30 oC (77.5%) (Table 4).
There was no GRI difference as for temperatures for the intact diaspore, whose values were lower compared to the diaspore with no epicarp; there were differences among the treatments, and the germination was quicker at the temperature of 20 oC, followed by the alternated temperature of 20 30 oC (Table 4). The MGT for intact diaspores presented differences among the temperatures, and the alternated 20-30 oC temperature presented germination on the first evaluation day. Diaspores without epicarp germinated quickly. One day after sowing, germination already occurred for the diaspores submitted to the temperatures of 30 oC and 20 oC. At the temperatures of 20 oC and 25 oC, the MGT was 1.4 and 1.2 days, respectively (Table 4).
The species S. marginatus was found in diversified environments, such as in woods and urban and agricultural areas (Citrus orchads), appearing established in the various environments. Temperature variations are expected between the environments; thus, the response of the species is related to its host and to the environment where it is located. According to Glatzel (1983Glatzel G. Mineral nutrition and water relations of hemiparasitic mistletoes: a question of partitioning. Experiments with Loranthus europaeus on Quercus petraea and Quercus robur. Oecologia. 1983;56:193-201.), there is a strict relation between hemiparasites and hosts.
The intact diaspores of P. mucronatum did not germinate, regardless of the tested temperature. After the fifth evaluation day, it was possible to observe their dryness and darkening, and on the 15th day they appeared completely deteriorated. However, diaspores that had their epicarp removed showed relatively high germination values; the alternated temperature of 20-30 oC stood out, providing 82.22% germination (Table 5). As occurred for G (%), MGT and GRI presented values only for diaspores with no epicarp (Table 5). The highest GRI was for diaspores with no epicarp submitted to the alternated temperature of 20-30 oC.
May (1971May D.S. The role of populational differentiation in experimental infection of Prosopis by Phoradendron. Am J Bot. 1971;58:921-1.) reports that Phoradendronspp. did not have a successful germination on different hosts under controlled environment conditions. However, the result of inoculations in an artificial environment may not reflect the host-parasite relation among natural populations on the field. In the case of the need to remove the epicarp demonstrated by P. mucronatum in this study, it may be due to the low oxygen quantity (in intact diaspores) to start the germination process, since the structures of the fruit may act as a barrier for the diffusion of this gas. Coder (2008Coder K.D. American Mistletoe (Phoradendron serotinum var. serotinum) Infection in Trees. WSFNR08, 2008. 37p. (Tree Helth Series)) states that oxygen is relatively low in seeds of Phoradendron serotinum var. serotinum intact fruits, and that this is overcome with the removal of the overlay by birds.
When evaluating the condition of diaspores from P. pyrifolia according to the quality of light, it was possible to observe the same tendency as the test with different temperatures, where diaspores without epicarp were always superior in the variables G (%) and germination rate. Quality did not influence diaspores without epicarp, whereas the red light increased the germination rate of intact diaspores (Table 6).
The responses of germination according to light generally vary between habitats, such as forests, where the presence of a strong light may be associated to the increase in the probability of seedling establishment and, thus, increase the germination percentage of some species (Flores et al., 2016Flores J. et al. Effect of light on seed germination and seedling shape of succulent species from Mexico. J.Plant Ecol. 2016;9:174-9.). However, in the present study, P. pyrifolia did not present this kind of pattern (germination only with light), which probably gives more germination capacity under different light conditions.
As for S. marginatus, the light quality altered the germination of intact and epicarp-free diaspores. The red light provided higher G (%) (80%) and GRI (7.3) for the diaspores without epicarp, whereas for the intact diaspores there was germination only with the white light (Table 7).
Plants may be classified, in terms of their response to germination, as: the ones requiring light to germinate (positively photoblastic); the ones requiring darkness to germinate (negatively photoblastic); and the ones having a great percentage of seeds that are neutral to light (neutrally photoblastic) (Baskin and Baskin, 2014Baskin C.C., Baskin J.M. Seeds: ecology, biogeography, and evolution of dormancy and germination. 2nd. ed. San Diego: Academic Press, 2014. 1600p.). Thus, intact diaspores of S. marginatus may be classified as positively photoblastic.
As for P. mucronatum, germination only occurred for diaspores without epicarp. Under this diaspore condition, the white light provided higher G (%) (40%) in relation to the other light qualities. Likewise, diaspores without epicarp had higher GRI (0.3) with the white light (Table 8).
In studies conducted by Messias et al. (2014Messias P.A. Host specificity and experimental assessment of the early establishment of the mistletoe Phoradendron crassifolium (Pohl ex DC.) Eichler (Santalaceae) in a fragment of Atlantic Forest in southeast Brazil. Acta Bot Bras. 2014;28:577-82.) with the germination of diaspores without epicarp, Phoradendron crassifolium under natural conditions (on different hosts) obtained average values (31%) that were lower than the ones observed in this study with P. mucronatum.
ACKNOWLEDGMENTS
This study was financed by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Edital MCTI/CNPQ/Universal 14/2014 - Faixa A.
REFERENCES
- Araújo Neto J.C. et al. Morphometric characterization, germination and conservation of Caesalpinia pulcherrima (L) SW. (Fabaceae: Caesalpinioidea) seeds. Semina: Ci Agr. 2014;35:2287-300.
- Aukema J.E. Vectors, Viscin, and Viscaceae: mistletoes as parasites, mutualists, and resources. Front Ecol Environ. 2003;1:212-9.
- Baskin C.C., Baskin J.M. Seeds: ecology, biogeography, and evolution of dormancy and germination. 2nd. ed. San Diego: Academic Press, 2014. 1600p.
- Brancalion P.H.S. Optimal temperature for seed germination of Brazilian tree species. Rev Bras Sementes. 2010;32:15-21.
- Brasil. Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Brasília: Secretaria de Defesa Agropecuária, 2009. 395p.
- Caires C.S., Dettke G.A. Loranthacea. In: Lista de Espécies da Flora do Brasil. Rio de Janeiro: Jardim Botânico do Rio de Janeiro, 2010 http://floradobrasil.jbrj.gov.br/2010/FB000152
» http://floradobrasil.jbrj.gov.br/2010/FB000152 - Cazzetta E., Galletti M. Frugivoria e especificidade por hospedeiro na erva-de-passarinho Phoradendron rubrun (L). Griseb (Viscaceae). Rev Bras Bot. 2007;30:321-50.
- Coder K.D. American Mistletoe (Phoradendron serotinum var. serotinum) Infection in Trees. WSFNR08, 2008. 37p. (Tree Helth Series)
- Córner E.J.H. The seeds of dicotyledons. Cambridge: University Press, 1976. v.1, 311p.
- Flores J. et al. Effect of light on seed germination and seedling shape of succulent species from Mexico. J.Plant Ecol. 2016;9:174-9.
- Glatzel G. Mineral nutrition and water relations of hemiparasitic mistletoes: a question of partitioning. Experiments with Loranthus europaeus on Quercus petraea and Quercus robur Oecologia. 1983;56:193-201.
- Harris R.W. Arboriculture: integrated management of landscape trees, shrubs and vines. New Jersey: Prentice-Hall, 1992. 674p.
- Lamont B. Host distribution, potassium content, water relations and control of two co-occurring mistletoe species. J R SocWest Aust. 1985; 68:21-5
- Lessa B.F.T. et al. Germination of Emilia coccinea (Sims) G. DON as a function of light, temperature, storage and sowing depth. Semina: Ci Agr . 2013;34:3193-3204.
- Lorentz L.H., Nunes U.R. Relationship between quality measuring of rice seed batches. Rev Ci Agron. 2013;44:798-804.
- May D.S. The role of populational differentiation in experimental infection of Prosopis by Phoradendron. Am J Bot. 1971;58:921-1.
- Messias P.A. Host specificity and experimental assessment of the early establishment of the mistletoe Phoradendron crassifolium (Pohl ex DC.) Eichler (Santalaceae) in a fragment of Atlantic Forest in southeast Brazil. Acta Bot Bras. 2014;28:577-82.
- Musselman L.J. et al. Parasitic plants major problem of food crops. Science. 2001;293:1434.
- Norton D.A. et al. Mistletoes as parasites: host specificity and speciation. Trends Ecol Evol. 1998;13:101-5.
- Okubamichael D.Y. et al. Reciprocal transplant experiment suggests host specificity of the mistletoe Agelanthus natalitius in South Africa. J Trop Ecol. 2014;30:153-63.
- Okubamichael D.Y. et al. Host specificity in parasitic plants - perspectives from mistletoes. AoB PLANTS. 2016; Sept 22:1-39
- Polli A. et al. Structural development of the fruits and seeds in three mistletoe species of Phoradendron (Visceae: Santalaceae). Rodriguésia. 2016;67:649-59.
- Press M.C. et al. Parasitic plants: physiological e ecological interactions whit their hosts. In: Press M.C. et al., editors Physiological plant ecology. Oxford: Blackwell Scientific, 1999. p.175-97.
- Soto-Gamboa M., Francisco Bozinovic F. Fruit-disperser interaction in a mistletoe-bird system: a comparison of two mechanisms of fruits processing on seed germination. Plant Ecol. 2002;159:171-4.
Publication Dates
-
Publication in this collection
2018
History
-
Received
24 Nov 2016 -
Accepted
15 Dec 2016




 ΔS = skewness coefficient; m = mean (mm); me = median (mm).
ΔS = skewness coefficient; m = mean (mm); me = median (mm).
 ΔS = skewness coefficient; m = mean (mm); me = median (mm).
ΔS = skewness coefficient; m = mean (mm); me = median (mm).
 ΔS = skewness coefficient; m = mean (mm); me = median (mm).
ΔS = skewness coefficient; m = mean (mm); me = median (mm).