ABSTRACT:
The cultivation and deposition of cover crops residues on the soil surface can reduce weed emergence. The objective of this study was to evaluate the emergence of weeds, in a greenhouse, on a soil with cover crops in an agroecological no-tillage system. The experiment was conducted in a greenhouse with soil samples from an experiment installed in Ituporanga (Santa Catarina state). On April 2010 and 2011, the following field treatments were implemented: weed vegetation (VE), Hordeum vulgare (2010)/Avena strigosa (2011) (CV/AV), Secale cereale (CE), Raphanus sativus (NF), Secale cereale + Raphanus sativus (NF+CE) and Hordeum vulgare/Avena strigosa + Raphanus sativus (NF+AV). After the rotation of the species, the soil samples were collected in July. The identification of the species and the emergence assessment occurred 15, 30, 45, 60, 75, 90 and 105 days after the beginning of the experiment. The species with the highest incidence were: Amaranthus lividus, Oxalis spp., Cyperus spp., Stachys arvensis, Cynodon spp. and Rumex obtusifolius. Residues from single species of Secale cereale, Avena strigosa and Hordeum vulgare have shown the greatest ability to suppress weeds during the first 45 evaluation days; this corresponds to the beginning of the critical period in the onion development.
Keywords:
green manuring; seed bank; Allium cepa
RESUMO:
O cultivo e a deposição dos resíduos de plantas de cobertura sobre a superfície do solo em sistema de plantio direto podem reduzir a emergência de plantas espontâneas. Este trabalho objetivou avaliar, em casa de vegetação, a emergência de plantas daninhas em solo com plantas de cobertura em sistema de plantio direto agroecológico de cebola. O experimento foi conduzido nos anos de 2010 e 2011, com amostras de solo de Ituporanga-SC. Em abril, foram implantados os tratamentos: testemunha com planta daninha (Test), cevada (2010)/aveia-preta (2011) (CV/AV), centeio (CE), nabo forrageiro (NF), nabo forrageiro + centeio (NF+CE) e nabo forrageiro + cevada/aveia-preta (NF+CV/NF+AV). Em julho, foram coletadas as amostras de solo. A identificação e a avaliação de emergência das espécies em casa de vegetação ocorreram aos 15, 30, 45, 60, 75, 90 e 105 dias após a instalação do experimento. As espécies com maior ocorrência foram: Amaranthus lividus, Cyperus spp., Oxalis spp., Stachys arvensis, Rumex obtusifolius e Galinsoga parviflora. Os resíduos das gramíneas centeio, cevada e aveia-preta solteiros sobre a superfície do solo apresentaram maior capacidade de supressão da emergência de plantas daninhas, principalmente nos primeiros 45 dias de avaliação, o que corresponde, em campo, ao início do período crítico de desenvolvimento da cebola e de maior competição com as plantas daninhas.
Palavras-chave:
adubação verde; banco de sementes; Allium cepa
INTRODUCTION
The conventional soil preparation system used in the main Brazilian regions and in the state of Santa Catarina is characterized by excessive soil plowing, as well as the indiscriminate use of herbicides to control weeds and to subsequently plant crops, for example onions (Altieri et al., 2011Altieri M.A. et al. Enhancing crop productivity via weed suppression in organic no-till cropping systems in Santa Catarina, Brazil. J Sust Agric. 2011;35:855-69.; Epagri, 2013Epagri. Sistema de produção para cebola: Santa Catarina (4ª. revisão). Florianópolis: 2013. 106p.). These management practices change the emergence, survival and redistribution of weed seeds on the surface and soil profile, contributing to the increase and maintenance of the seed bank (Monquero and Christoffoleti, 2005Monquero P.A., Christoffoleti P.J. Banco de sementes de plantas daninhas e herbicidas como fator de seleção. Bragantia. 2005;64:203-9. ), further aggravating the problem in successive crops.
As an alternative to the conventional tillage system, the cultivation of vegetables has been carried out in an agroecological no-tillage system, conducted with principles of ecological management. In this system, in addition to not using chemical synthesis fertilizers, single or intercropping cover crops are used instead of herbicides in order to control weeds (Kieling et al., 2009Kieling A.S. et al. Plantas de cobertura de inverno em sistema de plantio direto de hortaliças sem herbicidas: efeitos sobre plantas espontâneas e na produção de tomate. Ci Rural. 2009;39:2207-9.; Altieri et al., 2011Altieri M.A. et al. Enhancing crop productivity via weed suppression in organic no-till cropping systems in Santa Catarina, Brazil. J Sust Agric. 2011;35:855-69.).
Through the maintenance of covering plant residues, in the agroecological management, there is an attempt to eliminate and/or reduce chemical control. Covering plants can create a physical barrier on weeds through the amount of dry matter they produce, and by remaining on the soil surface (Queiroz et al., 2010Queiroz L.R. Supressão de plantas daninhas e produção de milho-verde orgânico em sistema de plantio direto. Planta Daninha. 2010;28:263-70.). The physical effect can also intervene in the germination and survival rate of the seedlings of some species, which require light to start the germination process. Often, nutrient reserves in seeds are not enough to guarantee the survival of the seedling inside the mulch until light is available to start the photosynthetic process (Monquero et al., 2009). The release of chemical compounds into the soil or compounds with allelopathic potential may also occur (Khanh et al., 2005Khanh T.D. et al. The exploitation of crop allelopathy in sustainable agricultural production. J Agron Crop Sci. 2005;191:172-84.; Lewinsohn and Gijzen, 2009Lewinsohn E., Gizen M. Phytochemical diversity: The sounds of silent metabolism. Plant Sci. 2009;176:161-9.; Hagemann et al., 2010Hagemann T.R. et al. Potencial alelopático de extratos aquosos foliares de aveia sobre azevém e amendoim-bravo. Bragantia. 2010;69:509-18. ); they are produced by cover plants that inhibit the emergence of weeds (Altieri et al., 2011Altieri M.A. et al. Enhancing crop productivity via weed suppression in organic no-till cropping systems in Santa Catarina, Brazil. J Sust Agric. 2011;35:855-69.; Camargo, 2013Camargo A.P. Aplicação de compostos polifenólicos de Canavalia ensiformis (L.) DC. e Mucuna aterrima (Piper & Tracy) Holland na germinação e na emergência de plantas espontâneas [dissertação]. Florianópolis: Universidade Federal de Santa Catarina, 2013. ; Bittencourt et al., 2013Bittencourt H.V.H. et al. Effect of winter cover crop biomass on summer weed emergence and biomass production. J Plant Protec Res. 2013;53:248-52.).
Predicting the emergence of these plants throughout the cycle and knowing which cover plant species can be used to reduce the emergence of weeds allow farmers to carry out proper crop management and weeding, and a reduction in the application of pre-emergence herbicides (Monquero and Christoffoleti, 2005Monquero P.A., Christoffoleti P.J. Banco de sementes de plantas daninhas e herbicidas como fator de seleção. Bragantia. 2005;64:203-9. ; Chauhan and Johnson, 2009Chauhan B.S., Johnson D. E. Influence of tillage systems on weed seedling emergence pattern in rainfed rice. Soil Till Res. 2009;106:15-21. ).
The study of weed species by sampling the soil seed bank, both in greenhouse and on the field, is an important parameter to estimate the quantity and diversity of the species found in the cultivated areas (Lousada et al., 2013Lousada L.L. et al. Correlation of soil properties with weed ocurrence in sugarcane areas. Planta Daninha. 2013;31:765-75. ). These studies, together with the adopted agronomic management, contribute to the reduction of the emergence and establishment of weeds during the crop cycle.
This work aimed at evaluating, in a greenhouse, the emergence of weeds in a soil with cover crops in an agroecological no - tillage system of onion crops.
MATERIAL AND METHODS
The study was conducted in a greenhouse in Florianópolis - Santa Catarina state, in 2010 and 2011, with samples of a humic Cambisol (Embrapa, 2013Empresa Brasileira de Pesquisa Agropecuária - Embrapa. Sistema Brasileiro de Classificação de Solos. 3ª.ed. Rio de Janeiro: Embrapa Solos, 2013. 312p.) and residues of winter cover crops from a long-term experiment, installed in an area in the municipality of Ituporanga - Santa Catarina state (latitude 27o24’52", longitude 49o36’9" and altitude 475 m).
The experiment from which the soil was collected was installed in an area with a 20-year history of onion cultivation in the conventional planting system (plowing and harrowing) until 1996. From that year on, the minimum tillage system was implemented, with the crop rotation of black oat (Avena strigosa), velvet bean (Mucuna aterrima), millet (Pennisetum glaucum), brown hemp (Crotalaria juncea) and common vetch (Vicia sativa). This system was maintained from 1996 to 2007, when it was replaced by the cultivation of sweet potatoes (Ipomoea batatas) until February 2009; in March, the experiment was installed, with an onion tillage system. In April 2009, weeds were desiccated using the herbicide glyphosate; then, limestone was applied and incorporated until the depth of 20 cm, in order to raise the pH in water to 6.0. Since then, no further applications of herbicides and mineral fertilizers were made in the experimental area.
The experimental design on the field was in randomized blocks with eight replications, and each experimental unit had 5 x 5 m. The treatments were: weed control sample (Test); barley (Hordeum vulgare) (CV) in 2010 and black oat (Avena strigosa) (AV) in 2011; rye (Secale cereale) (CE); radish (Raphanus sativus) (NF); radish + rye (NF + CE) and radish + barley (NF + CV) in 2010; and radish + black oat (NF + AV) in 2011. The species on the field were sown by broad casting on the soil surface; then a no-tillage cereal sowing machine passed twice on the area, in order to incorporate the seeds into the soil. Seed quantities were calculated based on the highest values, according to Monegat (1991Monegat C. Plantas de cobertura do solo: características e manejo em pequenas propriedades. Chapecó: Edição do Autor, 1991. 336p.) + 50%: black oat and barley (120 kg ha-1); rye (120 kg ha-1); radish (20 kg ha-1); radish + rye (10 + 60 kg ha-1); and radish + oat/barley (10 + 60 kg ha-1).
In July 2010 and 2011, soil samples were collected in order to evaluate the emergence of weeds in a greenhouse, according to a methodology adapted from Gasparino et al. (2006Gasparino D. et al. Quantificação do banco de sementes sob diferentes usos do solo em área de domínio ciliar. Rev Árvore. 2006;30:1-9.), in which a trench was opened with a shovel and, with a cutting metal plate and a knife, a soil sample of 37 x 27 x 10 cm (length x width x depth) was collected from each plot of the experiment. The residues of the cover plants from the same space were collected manually; this represented, on an average, 2.0 Mg ha-1 of weed vegetation; 2.5 Mg ha-1 of CV and 3.0 Mg ha-1 of AV; 3.0 Mg ha-1 of CE; 3.0 Mg ha-1 of NF; 4.3 and 3.5 Mg ha-1 of NF+CE; and 4.6 and 3.0 Mg ha-1 of NF+CV e NF+AV, per hectare, respectively.
The 48 soil samples were placed on black polypropylene trays, measuring 56 x 36 x 15 cm (length x width x height). During soil collection, the weeds contained in the samples were removed, and the trays were taken to the greenhouse. Each tray consisted of one experimental unit and they were placed on benches in a completely randomized design with eight replications and six treatments, and were replaced every 20 days.
The need for irrigation was defined through daily weighing and, when necessary, water was added to reach 70% of the field capacity, which had been previously determined in the laboratory. The temperature inside the greenhouse was maintained at the average of 20 oC. The emergence dynamics was determined according to the methodology proposed by Severino and Christoffoleti (2001Severino F.J., Christoffoleti P.J. Banco de sementes de plantas daninhas em solo cultivado com adubos verdes. Bragantia. 2001;60:201-4. ), in which all emerged weed seedlings were identified, counted and cut close to the soil surface 15, 30, 45, 60, 75, 90 and 105 days after the experiment was installed. The weed species that emerged throughout the experiment in the greenhouse were also identified, according to Lorenzi (2006Lorenzi H. Manual de identificação e controle de plantas daninhas: plantio direto e convencional. 6ª ed. Nova Odessa: Instituto Plantarum de Estudos da Flora, 2006. 339p.).
Statistical analysis
Data about the averages of weeds emerged in greenhouse were tested for normality using the Kolmogorov-Smirnov method, transformed using log (x), and then submitted to analysis of variance by the statistical software SISVAR (Ferreira, 2003Ferreira D.F. Programa de análises estatísticas (statistical analysis sotware) e planejamento de experimentos - SISVAR 5.0 (Build 67). Lavras: DEX/UFLA, 2003. ) . Averages were compared by Scott-Knott test (p<0.05).
RESULTS AND DISCUSSION
Emerged weed species
In 2010, 20 species and 15 weed families were identified and, in 2011, 21 species and 15 families were identified in the greenhouse soil samples, in 105 evaluation days (Table 1).
Weed species emerged in trays with soil and residues of cover plants during 105 evaluation days in a greenhouse, in 2010 and 2011
The species with the highest occurrence in all treatments in the two evaluation years were: Amaranthus lividus, Oxalisspp., Cyperusspp., Stachys arvensis, Cynodonspp., and Rumex obtusifolius (Table 1). These results agree with those obtained by Rowe (2006Rowe E. Manejo agroecológico da vegetação espontânea na cultura da cebola. In: Wordell Filho J.A. et al. Manejo fitossanitário na cultura da cebola. Florianópolis: Epagri, 2006. p.190-218.) in a Humic Cambisol and the main onion cultivation areas in the Alto Vale do Itajaí - Santa Catarina state (Epagri, 2013Epagri. Sistema de produção para cebola: Santa Catarina (4ª. revisão). Florianópolis: 2013. 106p.). These species are commonly found in studies about weeds in production systems, mainly because of their dissemination strategies (Lorenzi, 2006Lorenzi H. Manual de identificação e controle de plantas daninhas: plantio direto e convencional. 6ª ed. Nova Odessa: Instituto Plantarum de Estudos da Flora, 2006. 339p.; Cavigelli et al., 2008Cavigelli M.A. et al. Long-term agronomic performance of organic and conventional field crops in mid-Atlantic region. Agron J. 2008;100:785-94.). Studies conducted by Soares et al. (2004Soares D.J. et al. Efeito de diferentes períodos de controle das plantas daninhas na produtividade da cultura da cebola. Planta Daninha. 2004;22:517-27. ), while studying the interference periods of weeds in onion cultivation, found that the main emerged species were Amaranthus hybridus, Coronopus dydimus and Cyperus rotundus.
However, since this is a study in an agroecological no-tillage area, it is expected that, over the years, there will be a reduction in the diversity of weed species, due to the lack of harrowing, which promotes changes in the weed community. These changes involve aspects of the biology and ecology of the species, which can be altered by the soil management conditions and by the used cover crops (Voll et al., 2005Voll E. et al. A dinâmica das plantas daninhas e práticas de manejo. Londrina: 2005. 85p.). According to Voll et al. (2001Voll E. et al. Dinâmica do banco de sementes de plantas daninhas sob diferentes sistemas de manejo de solo. Planta Daninha. 2001;19:171-8. ), seeds such as the ones from pterogyne and beggar’s tick usually have a high germination and emergence rate, and are depleted in the soil in about three to four years.
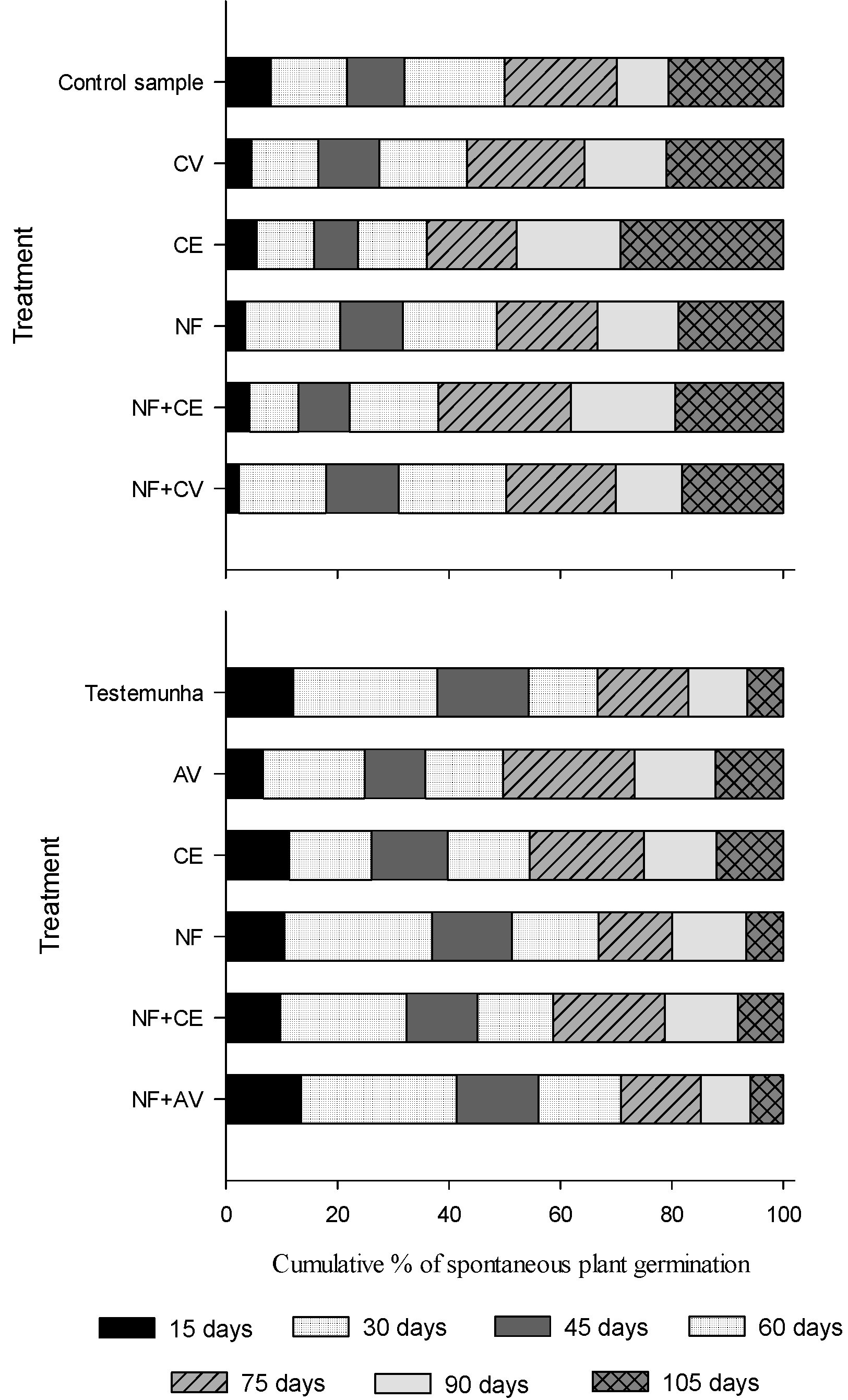
Weed emergence dynamics
Generally speaking, the emergence averages of weeds that emerged at 15, 30 and 45 days of evaluation, in 2010 and 2011, were lower in treatments with residues of rye, barley, black oat and radish intercropped with rye, and the highest averages were found in the control sample (Table 2). In 2010, it was observed that, out of the total weeds emerged in the control sample, 30% had already emerged during the first 30 evaluation days, 50% at 45 days, and 70% at 60 days (Figure 1). In 2011, 40% of all weeds emerged during the first 30 evaluation days; 60% at 45 days, which on the field corresponds to the period of greatest competitiveness with onion; and 70% at 60 days (Figure 1). This result may be due to the non-use of cover crops, leaving the soil exposed and favoring the emergence of weeds throughout the whole period. Studies conducted by Qasem (2006Qasem J.R. Response of onion (Allium cepa L.) plants to fertilizers, weed competition duration and planting times in the central Jordan Valley. Weed Biol Manage. 2006;6:212-20. ) showed that in 42 days of competition with weeds, there is a 5 Mg ha-1 reduction in the onion bulb production, compared to 21 days of competition.

Cumulative percentage of the germination averages of weeds emerged in soil samples with residues of cover crop species cultivated in an agroecological no-tillage system onion crop at 15, 30, 45, 60, 75, 90, and 105 evaluation days, in a greenhouse.
As for the different evaluation periods, it was possible to observe variations in the percentage of weeds that emerged throughout the first 30 and 45 evaluation days (Figure 1), as well as in the average of emerged seedlings, which in 2010 was lower in the treatment with rye residues (Table 2). In this treatment, only 25% of the total weeds emerged in the first 30 days, 30% at 45 days, and 50% at 60 evaluation days, differing from the other treatments (Figure 1A). In addition to this, at 75 evaluation days, the soil with rye residues presented 50% of the total emerged plants, while for the remaining treatments, 70% of the total weeds had already emerged in the same period (Figure 1A). Rowe (1997Rowe E. Avaliação de plantas de cobertura e da comunidade infestante em duas situações de cultivo [dissertação]. Florianópolis: Universidade Federal de Santa Catarina, 1997.), in an experiment conducted in Ituporanga - Santa Catarina state with different cover crops under no-tillage system, found that rye showed greater suppression of spontaneous plants. This result is due to the presence of cover crop species, and also to the 26.24 C/N ratio of rye plant material (Martins et al., 2014Martins R.P. et al. Mineralização do nitrogênio de plantas de cobertura, solteiras e consorciadas, depositadas sobre um solo com histórico de cultivo de cebola. Rev Ceres. 2014;61:587-96. ), which resulted in a longer permanence of residues on the soil surface. The persistence of rye straw corroborates what was observed by Oliveira et al. (2016Oliveira R.A. Cover crops effects on soil chemical properties and onion yield. Rev Bras Ci Solo. 2016;40:1-17.), who, when evaluating the decomposition of cover crops with the soil of the same experimental area, observed that rye residues remained on the soil surface for a longer time, around 90 days, throughout the onion cycle, and weeds remained for 30 days.
In 2011, the lowest averages of weeds that emerged at 45 days were observed in treatments with single black oat and rye residues (Table 2). Around 25% of the total weeds emerged during 30 evaluation days in rye and black oat; 40% of the total, at 45 days; and 50% of the total, at 60 evaluation days (Figure 1B). These results can be attributed to the higher C/N ratio of the plant material (Oliveira et al., 2016Oliveira R.A. Cover crops effects on soil chemical properties and onion yield. Rev Bras Ci Solo. 2016;40:1-17.), as well as the secondary compounds produced by these species (Jacobi and Fleck, 2000Jacobi U.S., Fleck N.G. Avaliação do potencial alelopático de genótipos de aveia no início do ciclo. Pesq Agropec Bras. 2000;35:11-9. ; Hagemann et al., 2010Hagemann T.R. et al. Potencial alelopático de extratos aquosos foliares de aveia sobre azevém e amendoim-bravo. Bragantia. 2010;69:509-18. ). Thus, this characteristic may have contributed to reduce the incidence of weeds through the decomposition of dry matter, releasing phenolic compounds that may have an inhibitory effect on weeds (Alvarenga et al., 2001Alvarenga R.C. et al. Plantas de cobertura de solo para sistema plantio direto. Inf Agropec. 2001;22:25-36.; Hagemann et al., 2010; Bittencourt et al., 2013Bittencourt H.V.H. et al. Effect of winter cover crop biomass on summer weed emergence and biomass production. J Plant Protec Res. 2013;53:248-52.).
The lower incidence of weeds both in 2010 and 2011 in the treatment with rye residues is also probably due to the amount of dry matter produced by this species: 2.0 Mg ha-1 at 60 days and 3.4 Mg ha-1 at 95 days after sowing (Souza et al., 2013Souza M. et al. Matéria seca de plantas de cobertura, produção de cebola e atributos químicos do solo em sistema de plantio direto agroecológico. Ci Rural. 2013;43:21-7.). Studies conducted by Gatiboni et al. (2009Gatiboni L.C. et al. Modificações na fauna edáfica durante a decomposição da palhada de centeio e aveia-preta, em sistema plantio direto. Rev Bragantia. 2009;22:45-53.) in Chapecó - Santa Catarina state show that, 88 days after lodging, black oat and rye presented, respectively, 58 and 60% of the initial amounts of dry matter on the soil. The same authors estimated that, 109 days after lodging, 50% of rye dry matter still remained on the soil.
However, in addition to grasses, the intercropped radish-rye treatment also showed low weed emergence at 45 days in 2011 (Table 2), where 45% of the weed total emerged during the first 45 evaluation days (Figure 1B). In this case, the persistence of radish dry matter on the soil was favored by the presence of grasses, such as rye, which contributed to a slower decomposition, probably due to the increase in the C/N ratio of the plant residue formed on the soil (Oliveira et al., 2016Oliveira R.A. Cover crops effects on soil chemical properties and onion yield. Rev Bras Ci Solo. 2016;40:1-17.), hindering the emergence of weeds. In studies conducted by Vilanova et al. (2014Vilanova C.C. et al. Interferência de plantas de cobertura sobre a incidência de plantas invasoras e a produção de cebola sob sistema de plantio direto. Sci Agr. 2014;15:9-14.) in field experiments, it was observed that the lowest dry matter productions of spontaneous plants during the onion cycle in 2009 occurred in the intercropped radish + rye treatment during the first 30 evaluation days.
At 60, 75, 90 and 105 days in 2010 and 2011, there were no differences among the treatments as for the average number of emerged seedlings (Table 2), since in these evaluation periods, the soil of all treatments was more exposed, due to the degradation of the residues that were on the surface which, when present, prevented the emergence of seedlings. According to Oliveira et al. (2016Oliveira R.A. Cover crops effects on soil chemical properties and onion yield. Rev Bras Ci Solo. 2016;40:1-17.), the remaining dry matter of the cover crop residues decreases over time. According to these authors, this happens because of the leaching of water-soluble compounds and the degradation of plant material by the microbial population. Most microorganisms, particularly fungi and bacteria, colonize plant residues, and part of the total organic carbon is used during decomposition as an energy source, whereas another part is released into the atmosphere as CO2 (Manzoni et al., 2008Manzoni S. et al. The global stoichiometry of litter nitrogen mineralization. Science. 2008;21:684-6.).
The emergence dynamics of weeds in the treatment with radish residues, both in 2010 and 2011, presented a similar behavior to the control sample in the different evaluation periods (Figure 1A, B). The species was more efficient during the first 15 days in 2010 (Table 2); since it presented a lower C/N ratio, of 16.3, it remained on the soil for less time, and this is reflected in a faster degradation of the material (Martins et al., 2014Martins R.P. et al. Mineralização do nitrogênio de plantas de cobertura, solteiras e consorciadas, depositadas sobre um solo com histórico de cultivo de cebola. Rev Ceres. 2014;61:587-96. ). As a result, the soil is exposed more quickly during the first 30 days and this, consequently, helps the emergence of weeds. According to Oliveira et al. (2016Oliveira R.A. Cover crops effects on soil chemical properties and onion yield. Rev Bras Ci Solo. 2016;40:1-17.), in an experiment conducted with the same species from the experimental area, radish residues remained on the soil surface for about 30 days.
As for the evaluation years, the average of emerged weeds was higher in 2011 (Table 2). Since cover crop species on the field were sown in 2010 and 2011 with the same seed quantities, the increase in the mean number of seedlings emerged in 2011 can be attributed to other factors, such as light and temperature. According to Vieira et al. (2010Vieira E.L. et al. Manual de fisiologia vegetal. São Luís: 2010. p.186.), the incidence of light and temperature fluctuations may affect the emergence of weeds.
In 2011, the incidence of artificial light in the greenhouse, due to other experiments at the site, probably altered the emergence dynamics of some species, reflecting higher weed emergence averages compared to 2010. This effect was more remarkable on the species Amaranthus lividus, which could be found during all evaluation. This species, when subjected to favorable conditions of temperature, humidity and light, can germinate more quickly. Moreover, the rapid takeover of the trays by the Amaranthus genus during the experiment demonstrated its competitive capacity. Species with a C4 photosynthetic metabolism, such as Amaranthus, are more efficient in the use of atmospheric CO2, and use light energy better, as well as having greater competitive ability in relation to C3 plants (Vieira et al. 2010Vieira E.L. et al. Manual de fisiologia vegetal. São Luís: 2010. p.186.), such as onion.
Finally, the results of the study indicate that residues of rye, barley and black oat alone on the soil surface showed a greater capacity of suppressing weed emergence, especially during the first 45 evaluation days; on the field, this corresponds to the beginning of the critical development period of the onion and the period of greater competition with weeds.
REFERENCES
- Altieri M.A. et al. Enhancing crop productivity via weed suppression in organic no-till cropping systems in Santa Catarina, Brazil. J Sust Agric. 2011;35:855-69.
- Alvarenga R.C. et al. Plantas de cobertura de solo para sistema plantio direto. Inf Agropec. 2001;22:25-36.
- Bittencourt H.V.H. et al. Effect of winter cover crop biomass on summer weed emergence and biomass production. J Plant Protec Res. 2013;53:248-52.
- Camargo A.P. Aplicação de compostos polifenólicos de Canavalia ensiformis (L.) DC. e Mucuna aterrima (Piper & Tracy) Holland na germinação e na emergência de plantas espontâneas [dissertação]. Florianópolis: Universidade Federal de Santa Catarina, 2013.
- Cavigelli M.A. et al. Long-term agronomic performance of organic and conventional field crops in mid-Atlantic region. Agron J. 2008;100:785-94.
- Chauhan B.S., Johnson D. E. Influence of tillage systems on weed seedling emergence pattern in rainfed rice. Soil Till Res. 2009;106:15-21.
- Epagri. Sistema de produção para cebola: Santa Catarina (4ª. revisão). Florianópolis: 2013. 106p.
- Empresa Brasileira de Pesquisa Agropecuária - Embrapa. Sistema Brasileiro de Classificação de Solos. 3ª.ed. Rio de Janeiro: Embrapa Solos, 2013. 312p.
- Ferreira D.F. Programa de análises estatísticas (statistical analysis sotware) e planejamento de experimentos - SISVAR 5.0 (Build 67). Lavras: DEX/UFLA, 2003.
- Gasparino D. et al. Quantificação do banco de sementes sob diferentes usos do solo em área de domínio ciliar. Rev Árvore. 2006;30:1-9.
- Gatiboni L.C. et al. Modificações na fauna edáfica durante a decomposição da palhada de centeio e aveia-preta, em sistema plantio direto. Rev Bragantia. 2009;22:45-53.
- Hagemann T.R. et al. Potencial alelopático de extratos aquosos foliares de aveia sobre azevém e amendoim-bravo. Bragantia. 2010;69:509-18.
- Jacobi U.S., Fleck N.G. Avaliação do potencial alelopático de genótipos de aveia no início do ciclo. Pesq Agropec Bras. 2000;35:11-9.
- Khanh T.D. et al. The exploitation of crop allelopathy in sustainable agricultural production. J Agron Crop Sci. 2005;191:172-84.
- Kieling A.S. et al. Plantas de cobertura de inverno em sistema de plantio direto de hortaliças sem herbicidas: efeitos sobre plantas espontâneas e na produção de tomate. Ci Rural. 2009;39:2207-9.
- Lewinsohn E., Gizen M. Phytochemical diversity: The sounds of silent metabolism. Plant Sci. 2009;176:161-9.
- Lorenzi H. Manual de identificação e controle de plantas daninhas: plantio direto e convencional. 6ª ed. Nova Odessa: Instituto Plantarum de Estudos da Flora, 2006. 339p.
- Lousada L.L. et al. Correlation of soil properties with weed ocurrence in sugarcane areas. Planta Daninha. 2013;31:765-75.
- Manzoni S. et al. The global stoichiometry of litter nitrogen mineralization. Science. 2008;21:684-6.
- Martins R.P. et al. Mineralização do nitrogênio de plantas de cobertura, solteiras e consorciadas, depositadas sobre um solo com histórico de cultivo de cebola. Rev Ceres. 2014;61:587-96.
- Monegat C. Plantas de cobertura do solo: características e manejo em pequenas propriedades. Chapecó: Edição do Autor, 1991. 336p.
- Monquero P.A., Christoffoleti P.J. Banco de sementes de plantas daninhas e herbicidas como fator de seleção. Bragantia. 2005;64:203-9.
- Oliveira R.A. Cover crops effects on soil chemical properties and onion yield. Rev Bras Ci Solo. 2016;40:1-17.
- Qasem J.R. Response of onion (Allium cepa L.) plants to fertilizers, weed competition duration and planting times in the central Jordan Valley. Weed Biol Manage. 2006;6:212-20.
- Queiroz L.R. Supressão de plantas daninhas e produção de milho-verde orgânico em sistema de plantio direto. Planta Daninha. 2010;28:263-70.
- Rowe E. Manejo agroecológico da vegetação espontânea na cultura da cebola. In: Wordell Filho J.A. et al. Manejo fitossanitário na cultura da cebola. Florianópolis: Epagri, 2006. p.190-218.
- Rowe E. Avaliação de plantas de cobertura e da comunidade infestante em duas situações de cultivo [dissertação]. Florianópolis: Universidade Federal de Santa Catarina, 1997.
- Severino F.J., Christoffoleti P.J. Banco de sementes de plantas daninhas em solo cultivado com adubos verdes. Bragantia. 2001;60:201-4.
- Soares D.J. et al. Efeito de diferentes períodos de controle das plantas daninhas na produtividade da cultura da cebola. Planta Daninha. 2004;22:517-27.
- Souza M. et al. Matéria seca de plantas de cobertura, produção de cebola e atributos químicos do solo em sistema de plantio direto agroecológico. Ci Rural. 2013;43:21-7.
- Vieira E.L. et al. Manual de fisiologia vegetal. São Luís: 2010. p.186.
- Vilanova C.C. et al. Interferência de plantas de cobertura sobre a incidência de plantas invasoras e a produção de cebola sob sistema de plantio direto. Sci Agr. 2014;15:9-14.
- Voll E. et al. Dinâmica do banco de sementes de plantas daninhas sob diferentes sistemas de manejo de solo. Planta Daninha. 2001;19:171-8.
- Voll E. et al. A dinâmica das plantas daninhas e práticas de manejo. Londrina: 2005. 85p.
Publication Dates
-
Publication in this collection
2018
History
-
Received
30 Aug 2016 -
Accepted
23 May 2017