
ABSTRACT:
A pot experiment was carried out to check the effect of Coronopus didymus (L.) Sm. dry biomass application (1%, 2% and 3% w/w) and two species of Trichoderma (T. viride and T. aureoviride) on growth and physiology of mungbean [Vigna radiata (L.) WIlczek] under biotic stress of Macrophomina phaseolina (Tassi) Goid. Inoculation of M. phaseolina (positive control) reduced plant survival, shoot and root length as well as plant dry biomass by 22%, 52%, 61% and 64%, respectively, over the negative control (without any amendment). There was 100% plant survival in treatments with T. aureoviride alone or in combination with 1% and 2% C. didymus biomass. Likewise, T. viride in combination with 2% biomass also showed 100% plant survival. Application of 3% C. didymus biomass had a pronounced effect on crop growth resulting in 101%, 233% and 342% increase in shoot length, root length and plant biomass, respectively, over the positive control. Sole inoculation of either of the two Trichoderma spp. significantly enhanced various plant growth parameters over the positive control. In general, in combination with 2% biomass of C. didymus, both Trichodermaspp. proved to be the best choice for improving mungbean biomass under stress of M. phaseolina. Activity of denfense related enzymes viz. peroxidase (PO), polyphenol oxidase (PPO) and phenylalanine ammonia lyase (PAL) was generally higher in treatments in which 1% C. didymus biomass was applied either alone or combined with Trichoderma spp. in M. phaseolina inoculated soil.
Keywords:
biological control; soil amendment; swinecress; Vigna radiata
RESUMO:
Um experimento em vaso foi realizado para verificar o efeito da aplicação da biomassa seca de Coronopus didymus (L.) Sm. (1%, 2% e 3% p/p) e duas espécies de Trichoderma (T. viride e T. aureoviride) sobre o crescimento e a fisiologia do feijão-mungo [Vigna radiata (L.) WIlczek] sob estresse biótico causado pelo fungo Macrophomina phaseolina (Tassi) Goid. A inoculação de M. phaseolina (controle positivo) reduziu a sobrevivência das plantas, o comprimento da parte aérea e da raiz e a biomassa seca das plantas em 22%, 52%, 61% e 64%, respectivamente, em relação ao controle negativo (sem qualquer alteração). Foram observados 100% de sobrevivência das plantas nos tratamentos com T. aureoviride isoladamente ou em combinação com 1% e 2% da biomassa de C. didymus. Da mesma forma, T. viride em combinação com 2% de biomassa também resultou em 100% de sobrevivência das plantas. A aplicação de 3% de biomassa de C. didymus teve efeito acentuado no crescimento da cultura, resultando em aumento de 101%, 233% e 342% no comprimento da parte aérea, comprimento da raiz e biomassa da planta, respectivamente, em relação ao controle positivo. A inoculação isolada de uma das duas espécies de Trichoderma spp. melhorou de forma significativa vários parâmetros de crescimento das plantas, em comparação ao controle positivo. Em geral, em combinação com 2% de biomassa de C. didymus, ambas as espécies de Trichoderma spp. provaram ser a escolha mais apropriada para melhorar a biomassa de feijão-mungo sob estresse causado por M. phaseolina. A atividade das enzimas relacionadas à defesa da planta, ou seja, peroxidase (PO), polifenol oxidase (PPO) e fenilalanina amônia liase (PAL), foi geralmente maior nos tratamentos em que 1% da biomassa de C. didymus foi aplicada isoladamente ou combinada com Trichoderma spp. em solo inoculado com M. phaseolina.
Palavras-chave:
controle biológico; correção do solo; mastruço; Vigna radiata
INTRODUCTION
Mungbean [Vigna radiata (L.) WIlczek] is one of the most important legumes in South Asian countries, particularly in Pakistan (Sue et al., 2015Sue KK, Nair RM, Lee J, Lee S-H. Genomic resources in mungbean for future breeding programs. Front Plant Sci. 2015;6:626-37.). It has two growing seasons and it is cultivated mostly in the districts of Bhakkar, Mianwali, Jhang, Khushab, Sargodha and in the Pothwar region. In Pakistan, it was grown on 130.9 thousand ha in 2013-2014, with annual production of 92.9 thousand ton (Pakistan, 2014Pakistan. Government of Pakistan. Federal Bureau of Statistics, Government of Pakistan. 2014.). It has high nutritional value as it contains 24-26% proteins, 51% carbohydrates, 3% vitamins and 4% minerals (Manoj et al., 2015Manoj K, Sachin K, Narendra K. Path analysis, association and variation of grain yield attributes in mungbean (Vigna radiata). Int J Adv Res. 2015;3:2410-13.). Its production is badly affected by charcoal rot disease, caused by Macrophomina phaseolina (Iqbal and Mukhtar, 2014Iqbal U, Mukhtar T. Morphological and pathogenic variability among Macrophomina phaseolina isolates associated with mungbean (Vigna radiata). Sci World J. 2014;1:27-35.). This soil-borne fungal plant pathogen has more than 500 hosts in different plant families with 67 hosts in Pakistan, including many economically important vegetables, oil-seed crops, pulses and ornamental flowers (Pawlowski et al., 2015Pawlowski ML, Hill CB, Hartman GL. Resistance to charcoal rot identified in ancestral soybean germplasm. Crop Sci Soc Am. 2015;55:1230-5.; Banaras et al., 2017Banaras S, Javaid A, Shoaib A, Ahmed E. Antifungal activity of Cirsium arvense extracts against a phytopathogenic fungus Macrophomina phaseolina. Planta Daninha. 2017;35: e017162738. ).
Use of synthetic fungicides is considered as the most important strategy for protecting plants against fungal pathogens. However, numerous fungicides available in the market are not only toxic but also pollute the environment (Anggriani et al., 2015Anggriani N, Putri NL, Supriatna AK. Stability analysis and optimal control of plant fungal epidemic: An explicit model with curative factor. AIP Conference Proceedings. 2015:40:1651.) and create resistance in fungal pathogens (Kim et al., 2007Kim BS, Hwang BK. Microbial fungicides in the control of plant diseases. J Phytopathol. 2007;155:641-53. ). A likely alternative to resolve such problems is the use of natural resources, including microorganisms and products from higher plants (De Corato et al., 2014De Corato U, Viola E, Arcieri G, Valerio V, Cancellara FA, Zimbardi F. Antifungal activity of liquid waste obtained from the detoxification of steam exploded plant biomass against plant pathogenic fungi. Crop Prot. 2014;55:109-118. ; Ali et al., 2017Ali A, Javaid A, Shoaib A. GC-MS analysis and antifungal activity of methanolic root extract of Chenopodium album against Sclerotium rolfsii. Planta Daninha. 2017;35: e017164713.; Khurshid et al., 2018Khurshid S, Javaid A, Shoaib A, Javed S, Qaiser U. Antifungal activity and GC-MS analysis of aerial parts of Cenchrus pennisetiformis against Fusarium oxysporum f. sp. lycopersici. Planta Daninha. 2018;36:e017166627.). Protecting plants against fungal pathogens using such alternatives may also reduce development of resistance in the pathogens (Trigui et al., 2013Trigui M, Hsouna AB, Hammami I, Culioli G, Ksantini M, Tounsi S. et al. Efficacy of Lawsonia inermis leaves extract and its phenolic compounds against olive knot and crown gall diseases. Crop Prot. 2013;45:83-8.). Members of the family Brassicaceae and Chenopodiaceae are especially known to control soil-borne plant pathogens when mixed in the soil before cultivation of crops (Javaid and Rauf, 2015Javaid A, Rauf S. Management of basal rot disease of onion with dry leaf biomass of Chenopodium album as soil amendment. Int J Agric Biol. 2015;17:142-8.; Javaid et al., 2017aJavaid A, Afzal L, Shoaib A. Biological control of charcoal rot of mungbean by Trichoderma harzianum and shoot dry biomass of Sisymbrium irio. Planta Daninha 2017a:35:e017165756.,bJavaid A, Niaz L, Shoaib A. Effect of incorporation of leaf biomass of Coronopus didymus on management of basal rot disease of onion and plant physiology. Int J Agric Biol. 2017b;19:445-52. ). Use of biological agents including fungi and bacteria are known to be very effective against Macrophomina root rot and other diseases (Talla et al., 2015Talla SG, Raju ASR, Karri S, Kumar YS. Production and antagonistic effect of Trichoderma spp. on pathogenic microorganisms (Botrytis cinerea, Fusarium oxysporium, Macrophomina phaseolina and Rhizoctonia solani). Afr J Biotechnol. 2015;14:668-75.; Munir et al., 2018Munir M, Shoaib A, Javaid A, Arshad Z, Rafiq M. Anti-mycotic potential of Trichoderma spp. and leaf biomass of Azadaricta indica against the charcoal rot pathogen Macrophomina phaseolina (Tassi) Goid in cowpea. Egyp J Biol Pest Control. 2018;28: 26.). Several fungi, especially species of Trichoderma (Javaid et al., 2014Javaid A, Afzal L, Bashir A, Shoaib A. A. In vitro screening of Trichoderma species against Macrophomina phasolina and Fusarium oxysporum f. sp. lycopersici. Pak J Phytopathol. 2014;26:39-43.; Walunj et al., 2015Walunj AA, Abhang PB, John P. In vitro evaluation of mutant and wild strain of Trichoderma harzianum against soil borne plant pathogen. Int J Plant Prot. 2015;8:108-11.), Penicillium(Murali and Amruthesh, 2015Murali M, Amruthesh KN. Plant growth promoting fungus Penicillium oxalicum enhances plant growth and induces resistance in pearl millet against downy mildew disease. J Phytopathol. 2015;163:743-54.) and Aspergillus (Kriaa et al., 2015Kriaa M, Sahnoun I, Azebou M, Triki MC, Kammoun MA. Purification, biochemical characterization and antifungal activity of a novel Aspergillus tubingensis glucose oxidase steady on broad range of pH and temperatures. Bioprocess and Biosystems Engineering, Springer Berlin Heidelberg Publisher, 2015.), are reported as effective biological control agents. The present study was, therefore, undertaken to evaluate the potential of application of the brassicaceous weed Coronopus didymus with either T. aureovirideor T. viride on plant survival, crop growth and physiology of mungbean in soil contaminated by M. phaseolina.
MATERIALS AND METHODS
Procurement of fungal species
Sub-culturing of M. phaseolina (FCBP 751), T. viride (FCBP 644) and T. aureoviride (FCBP 691) was performed on malt extract agar medium. Mass inoculum of each fungus was prepared on pre-boiled double autoclaved pearl millet seeds.
Collection of plant material
C. didymus plants were collected from different areas of the University of the Punjab, Lahore, Pakistan in February 2013. Plant material was sun-dried and thoroughly ground for further use in pot experiment.
Pot experiment
Pot trial was carried out by amending the soil with M. phaseolina, dried leaves of C. didymus and two species of Trichoderma, namely T. viride and T. aureoviride. The protocol given by Javaid and Saddique (2011Javaid A, Saddique A. Management of Macrophomina root rot of mungbean using dry leaves manure of Datura metel as soil amendment. Span J Agric Res. 2011:9:901-5.) was generally followed with some modifications. Soil was filled (2.0 kg pot-1) in earthen pots (20 cm diameter and 15 cm deep) and inoculated with inoculum of M. phaseolina (15 g pot-1) and watered. Likewise, T. viride and T. aureoviride inocula were also mixed in respective potted soil and watered. Later, after a week, soil was amended with C. didymus leaf biomass at 1%, 2% and 3% in the respective pots. Positive control consisted of the fungus (M. phaseolina) only whereas negative control was devoid of any inoculation or amendment. After 10 days, surface sterilized healthy mungbean seeds of uniform size were sown in each pot (10 seeds pot-1). A total of 13 treatments were used in the pot study: T1: Control; T2:+Control [only M. phaseolina (MP)]; T3: MP + 1% dry biomass of C. didymus (DBC); T4: MP + 2% DBC; T5: MP + 3% DBC; T6: MP + T. aureoviride (TA); T7: MP + 1% LDB + TA; T8: MP + 2% DBC + TA; T9: MP + 3% DBC + TA; T10: MP + T. viride (TV); T11: MP + 1% DBC + TV; T12: MP + 2% DBC + TV; T13: P + 3% DBC + TV. All the treatments were replicated thrice in a completely randomized design, kept under natural environmental conditions and watered whenever required.
Physiological tests
Various physiological tests were carried out after 35 days of growth just prior to flowering. For all the physiological parameters of the study, fresh leaves were taken from pot grown mungbean plants and, immediately after picking of leaves, physiological tests were performed on the fresh leaves. Total protein content was checked in leaf tissues (0.5 g), following the protocol of Baskaran et al. (2009Baskaran L, Chidambaram ALA, Sundaramoorthy P, Unnikannan P.. Bioreclamation of sugar mill effluent polluted soil using earthworm and its responses on green gram (Vigna radiata). J Phytol. 2009;1:7-12. ) by measuring absorbance at 650 nm using bovine serum albumin (BSA) as standards. Peroxidase (PO) activity was determined spectrophotometrically by using pyrogallol as a substrate (Kumar and Khan, 1982Kumar KB, Khan PA. Peroxidase in excised ragi (Eleusine coracana cv. PR 202) leaves during senescence. Ind J Exp Biol. 1982:20:412-6. ). The increase in absorbance resulting from formation of oxidized product (purpurogallin) was recorded at 420 nm. The reaction mixture [(2 mL of 0.1 M phosphate buffer (pH 6.8) + 1 mL of pyrogallol + 1 mL of 0.05 M H2O2)] was mixed with enzyme extract (0.5 mL). After incubation at 25 oC, 2.5 N H2SO4 (24.5 mL of H2SO4 + 100 mL of distilled water) was added in the reaction mixture. For estimation of polyphenol oxidase activity (PPO), the enzyme extract (100 μL) was mixed with 0.1 M of pH 7.0 sodium phosphate buffer (1.5 mL). The reaction started when 200 μL of 0.01 M catechol was added. The absorbance of the sample was measured at 30 sec interval for 3 min at 495 nm (Mayer et al., 1965Mayer AM, Harel E, Shaul RB. Assay of catechol oxidase: a critical comparison of methods. Photochemistry. 1965;5:783-9.). For determination of phenylalanine ammonia-lyase (PAL) activity, the reaction mixture [(0.4 mL of enzyme extract + 0.1 M sodium borate buffer (pH 8.8) + 0.5 mL of 12 mM L-1 phenylalanine)] was incubated for 1 h in light at 25 oC and the reaction was stopped by incubation at 47 oC for 10 min. The amount of resulting trans-cinnamic acid was calculated after measuring absorbance of the samples at 290 nm (Dickerson et al., 1984Dickerson DP, Pascholati SF, Hagerman AE, Butler LG, Nicholson RL. Phenylalanine ammonia lyase and hydroxycinnamate: CoA ligase in maize mesocotyls inoculated with Helminthosporium maydis or Helminthosporium carbonum. Physiol Plant Pathol. 1984;25:111-23. ).
Harvesting and data collection
After harvesting data on the number of surviving plants, shoot and root length, and plant dry weight were recorded.
Statistical data analysis
All the data were subjected to Analysis of variance (ANOVA) followed by the LSD test to separate treatment means at 5% level of significance.
RESULTS AND DISCUSSION
Effect of treatments on plant growth
ANOVA showed that there was a significant (P = 0.05 and 0.001) effect of treatments on plant survival, shoot length, root length and dry weight of mungbean plants (Table 1). There was 100% survival of plants in the negative control but that rate was reduced to 78% in the positive control. Application of 1% C. didymus further reduced survival percentage to 37%, which was increased to 67% and 73% by increasing the dose of C. didymus to 2% and 3%, respectively (Figure 1). Shoot length in control was 19.3 cm. Inoculation of M. phaseolina significantly reduced this growth parameter to 9.2 cm. In general, the effect of all the C. didymus biomass amendment treatments was significant on shoot length as compared to the positive control treatment. Application of 1%, 2% and 3% C. didymus biomass gradually increased shoot length to 14.9, 17.6 and 18.6 cm, respectively (Figure 2A). Root length in the negative control was 12.33 cm but it significantly decreased to 4.77 cm in positive control. Application of 1% C. didymus biomass had an insignificant effect on root length as compared to the positive control. However, further increase in C. didymus biomass to 2% and 3% significantly increased root length to 11.70 cm and 15.93 cm, respectively (Figure 2B). Inoculation of M. phaseolina caused a 64% decline in dry biomass of mungbean. Application of 1% biomass of C. didymus failed to change the adverse effect of M. phaseolina on the biomass of mungbean. However, a further increase in the dose of C. didymus biomass as soil amendment significantly increased plant biomass up to 342% over the positive control (Figure 2C). Earlier, Coelho de Souza (2004Coelho de Souza G, Haas APS, von Poser GL, Schapoval EES, Elisabetsky E. Ethnopharmacological studies of antimicrobial remedies in the south of Brazil. J Ethnopharmacol. 2004;90:135-43.) studied the antimicrobial activity of crude methanolic extracts of C. didymus against seven microorganisms and a significant result was achieved. Likewise, Iqbal and Javaid (2012Iqbal D, Javaid A . Bioassays guided fractionation of Coronopus didymus for its antifungal activity against Sclerotium rolfsii. Nat Prod Res. 2012;26:1638-44.) reported that methanolic extracts with concentrations of 15 mg mL-1 of leaf, stem, inflorescence and root of C. didymus reduced the biomass of Sclerotium rolfsii by 67%, 26%, 40% and 58%, respectively. Similarly, a 4% methanolic extract of C. didymus reduced biomass of Fusarium moniliforme by 48% (Javaid et al., 2018Javaid A, Latif U, Akhtar N, Ahmed D, Perveen S. Molecular characterization of Fusarium moniliforme and its management by methanolic extract of Coronopus didymus. Pak J Bot. 2018; 50:2069-75.). Khan et al. (2010Khan SN, Javaid A, Riaz T. Management of Fusarium corm rot of gladiolus (Gladiolus grandiflorus sect. Blandus cv. Aarti) by using leaves of allelopathic plants. Afr J Biotechnol. 2010;9:4681-6. ) studied the effect of C. didymus leaf incorporation on Fusarium corm rot of gladiolus. Different doses of this weed species significantly reduced disease incidence and mortality in gladiolus. Javaid and Iqbal (2014Javaid A, Iqbal D. Management of collar rot of bell pepper (Capsicum annuum L.) by extracts and dry biomass of Coronopus didymus shoot. Biol Agric Hortic. 2014;30:164-72.) reported that a 3% dose of dry leaves of C. didymus significantly reduced disease incidence and mortality by S. rolfsii in bell pepper to 50% and 13% as compared to 91% and 40% in the positive control, respectively. In a recent study, Javaid et al. (2017b) found that basal rot of onion caused by Fusarium oxysporum f. sp. cepae can effectively be managed by application of 2% C. didymus biomass as soil amendment. C. didymus contains glucotrapaeolin, a glucoside of benzyl isothiocyanate (Prabhakar et al., 2006Prabhakar KR, Veeresh VP, Vipan K, Sudheer M, Priyadarsini KI, Satish RB. et al. Bioactivity-guided fractionation of Coronopus didymus: A free radical scavenging perspective. Phytomedicine. 2006;13:591-5.), which possibly reduced the fungal inoculum in the rhizosphere and, thus, reduced disease incidence and mortality. Gamliel and Stapleton (1993Gamliel A, Stapleton JJ. Characterization of antifungal volatile compounds evolved from solarized soil amended with cabbage residues. Phytopthology. 1993;83:899-905.) demonstrated that members of the family Brassicaceae have the ability to control the growth of phytopathogenic fungi. These toxic effects are linked to the biologically active degradation products of glucosinolates, which are reported to be present in 16 families of dicotyledonous angiosperms, mainly in Brassicaceae (Fahey et al., 2001Fahey JW, Zalcmann AT, Talalay P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry, 2001;56:5-51.).

Effect of soil amendment with dry biomass of Coronopus didymus (DBC), Macrophomina phaseolina (MP) and two Trichoderma spp. species [T. aureoviride (TA) and T. viride (TV)] on survival percentage of mungbean plants.

Effect of soil amendment with dry biomass of Coronopus didymus (DBC), Macrophomina phaseolina (MP) and two Trichoderma spp. species [T. aureoviride (TA) and T. viride (TV)] on shoot and root growth of mungbean.
There was 100% plant survival in treatments with T. aureoviride alone or in combination with 1% and 2% C. didymus biomass as compared to 78% plant survival in the positive control. Likewise, T. viride in combination with 2% biomass also showed 100% plant survival. T. aureoviride with 3% biomass application significantly reduced plant survival while the effect of the combined application of T. viride and 3% biomass had an insignificant effect as compared to the negative control (Figure 1). T. aureoviride in combination with 1% and 2% biomass of C. didymus biomass showed maximum shoot length, i.e., 20.1 and 20.4 cm. However, T. aureoviride inoculated along with 3% C. didymus biomass abruptly declined shoot length to 12.1 cm. The effect of T. viride alone or combined with 1% and 2% C. didymus biomass on shoot length was less pronounced as compared to the effect of similar treatments with T. aureoviride (Figure 2A). Inoculation of T. aureoviride alone or combined with 1% and 2% C. didymus biomass significantly enhanced root length by 134-180% over the positive control. However, the effect of T. aureoviride + C. didymus biomass was less pronounced as there was 82% increase in root length over the positive control. Likewise, inoculation of T. viride alone or combined with different doses of C. didymus biomass significantly enhanced root length by 124-185% over the positive control (Figure 2B). Sole inoculation of either of the two Trichoderma species significantly increased mungbean biomass. However, the positive effect of T. aureoviride was more pronounced than the effect of T. viride. In combination with different doses of C. didymus, the two Trichoderma species showed different behaviours on plant biomass of mungbean. In combination with 2% C. didymus biomass, both species had a similar and highly pronounced effect on mungbean biomass. In combination with 3% C. didymus biomass, T. viride had a similar effect while T. aureoviride showed a markedly diminished effect on mungbean biomass (Figure 2C). Inhibition of pathogenic fungal growth by Trichodermaspp. occurs by physical as well as chemical interactions in which a variety of chemicals are released by Thrichodermaspp., inducing localized or systemic resistance responses in plants (Harman et al., 2004Harman GE, Howell CR, Viterbo A, Chet I, Lorito M. Trichoderma species opportunistic avirulent plant symbionts. Nat Rev Microbiol. 2004;2:40-3.). Faster metabolic rate of Trichodermaspp., competition for food and space, enzymatic antibiosis, release of secondary antimicrobial metabolites and physiological conformation are the key factors involved in the antagonistic interaction of Trichodermaspp. with pathogenic fungi (Verma et al., 2007Verma M, Brar SK, Tyagi RD, Surampalli RY, Valéro JR. Antagonistic fungi Trichoderma spp.: Panoply of biological control. Biochem Eng J. 2007;37:1-20).
Effect of treatments on plant physiology
ANOVA illustrates that different treatments had a significant effect (P=0.001) on the activities of peroxidase (PO), polyphenol oxidase (PPO) and phenylalanine ammonia lyase (PAL) of mungbean leaves (Table 1). The difference in PO activity of the negative and positive control treatments was insignificant. Addition of 1% C. didymus biomass alone or combination with T. aureoviride significantly increased PO activity. An increase in the dose of C. didymus biomass alone or combined with either of the two Trichoderma species gradually decreased PO activity (Figure 3A).

Effect of soil amendment with dry biomass of Coronopus didymus (DBC), Macrophomina phaseolina (MP) and two Trichoderma spp. species [T. aureoviride (TA) and T. viride (TV)] on activities of defense related enzymes of mungbean.
Similar to that of PO activity, difference in polyphenol oxidase (PPO) activity between negative and positive control was insignificant and application of 1% C. didymus biomass alone or combined with T. aureoviride significantly enhanced this activity. Higher doses of C. didymus biomass significantly reduced PPO activity (Figure 3B). The highest PAL activity was recorded in 1% C. didymus + T. aureoviride treatment. Generally, the increase in C. didymus biomass adversely affected this parameter (Figure 3C).
Plants infected by M. phaseolina only showed an insignificant effect in enzyme activity (POX, PPO and PAL) in the positive control as compared to the negative control as the susceptible host does not have the ability to detect the threat posed by pathogen (Fortunato et al., 2015Fortunato AA, Debona D, Bernardeli AM, Rodrigues FÁ. Changes in the antioxidant system in soybean leaves infected by Corynespora cassiicola. Phytopathology, 2015;105:1050-8.). A directly proportional relationship was found between enzyme production and disease suppression after application of different management agents attributed to a higher production of reactive oxygen species (ROS) and antioxidant enzymes in order to overcome stress. As the plant gets rid of the stress, enzyme production is also reduced with increase in biomass of C. didymus alone or along with either of the two Trichoderma species.
The present study concludes that in combination with 2% biomass of C. didymus, both Trichodermaspp. species have been proven to improve mungbean biomass under stress of M. phaseolina.
ACKNOWLEDGEMENTS
The University of the Punjab, Lahore, Pakistan, provided funding that was essential for the success of this project.
REFERENCES
- Ali A, Javaid A, Shoaib A. GC-MS analysis and antifungal activity of methanolic root extract of Chenopodium album against Sclerotium rolfsii Planta Daninha. 2017;35: e017164713.
- Anggriani N, Putri NL, Supriatna AK. Stability analysis and optimal control of plant fungal epidemic: An explicit model with curative factor. AIP Conference Proceedings. 2015:40:1651.
- Banaras S, Javaid A, Shoaib A, Ahmed E. Antifungal activity of Cirsium arvense extracts against a phytopathogenic fungus Macrophomina phaseolina Planta Daninha. 2017;35: e017162738.
- Baskaran L, Chidambaram ALA, Sundaramoorthy P, Unnikannan P.. Bioreclamation of sugar mill effluent polluted soil using earthworm and its responses on green gram (Vigna radiata). J Phytol. 2009;1:7-12.
- De Corato U, Viola E, Arcieri G, Valerio V, Cancellara FA, Zimbardi F. Antifungal activity of liquid waste obtained from the detoxification of steam exploded plant biomass against plant pathogenic fungi. Crop Prot. 2014;55:109-118.
- Dickerson DP, Pascholati SF, Hagerman AE, Butler LG, Nicholson RL. Phenylalanine ammonia lyase and hydroxycinnamate: CoA ligase in maize mesocotyls inoculated with Helminthosporium maydis or Helminthosporium carbonum Physiol Plant Pathol. 1984;25:111-23.
- Coelho de Souza G, Haas APS, von Poser GL, Schapoval EES, Elisabetsky E. Ethnopharmacological studies of antimicrobial remedies in the south of Brazil. J Ethnopharmacol. 2004;90:135-43.
- Fahey JW, Zalcmann AT, Talalay P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry, 2001;56:5-51.
- Fortunato AA, Debona D, Bernardeli AM, Rodrigues FÁ. Changes in the antioxidant system in soybean leaves infected by Corynespora cassiicola Phytopathology, 2015;105:1050-8.
- Gamliel A, Stapleton JJ. Characterization of antifungal volatile compounds evolved from solarized soil amended with cabbage residues. Phytopthology. 1993;83:899-905.
- Harman GE, Howell CR, Viterbo A, Chet I, Lorito M. Trichoderma species opportunistic avirulent plant symbionts. Nat Rev Microbiol. 2004;2:40-3.
- Pakistan. Government of Pakistan. Federal Bureau of Statistics, Government of Pakistan. 2014.
- Iqbal D, Javaid A . Bioassays guided fractionation of Coronopus didymus for its antifungal activity against Sclerotium rolfsii Nat Prod Res. 2012;26:1638-44.
- Iqbal U, Mukhtar T. Morphological and pathogenic variability among Macrophomina phaseolina isolates associated with mungbean (Vigna radiata). Sci World J. 2014;1:27-35.
- Javaid A, Saddique A. Management of Macrophomina root rot of mungbean using dry leaves manure of Datura metel as soil amendment. Span J Agric Res. 2011:9:901-5.
- Javaid A, Iqbal D. Management of collar rot of bell pepper (Capsicum annuum L.) by extracts and dry biomass of Coronopus didymus shoot. Biol Agric Hortic. 2014;30:164-72.
- Javaid A, Afzal L, Bashir A, Shoaib A. A. In vitro screening of Trichoderma species against Macrophomina phasolina and Fusarium oxysporum f. sp. lycopersici Pak J Phytopathol. 2014;26:39-43.
- Javaid A, Rauf S. Management of basal rot disease of onion with dry leaf biomass of Chenopodium album as soil amendment. Int J Agric Biol. 2015;17:142-8.
- Javaid A, Afzal L, Shoaib A. Biological control of charcoal rot of mungbean by Trichoderma harzianum and shoot dry biomass of Sisymbrium irio Planta Daninha 2017a:35:e017165756.
- Javaid A, Niaz L, Shoaib A. Effect of incorporation of leaf biomass of Coronopus didymus on management of basal rot disease of onion and plant physiology. Int J Agric Biol. 2017b;19:445-52.
- Javaid A, Latif U, Akhtar N, Ahmed D, Perveen S. Molecular characterization of Fusarium moniliforme and its management by methanolic extract of Coronopus didymus Pak J Bot. 2018; 50:2069-75.
- Khan SN, Javaid A, Riaz T. Management of Fusarium corm rot of gladiolus (Gladiolus grandiflorus sect. Blandus cv. Aarti) by using leaves of allelopathic plants. Afr J Biotechnol. 2010;9:4681-6.
- Khurshid S, Javaid A, Shoaib A, Javed S, Qaiser U. Antifungal activity and GC-MS analysis of aerial parts of Cenchrus pennisetiformis against Fusarium oxysporum f. sp. lycopersici Planta Daninha. 2018;36:e017166627.
- Kim BS, Hwang BK. Microbial fungicides in the control of plant diseases. J Phytopathol. 2007;155:641-53.
- Kriaa M, Sahnoun I, Azebou M, Triki MC, Kammoun MA. Purification, biochemical characterization and antifungal activity of a novel Aspergillus tubingensis glucose oxidase steady on broad range of pH and temperatures. Bioprocess and Biosystems Engineering, Springer Berlin Heidelberg Publisher, 2015.
- Kumar KB, Khan PA. Peroxidase in excised ragi (Eleusine coracana cv. PR 202) leaves during senescence. Ind J Exp Biol. 1982:20:412-6.
- Manoj K, Sachin K, Narendra K. Path analysis, association and variation of grain yield attributes in mungbean (Vigna radiata). Int J Adv Res. 2015;3:2410-13.
- Mayer AM, Harel E, Shaul RB. Assay of catechol oxidase: a critical comparison of methods. Photochemistry. 1965;5:783-9.
- Munir M, Shoaib A, Javaid A, Arshad Z, Rafiq M. Anti-mycotic potential of Trichoderma spp. and leaf biomass of Azadaricta indica against the charcoal rot pathogen Macrophomina phaseolina (Tassi) Goid in cowpea. Egyp J Biol Pest Control. 2018;28: 26.
- Murali M, Amruthesh KN. Plant growth promoting fungus Penicillium oxalicum enhances plant growth and induces resistance in pearl millet against downy mildew disease. J Phytopathol. 2015;163:743-54.
- Pawlowski ML, Hill CB, Hartman GL. Resistance to charcoal rot identified in ancestral soybean germplasm. Crop Sci Soc Am. 2015;55:1230-5.
- Prabhakar KR, Veeresh VP, Vipan K, Sudheer M, Priyadarsini KI, Satish RB. et al. Bioactivity-guided fractionation of Coronopus didymus: A free radical scavenging perspective. Phytomedicine. 2006;13:591-5.
- Sue KK, Nair RM, Lee J, Lee S-H. Genomic resources in mungbean for future breeding programs. Front Plant Sci. 2015;6:626-37.
- Talla SG, Raju ASR, Karri S, Kumar YS. Production and antagonistic effect of Trichoderma spp. on pathogenic microorganisms (Botrytis cinerea, Fusarium oxysporium, Macrophomina phaseolina and Rhizoctonia solani). Afr J Biotechnol. 2015;14:668-75.
- Trigui M, Hsouna AB, Hammami I, Culioli G, Ksantini M, Tounsi S. et al. Efficacy of Lawsonia inermis leaves extract and its phenolic compounds against olive knot and crown gall diseases. Crop Prot. 2013;45:83-8.
- Verma M, Brar SK, Tyagi RD, Surampalli RY, Valéro JR. Antagonistic fungi Trichoderma spp.: Panoply of biological control. Biochem Eng J. 2007;37:1-20
- Walunj AA, Abhang PB, John P. In vitro evaluation of mutant and wild strain of Trichoderma harzianum against soil borne plant pathogen. Int J Plant Prot. 2015;8:108-11.
Publication Dates
-
Publication in this collection
2018
History
-
Received
27 July 2017 -
Accepted
08 Aug 2017

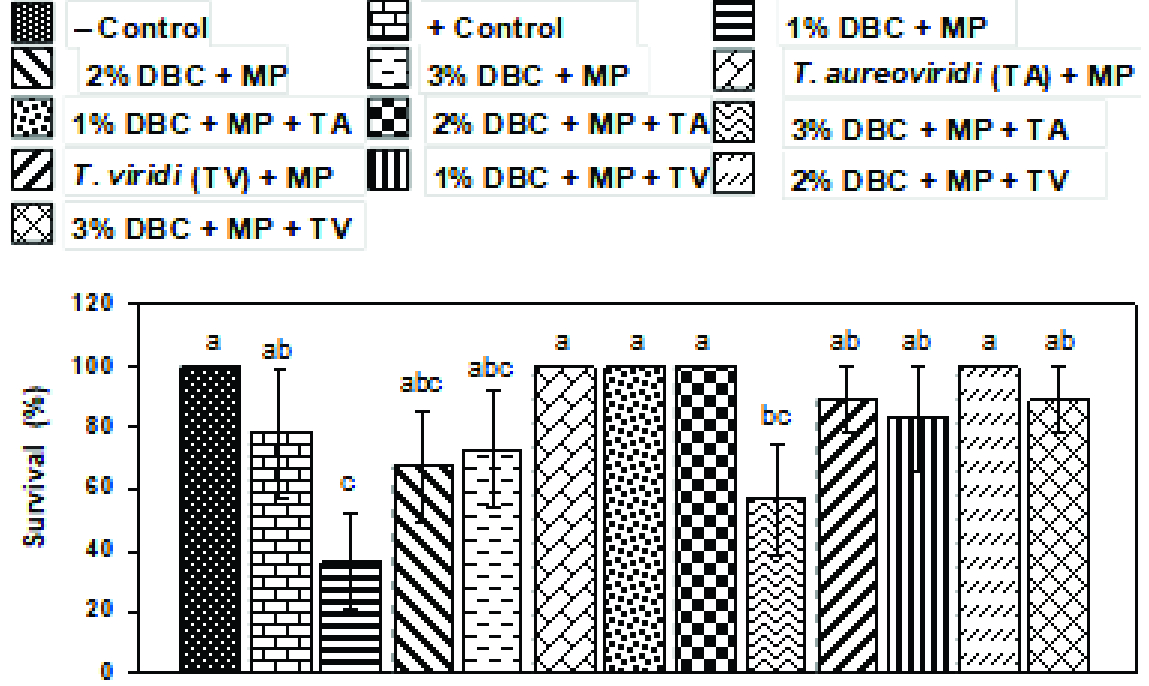 Vertical bars show standard errors of means of five replicates. Values with different letters at the top show a significant difference (P≤0.05) as determined by the LSD Test.
Vertical bars show standard errors of means of five replicates. Values with different letters at the top show a significant difference (P≤0.05) as determined by the LSD Test.
 Vertical bars show standard errors of means of five replicates. Values with different letters at the top show a significant difference (P≤0.05) as determined by the LSD Test.
Vertical bars show standard errors of means of five replicates. Values with different letters at the top show a significant difference (P≤0.05) as determined by the LSD Test.
 Vertical bars show standard errors of means of five replicates. Values with different letters at the top show a significant difference (P≤0.05) as determined by the LSD Test.
Vertical bars show standard errors of means of five replicates. Values with different letters at the top show a significant difference (P≤0.05) as determined by the LSD Test.