
ABSTRACT:
Volunteer corn is considered a weed when its emergence occurs during the crop cycle in succession, as is the case of volunteer corn plants of hybrids of Roundup Ready (RR) in crops implanted with RR soybean cultivars. The objective of this study was to verify the effect of the interference of different RR plant densities on the RR soybean crop, as well as to estimate the level of economic damage (NDE). For this, a field experiment was carried out at two sites in a factorial scheme with four replications, the treatments consisted of densities of volunteer corn RR (0; 1; 2; 3; 4; 5; 6; 7 and 8 plants m-2) competing in the entire soybean cycle. The changes in morphology and yield components behaved directly proportional to the increment of volunteer corn densities, with the variable vegetables per plant being the most strongly affected. The loss of soybean yield at the highest competitor density reached 58%. NDE was closely dependent on the cost of control of volunteer corn plants, crop yield expectancy, herbicide efficiency, and the price received for the harvested product, with yield expectancy the most impacting factor. In all simulations, NDE was found to be very low, less than 0.48 plants m-2 evidencing the need of herbicides in the control of RR volunteer corn even at low densities.
Keywords:
Glycine max; level of economic damage; volunteer corn density; yield loss
RESUMO:
O milho voluntário é considerado uma planta daninha quando sua emergência ocorrer durante o ciclo da cultura em sucessão, como é o caso de plantas de milho voluntário de híbridos Roundup Ready (RR) em lavouras implantadas com cultivares de soja RR. O objetivo deste estudo foi verificar o efeito da interferência de diferentes densidades de plantas de milho voluntário RR sobre a cultura da soja RR, bem como estimar o nível de dano econômico (NDE). Para isso, foi conduzido um experimento em campo em dois locais em esquema fatorial com quatro repetições; os tratamentos constaram de densidades de milho voluntário RR (0, 1, 2, 3, 4, 5, 6, 7 e 8 plantas por m-2) competindo em todo o ciclo da soja. As alterações na morfologia e nos componentes de rendimento se comportaram de forma diretamente proporcional ao incremento das densidades de milho voluntário, sendo a variável legumes por planta a mais intensamente afetada. A perda de rendimento da soja na maior densidade de competidor chegou a 58%. O NDE foi estreitamente dependente do custo de controle das plantas de milho voluntário, da expectativa de rendimento da cultura, da eficiência do herbicida e do preço recebido pelo produto colhido, sendo a expectativa de rendimento o fator mais impactante. Em todas as simulações, o NDE encontrado foi muito baixo, menor que 0,48 planta m-2, evidenciando a necessidade do uso de herbicidas no controle do milho voluntário RR mesmo em baixas densidades.
Palavras-chave:
Glycine max; nível de dano econômico; densidade de milho voluntário; perda de rendimento
INTRODUCTION
The use of herbicide-resistant crops allows greater flexibility in the control of weeds in the post-emergence period. Thus, the use of resistant crops has grown in Brazil and Latin America. According to the International Service for the Acquisition of Agri-Biothec Applications (ISAAA) annual report, in 2014 alone there was an increase of 5.8 million hectares of herbicide-resistant soybeans grown in Brazil, Argentina, Paraguay and Uruguay (ISAAA, 2014International Service for the Acquisition of Agri-biothec Applications - ISAAA. Global status of commercialized Biotech/GM Crops: 2014. (ISAAA Brief, 49).). With the intensive use of herbicide-resistant crops, problems have arisen with the evolution of resistant weeds, in addition to the occurrence of resistant volunteer plants, mainly characterized by resistance to glyphosate.
In corn and soybean succession, Roundup Ready (RR) volunteer corn has become a problem for post-emergence control in RR soybeans (Marquardt et al., 2012Marquardt PT, Krupde C, Johnson WG. Competition of transgenic volunteer corn with soybean and the effect on western corn rootworm emergence. Weed Sci. 2012;60:193-8.). Volunteer corn was documented as a weed in soybeans prior to the introduction of RR technology (Andersen and Geadelmann 1982Andersen RN, Geadelmann JL. The effect of parentage on the control of volunteer corn (Zea mays) in soybean (Glycine max). Weed Sci. 1982;30:127-31.; Beckett and Stoller, 1988Beckett TH, Stoller EW. Volunteer corn (Zea mays) interference in soybeans (Glycine max). Weed Sci. 1988;36:159-66.). Volunteer corn plants have a great competitive ability, being able to grow and develop above the canopy of soybean plants and compete mainly for light, resulting in yield losses (Page et al., 2010Page ER, Tollenaar M, Lee EA, Lukens L, Swanton CJ. Shade avoidance: an integral component of crop-weed competition. Weed Res. 2010;50:281-8.). The potential for damage to soybean yield by volunteer corn is high compared to other weed plants. The corn, as a competing plant, reduces soybean yield by the order of 10 to 22% at one plant m-2; this can reach up to 40% with higher densities (Marquardt et al., 2012; Alms et al., 2016Alms J, Moechnig M, Vos D, Clay SA. Yield loss and management of volunteer corn in soybean. Weed Technol. 2016;30:254-62.). However, the plants of Euphorbia heterophylla, Brachiaria plantaginea, Ipomoea grandifolia and Senna obtusifolia at one plant m-2 have low potential to cause damage to soybean yield (Voll et al., 2002Voll E, Gazziero DLP, Brighenti AAM, Adegas FS. Competição relativa de espécies de plantas daninhas com dois cultivares de soja. Planta Daninha. 2002;20:17-24.). Similarly, high density of Raphanus sativus (55 plants m-2), it has caused losses of soybean yield of 3 to 15% (Bianchi et al., 2011Bianchi MA, Fleck NG, Agostinetto D, Rizzardi MA. Interferência deRaphanus sativusna produtividade de cultivares de soja. Planta Daninha. 2011;29:783-92.).
One of the main problems that intensify the interference of RR volunteer corn in soybeans is related to the slow initial development of the soybean plant and its reduced size compared to the corn plants. These attributes cause the corn plants to cause intense shading, reducing photosynthetic rate in soybeans and stimulating their stem elongation (Cardoso et al., 2010Cardoso GD, Alves PLCA, Beltrão NEM, Vale LS. Períodos de interferência das plantas daninhas em algodoeiro de fibra colorida ‘BRS Safira’. Rev Cienc Agron. 2010;41:456-62.). In addition to competing for environmental resources, such as water, nutrients and CO2, volunteer corn may in some cases interfere with the mechanical harvesting process and serve as hosts for several insect pests (Marquardt et al., 2012Marquardt PT, Krupde C, Johnson WG. Competition of transgenic volunteer corn with soybean and the effect on western corn rootworm emergence. Weed Sci. 2012;60:193-8.).
Estimation of crop yield losses due to weed interference and determination of an economic threshold are important components for decision making in the adoption of an integrated weed management strategy. Economic models based on the economic threshold (ET) help in the decision making process in terms of methods of control for certain weed populations. Thus, the ET is characterized by the use of control methods on a plant population when the damage caused to the grain yield is higher than the cost of the control measures to be adopted (Pester et al., 2000Pester TA, Westra P, Anderson RL, Lyon DJ, Miller SD, Sthalman PW, Northan FE, Wicks GA. Secale cereal interference and economic thresholds in winter Triticum aestivum. Weed Sci. 2000;48:720-7.). The ET takes into consideration the relationship between weed density, expected crop yield, crop price, and control efficiency. In order to establish an ET, it is necessary to take into account variables that are dependent on environmental characteristics, crop management, prices of food and other external factors (Berti et al., 1997Berti A, Zanin G. A decision model for post-emergence weed management in soybean (Glycine max (L.) Merr.). Crop Prot. 1997;16:109-16.).
Information on interference and the ET from volunteer corn plants in soybean crop are scarce. When available, the information based on the ET is important in supporting measures for volunteer corn management, providing a more economical and friendly system of crop production.
The hypothesis of the study is that the loss of soybean yield caused by the interference of volunteer corn varies with the density of the weeds and can be quantified by the use of mathematical models that enable levels of damage in the crop to be established. Therefore, the objective of the study was to verify the effects of interference of RR volunteer corn densities on RR soybean, as well as to estimate the economic threshold level.
MATERIAL AND METHODS
Fields researches were conducted at two locations in the crop season 2014/2015. The first site was in the experimental field of the Federal University of Santa Maria, campus of Frederico Westphalen; the second site was a commercial field near Frederico Westphalen, RS. Soil at these locations was a typical dystrophic red latosol. The climate in the locations where the experiments were conducted, according to Köppen’s Climate classification, is humid subtropical climate (CFA). The soil physical and chemical characteristics are showed in Table 1.
The experimental design was a randomized block design, with a 2 x 7 factorial scheme and four replicates. The treatments consisted of two sites and RR volunteer corn densities of 0, 1, 2, 3, 4, 5, 6, 7 and 8 plants m-2. Each experimental plot contained eight rows of plants, 4.0 m long, with row spacing of 0.45 m. The soybean crop was grown in succession to corn. Experimental sites were kept free of weeds other than volunteer corn by application pre-planting of glyphosate (900 g ha-1) and paraquat (400 g ha-1) in sequential application with a seven-day interval. The soybean seeds were inoculated with Bradyrhizobium japonicum, in the proportion of 0.5 kg of inoculant for each 60 kg of seeds. A glyphosate-resistant soybean cultivar (NA 5909 RR) was planted manually on December 19, 2014 at 288.888 seeds ha-1. Volunteer corn F2 was planted in soybean narrow row adjusted according to the respective volunteer corn densities. After planting, irrigation was performed equivalent to 15 mm of rainfall. Figure 1 presents data for rainfall (mm) and maximum temperatures during the crop season. The emergence of soybean and volunteer corn occurred at eight days after planting.
The post-emergence application of herbicides was performed with glyphosate (900 g ha-1) at the V2 and V6 soybean growth stages. Lufenuron and Tiametoxam + Lambda-Cialotrina were applied as necessary according to recommendations for insect control. Control of soybean Asiatic rust involved application of Trifloxistrobina + Protioconazol from the R1 soybean growth stage and then every 21 days. All pesticide treatments were applied using a back-pack sprayer fitted with a 80015 conical nozzle and calibrated to deliver 200 L ha-1.
At maturity, 10 arbitrarily selected plants was used for measurement of soybean height, stem diameter, and number of branches, nodes and pods plant-1. Soybean grain yield was determined by manual harvesting of two central rows from each plot; the grains were adjusted to 13% moisture content and yield expressed as kg ha-1.
The soybean yield data were transformed into a yield loss (YL%) in relation to volunteer corn-free (Equation 1):
where: R1 = yield of grain from volunteer corn-free plots; R2 = yield of grain at each density of volunteer corn.
The data were submitted to analysis of variance using the F-test (p<0.05), and when the significant was adjusted to the linear or nonlinear regression model between independent and dependent variables. The soybean yield was adjusted to the nonlinear regression model of the rectangular hyperbola proposed by Cousens (1985Cousens R. An empirical model relating crop yield to weed and crop density and a statistical comparison with other models. J Agric Sci. 1985;105:513-21.) (Equation 2), which describes the behavior of the grain yield as a function of the level of volunteer corn:
where: YL = yield loss relative to volunteer corn-free plots; D = volunteer corn density; I= yield loss per unit of volunteer corn when the density approached zero; a =maximum yield loss with the increase of volunteer corn density tending to infinity.
The adjustment of the data to the model was performed by the PROC NLIN procedure of the Statistical Analysis System (SAS) computer program, and the graphs were constructed using SigmaPlot 10. To calculate the ET, we used the estimates of parameter i obtained from the equation cited before and Equation 3, adapted from Lindquist and Kropff (1996Lindquist JL, Kropff MJ. Application of an ecophysiological model for irrigated rice (Oryza sativa) - Echinochloa competition. Weed Sci. 1996;44:52-56.):
In which: ET = economic threshold (plants ha-1); WC = weed control cost (herbicide and application, in US$ ha-1); Y = soybean grain yield (kg ha-1); P = soybean price (US$ bag-1); i= soybean loss yield (%) per volunteer corn unit when the density level approaches zero and H = herbicide efficiency (%).
For ET calculation purposes, three values were estimated for soybean yield potential, soybean price, control cost and herbicide efficiency in volunteer corn. The crop yield range was estimated as 2000, 3500 and 5000 kg ha-1, which are expected soybean yields in the state of Rio Grande do Sul (CONAB, 2016Campanha Nacional de Abastecimento - Conab. Acompanhamento da safra brasileira de grãos. Safra 2015/16 - Quarto levantamento. Brasília, DF: 2016. v.4. 154p.). The minimum, average and maximum prices offered for a 60 kg bag of soybean in the last five years in the average of Rio Grande do Sul were US$ 16.4, 21.8 and 27.3 bag-1, respectively (CONAB, 2016Campanha Nacional de Abastecimento - Conab. Acompanhamento da safra brasileira de grãos. Safra 2015/16 - Quarto levantamento. Brasília, DF: 2016. v.4. 154p.). The cost of control took into account the average price of herbicides recommended for post-emergence management and the cost of application (tractor + sprayer); this was US$ 30.3, 36.4 and 42.4 ha-1. For herbicide efficiency in the control of volunteer corn, the range of values consisted of 80, 90 and 100%.
RESULTS AND DISCUSSION
There was a significant interaction between location and volunteer corn density to soybean stem diameter. For the others variables, there was no local effect, only an effect of volunteer corn density.
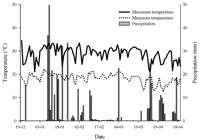
The volunteer corn densities caused changes in the height and stem diameter of the soybean (Figure 2). Soybean height was correlated to increasing volunteer corn density, and there was an estimated increase of 1 cm for each corn plant added (Figure 2A). The increase in soybean height is outcome of the light interference by corn, as a strategy to avoid future competition, and its result in alteration of physiological processes related to hormonal dynamics and cell division, reducing the crops ability to absorb water and soil nutrients required to perform photosynthesis (Jensen et al.,1998Jensen PJ, Hangarter RP, Mark E. Auxin transport is required for hypocotyl elongation in light-growth but not dark-growth Arabdopsis. Plant Physiol. 1998;116:455-62.).
The soybean height (A) and reduction of the soybean stem diameter (B) as a function of the volunteer corn density. Frederico Westphalen, RS, Brazil, 2014/15.
The greatest height of the soybean plants with increasing volunteer corn density can be attributed to the weed community, and the changes caused by weeds in light quality help the crop detect the proximity of neighboring, which impacts on the increase of the plant height as a way of capturing the maximum available radiation and shading weeds (Rajcan and Swanton, 2001Rajcan I, Swanton CJ. Understanding maize-weed competition: resource competition, light quality and the whole plant. Field Crops Res. 2001;71:139-50.). The red/far-red ratio perceived by phytochromes plays an important role in the induction of many morphological alterations in plant architecture, such as an increase in the proportion of stem accumulation (Ballare and Casal, 2000Ballare CL, Casal JJ. Light signals perceived by crop and weed plants. Fields Crops Res. 2000;67:149-60.). The shading caused by weeds changes the patterns of photoassimilate allocation for investment in shoot growth, will result on fitness cost for below-ground resource competition, like water and soil nutrients (Rajcan and Swanton, 2001).
The reduction in soybean stem diameter was proportional to the increase in weed plant density (Figure 2B). At site 1, there was a greater effect of volunteer corn densities, where the reduction was 10.2 and 52.7% with increase in corn density from 1 to 8 plants m-2, respectively. However, the reduction in soybean stem diameter at site 2 was 11 and 49.5%, respectively, with the same increase in weed plant density. The reduction of stem diameter is directly linked to increase of height plant. The elongation stem causes a decrease in stem diameter of the crop, since weeds induce a change in the light quality in the lower canopy, causing a reduction in the stem thickness development (Crotser et al., 2003Crotser MP, Witt WW, Spomer LA. Neutral density shading and far-red radiation influence black nightshade (Solanum nigrum) and eastern black nightshade (Solanum ptycanthum) growth. Weed Sci. 2003;51(2):208-13.).
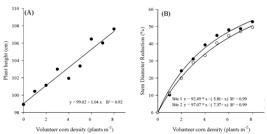
The interference of volunteer corn also reduced the number of nodes on the main stem (Figure 3A). The nodes was reduced to 20, 30 and 48% when volunteer corn density varied from 2, 4 and 8 plants m-2, respectively, competing throughout the soybean cycle. Similarly, the number of lateral branches was reduced by 15, 41 and 60% when the competing corn densities were 2, 4 and 8 plants m-2, respectively (Figure 3B).
Reduction of the soybean number of nodes in the main stem (A) and the number of lateral branches (B) as a function of the volunteer corn density. Frederico Westphalen, RS, Brazil, 2014/15.
In conditions of high competition, there is less availability of photoassimilates for branch growth, since they are preferentially destined for main stem growth (Martins et al., 1999Martins MC, Câmara GMS, Peixoto CP, Marchiori LFS, Leonardo V, Mattiazzi P. Épocas de semeadura, densidades de plantas e desempenho vegetativo de cultivares de soja. Sci Agríc. 1999;56:851-8.). In the same way, treatments with lower plant densities provided a faster growth of the crop, therefore, it can be inferred that weed interference inhibits lateral branch growth in soybean plants (Carvalho and Veline, 2001Carvalho FT, Velini ED. Períodos de interferência de plantas daninhas na cultura da soja. I - Cultivar IAC-11. Planta Daninha. 2001;19:317-22.). The redution in the number of soybean branches was noted with Bidensspp. and Sida rhombifolia interference in soybean, which negatively impacted mainly on the number of lateral branches on the soybean plant (Fleck et al., 2004Fleck NG, Rizzardi MA, Agostinetto D, Balbinot Junior AA. Interferência de picão-preto e guanxuma com a soja: efeitos da densidade de plantas e época relativa de emergência. Cienc Rural. 2004;34:41-8.).
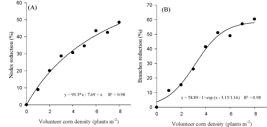
The number of pods per plant was the component most severely affected by the increase in volunteer corn densities (Figure 4A). Corn interference caused a 24 to 74% reduction in the number of pods when volunteer corn densities increased from 1 to 8 plants m-2, respectively. These results are consistent with those reported by Silva et al., (2008Silva AF, Ferreira EA, Concenço G, Ferreira FA, Aspiazu I, Galon L et al. Densidades de plantas daninhas e épocas de controle sobre os componentes de produção da soja. Planta Daninha. 2008;26:65-71.), who found that number of soybean pods was the variable that was most affected by low, medium and high infestation of the weed community, reducing the number of pods per plant by 58 to 78%.

Reduction of the soybean number of pods per plant (A) and soybean yield loss (B) as a function of the volunteer corn density. Frederico Westphalen, RS, Brazil, 2014/15.
The most important variable that was affected by weed interference is crop grain yield. Volunteer corn plants, even at low densities, are capable of altering the development of soybean by reducing their yield. The reduction in soybean yield in competition with volunteer corn plants ranged from 11 to 58% in the range of plant densities used (Figure 4B).
From the regression analysis by the hyperbola model, it was possible to estimate the parameters that enabled comparing the interference of the species on the crops (Rizzardi et al., 2003Rizzardi MA, Fleck NG, Mundstock CM, Bianchi MA. Perdas de rendimento de grãos de soja causadas por interferência de picão-preto e guanxuma. Cienc Rural. 2003;33:621-7.). The model incorporates the parameteri, which represents the loss of yield caused by the addition of the first weed. For the interference of volunteer corn, the ivalue was 15.4, and the maximum yield loss (parametera) was estimated as 58.9%.
In soybeans, it was estimated that the yield loss caused by the addition of each Bidensspp. and Sida rhombifolia plant was 1.58 and 0.69%, respectively (Rizzardi et al., 2003Rizzardi MA, Fleck NG, Mundstock CM, Bianchi MA. Perdas de rendimento de grãos de soja causadas por interferência de picão-preto e guanxuma. Cienc Rural. 2003;33:621-7.), indicating an extremely low value when compared to i values of volunteer corn (Figure 4B). Reports have verified that volunteer corn causes yield losses in soybean of 10 and 41% with 0.5 and 16 corn plants m-2, respectively, and showed an i value of approximately 25% and a value of 46%, (Marquardt et al., 2012Marquardt PT, Krupde C, Johnson WG. Competition of transgenic volunteer corn with soybean and the effect on western corn rootworm emergence. Weed Sci. 2012;60:193-8.). In a previous study, 0.3 volunteer corn plants m-2 reduced soybean yields by 9%, increasing to 51% with 4.4 plants m-2, with an i value of 40 and a value of 70% (Alms et al., 2016Alms J, Moechnig M, Vos D, Clay SA. Yield loss and management of volunteer corn in soybean. Weed Technol. 2016;30:254-62.). This indicated the high competitive ability of volunteer corn which may make the crop more susceptible to its interference.
The yield of soybean in weed-free plots was 3984 kg ha-1, and the loss estimated at the i value corresponded to 614 kg ha-1. Taking into account the price of a 60 kg bag of soybean of US$ 21.8 bag-1, this equates to a monetary loss of US$ 223 ha-1 for each volunteer corn plant m-2. Assuming the price of the product to be from US$ 27.3 to 16.4 bag-1, the loss in monetary value varied from US$ 279 to 167 ha-1, respectively, for each volunteer corn m-2 according to the ivalue.
In general, the ET values for volunteer corn are low, which indicates the aggressiveness of the volunteer corn in reducing crop yield. The cost of weed control is a relevant factor in determining the level of ET (Figure 5). Any increase in the herbicide costs or its application means increases in the ET. Increasing weed control costs from US$ 30.3 to 42.4 ha-1, increases the ET from 913 to 2737 volunteer corn plant ha-1 (0.930 to 0.2737 plants m-2), respectively, based on an average soybean yield of the 3500 kg ha-1. On the other hand, when soybean yields are reduced and weed control costs are high, the ET values increase to 4800 plants ha-1 (0.4800 plants m-2).

Economic thresholds (ET) of volunteer corn for soybean, according the simulations of expected crop yield, control cost (A), soybean price (B) and herbicide efficiency (C). Frederico Westphalen, RS, Brazil, 2014/2015.
Effectiveness of herbicide treatment depends on a number of factors related to herbicides, weeds and climatic conditions. Thus, the choice of the active ingredient, dose, plant size, and soil and air humidity conditions at the time of application may result in high levels of control when these variables are favorable, or, where any of these factors are limiting, control may be less effective and may not totally prevent interference with the crop.
The price of the soybean crop is another factor that affects decision making in the control of volunteer corn (Figure 5B). Three different prices paid for soybean grain (US$ 16.4, 21.8 and 27.3 bag-1) were estimated in this study. When the high price are offers for soybean, the ET of the volunteer corn is reduced. When the price of soybean increased from US$ 16.4 to 27.3 bag-1, the ET reduced from 2756 to 1372 volunteer corn plants ha-1 (0.2756 to 0.1372 plants m-2) based on average values of crop yield. In the same sense, when the price offered for the product rises and the expected crop yield increases, the ET of the volunteer corn is even lower.
Assuming the receipt of US$ 27.3 bag-1 soybean and the expectation of a high yield of 5000 kg ha-1, the ET presented an extremely low value of 959 ha-1 volunteer corn, emphasizing the high capacity for interference in soybean crop and the need for control of volunteer corn, even at low densities.
The selection of herbicides with lower efficiency indicates an increase in ET (Figure 5C). The recommended doses for herbicides are usually defined in order to achieve an effective control under a wide range of environmental and management conditions, but sometimes inadequate weed management or adverse environmental conditions can reduce herbicide efficiency. Three levels of herbicide efficiency (80, 90 and 100%) were considered in the study. Considering an efficiency of 80%, the ET for the volunteer corn was 2044 plants ha-1 (0.2044 plants m-2). However, with 100% control, the ET dropped to 1640 plants ha-1 (0.1640 plants m-2), based on average values for crop yield of 3500 kg ha-1. Although low densities of competing plants may not significantly affect the final grain yield of the crop, harvesting delays and problems with the harvesting equipment itself are likely to occur (Czapar et al., 1997Czapar GF, Curry MP, Wax LM. Grower acceptance of economic thresholds for weed management in Illinois. Weed Technol. 1997;11:828-31.).
Regardless of the simulations, the ET of volunteer corn in soybean was very low (<0.48 plants m-2). This was interpreted as indicative of the need to control this earlier volunteer plant due to the high risk of yield losses. Some herbicides are indicated for the control of volunteer corn, where soybean yield was ensured by effectively controlling glyphosate-tolerant volunteer corn with a single application of clethodim (108 g ha-1) at stage V6 of soybean growth, or sequential applications at V3 and V6 growth stages (López Ovejero et al., 2016López-Ovejero RF, Soares DJ, Oliveira NC, Kawaguchi IT, Berger GU, Carvalho SJP, Christoffoleti PJ. Interferência e controle de milho voluntário tolerante ao glifosato na cultura da soja. Pesq Agropec Bras. 2016;51:350-7. ).
It is concluded that glyphosate-resistant volunteer corn adversely affects the soybean grain yield proportionally to the increase in plant density, with the number of pods per plant being the variable most affected by the interference. With the determination of the ET, it can be concluded that with increases in soybean grain yield, soybean prices and herbicide efficiency, there is a reduction in ET. In contrast, increase in the costs of weed control raise the ET. In all simulations, ET values were very low, less than 0.48 plants m-2, demonstrating the need for herbicide control of volunteer corn, even at low densities.
REFERENCES
- Alms J, Moechnig M, Vos D, Clay SA. Yield loss and management of volunteer corn in soybean. Weed Technol. 2016;30:254-62.
- Andersen RN, Geadelmann JL. The effect of parentage on the control of volunteer corn (Zea mays) in soybean (Glycine max). Weed Sci. 1982;30:127-31.
- Ballare CL, Casal JJ. Light signals perceived by crop and weed plants. Fields Crops Res. 2000;67:149-60.
- Beckett TH, Stoller EW. Volunteer corn (Zea mays) interference in soybeans (Glycine max). Weed Sci. 1988;36:159-66.
- Berti A, Zanin G. A decision model for post-emergence weed management in soybean (Glycine max (L.) Merr.). Crop Prot. 1997;16:109-16.
- Bianchi MA, Fleck NG, Agostinetto D, Rizzardi MA. Interferência deRaphanus sativusna produtividade de cultivares de soja. Planta Daninha. 2011;29:783-92.
- Campanha Nacional de Abastecimento - Conab. Acompanhamento da safra brasileira de grãos. Safra 2015/16 - Quarto levantamento. Brasília, DF: 2016. v.4. 154p.
- Cardoso GD, Alves PLCA, Beltrão NEM, Vale LS. Períodos de interferência das plantas daninhas em algodoeiro de fibra colorida ‘BRS Safira’. Rev Cienc Agron. 2010;41:456-62.
- Carvalho FT, Velini ED. Períodos de interferência de plantas daninhas na cultura da soja. I - Cultivar IAC-11. Planta Daninha. 2001;19:317-22.
- Cousens R. An empirical model relating crop yield to weed and crop density and a statistical comparison with other models. J Agric Sci. 1985;105:513-21.
- Crotser MP, Witt WW, Spomer LA. Neutral density shading and far-red radiation influence black nightshade (Solanum nigrum) and eastern black nightshade (Solanum ptycanthum) growth. Weed Sci. 2003;51(2):208-13.
- Czapar GF, Curry MP, Wax LM. Grower acceptance of economic thresholds for weed management in Illinois. Weed Technol. 1997;11:828-31.
- Fleck NG, Rizzardi MA, Agostinetto D, Balbinot Junior AA. Interferência de picão-preto e guanxuma com a soja: efeitos da densidade de plantas e época relativa de emergência. Cienc Rural. 2004;34:41-8.
- International Service for the Acquisition of Agri-biothec Applications - ISAAA. Global status of commercialized Biotech/GM Crops: 2014. (ISAAA Brief, 49).
- Jensen PJ, Hangarter RP, Mark E. Auxin transport is required for hypocotyl elongation in light-growth but not dark-growth Arabdopsis. Plant Physiol. 1998;116:455-62.
- Lindquist JL, Kropff MJ. Application of an ecophysiological model for irrigated rice (Oryza sativa) - Echinochloa competition. Weed Sci. 1996;44:52-56.
- López-Ovejero RF, Soares DJ, Oliveira NC, Kawaguchi IT, Berger GU, Carvalho SJP, Christoffoleti PJ. Interferência e controle de milho voluntário tolerante ao glifosato na cultura da soja. Pesq Agropec Bras. 2016;51:350-7.
- Marquardt PT, Krupde C, Johnson WG. Competition of transgenic volunteer corn with soybean and the effect on western corn rootworm emergence. Weed Sci. 2012;60:193-8.
- Martins MC, Câmara GMS, Peixoto CP, Marchiori LFS, Leonardo V, Mattiazzi P. Épocas de semeadura, densidades de plantas e desempenho vegetativo de cultivares de soja. Sci Agríc. 1999;56:851-8.
- Page ER, Tollenaar M, Lee EA, Lukens L, Swanton CJ. Shade avoidance: an integral component of crop-weed competition. Weed Res. 2010;50:281-8.
- Pester TA, Westra P, Anderson RL, Lyon DJ, Miller SD, Sthalman PW, Northan FE, Wicks GA. Secale cereal interference and economic thresholds in winter Triticum aestivum Weed Sci. 2000;48:720-7.
- Rajcan I, Swanton CJ. Understanding maize-weed competition: resource competition, light quality and the whole plant. Field Crops Res. 2001;71:139-50.
- Rizzardi MA, Fleck NG, Mundstock CM, Bianchi MA. Perdas de rendimento de grãos de soja causadas por interferência de picão-preto e guanxuma. Cienc Rural. 2003;33:621-7.
- Silva AF, Ferreira EA, Concenço G, Ferreira FA, Aspiazu I, Galon L et al. Densidades de plantas daninhas e épocas de controle sobre os componentes de produção da soja. Planta Daninha. 2008;26:65-71.
- Voll E, Gazziero DLP, Brighenti AAM, Adegas FS. Competição relativa de espécies de plantas daninhas com dois cultivares de soja. Planta Daninha. 2002;20:17-24.
Publication Dates
-
Publication in this collection
2018
History
-
Received
23 May 2017 -
Accepted
13 July 2017





