
ABSTRACT:
Ivy-leaved morning-glory (Ipomoea hederacea Jacq.) is an exotic species that is becoming an increasing problem in soybean fields of Golestan province, Iran. Because little information is available on the biology of this weed species in Iran, experiments were conducted to investigate the effects of different factors on seed germination and emergence of Ivy-leaved morning-glory. Maximum germination occurred at constant temperature of 20 oC (82%) and alternating temperature of 15/25 oC (94%). Germination was reduced with increasing salinity and drought stresses. Sodium chloride concentration and osmotic potential that inhibited 50% maximum germination were -1.64 and -1.03 MPa, respectively. Acidity was a limiting factor for the germination, due to inhibiting effect of alkaline conditions on germination. However, the results showed that high temperature pretreatment >100 oC decreased seed germination. Germination decreased from 82 to 3% as temperature increased from 100 to 130 oC. There was no significant difference between seedling emergence in burial depths of 1 to 10 cm, but emergence reduced with increasing burial depth from 10 to 14 cm, and no seedling was emerged from a depth of 15 cm. The results of the flooding experiment also revealed that the seeds of this species are sensitive to this stress, so that emergence was 9% after 3 d flooding.
Keywords:
biotic stress; alternating temperatures; exotic species; pH
RESUMO:
Ipomoea hederaceae Jacq. é uma espécie exótica que está se tornando um problema crescente em lavouras de soja da província de Golestan, Irã. Considerando que há pouca informação disponível sobre a biologia dessa espécie no Irã, experimentos foram conduzidos para investigar os efeitos de diferentes fatores sobre a germinação de sementes e emergência de Ipomoea hederaceae. A máxima germinação ocorreu em temperatura constante de 20 oC (82%) e em temperatura alternada de 15/25 oC (94%). A germinação foi reduzida com o aumento da salinidade e do estresse hídrico. A concentração de cloreto de sódio e o potencial osmótico que inibiram 50% da máxima germinação foram de -1,64 e -1,03 Mpa, respectivamente. O pH foi um fator limitante para a germinação, pois houve efeitos inibitórios das condições alcalinas sobre a germinação. Os resultados demonstraram que o pré-tratamento das sementes com temperatura elevada, maior que 100 oC, resultou em decréscimo da germinação. A germinação decresceu de 82% para 3% com o aumento da temperatura de 100 oC para 130 oC. Não houve diferenças de emergência de plântulas em profundidades de enterrio entre 1 e 10 cm, mas a emergência foi reduzida com o aumento da profundidade de enterrio de 10 para 14 cm, e nenhuma plântula emergiu a 15 cm de profundidade. Os resultados de experimentos com inundação também revelaram que as sementes dessa espécie são sensíveis a esse estresse, tanto que a emergência após três dias de inundação foi de 9%.
Palavras-chave:
estresse abiótico; temperaturas alternadas; espécies exóticas; pH
INTRODUCTION
Ivy-leaved morning-glory (Ipomoea hederacea Jacq.) belongs to Convolvulaceae family. It is a summer climbing vine with alternate, ivy-shaped leaves and erect hairs on both surfaces of leaves. Flowers are large, showy, and white to pink to dark purple (Tenaglia, 2017Tenaglia D. Photographs and descriptions of the flowering and non-flowering plants of Missouri [acesso em: 10 nov 2017]. Disponível em: Disponível em: http://www.missouriplants.com/
.
http://www.missouriplants.com/...
). This plant is native to tropical America (Anonymous, 2018Anonymous. Ipomoea hederacea (Convolvulaceae): the global compendium of weeds [acesso em: 10 mar 2018]. Disponível em: Disponível em: http://www.hear.org/gcw/html/autogend/species/10267.HTM
.
http://www.hear.org/gcw/html/autogend/sp...
). In some Asian countries such as India and Pakistan, its seeds are used for the medicinal properties (Singh and Singh, 2012Singh BP, Singh S. Chemical investigation of seed of Ipomoea hederacea and its biological activity. J Chem Pharm Res. 2012;4(2):1441-8.; Ul-Haq et al., 2012Ul-Haq MZ, De-Feo V, Riaz M. Ipomea hederaceaJacq: a medicinal herb with promising health benefits. Molecules. 2012;17(11):13132-45.). But this species is known as an important weed in subtropical areas. It is considered as one of 10 worst weeds in soybean, cotton, and tobacco in the southeast of USA. This weed is also observed in Africa (Ghana), South America (Brazil), and it is reported as a quarantine weed from Australia. As well as, it was reported in 2010 for the first time from Soybean fields of Aliabad region, Gorgan, Golestan province in Iran (Pahlevani and Sajedi, 2011Pahlevani AH, Sajedi S. Alerting occurrence of several noxious weeds and invasive plants in arable lands in Iran. Rostaniha. 2011;12 (2):129-34. ).
Ipomea spp. are soybean competitive weed, as Culpepper et al. (2001Culpepper AS, Gimenez AE, York AC, Batts RB, Wilcut JW. Morning-glory (Ipomoea spp.) and large crabgrass (Digitaria sanguinalis) control with glyphosate and 2,4-DB mixtures in glyphosate-resistant soybean (Glycine max). Weed Technol. 2001;15(1):56-61. ) stated that these weeds can reduce soybean yield 25-43% at densities of 2 to 8 plants m-2. Furthermore, various species of morning-glory may result in the crop lodging, interfere with harvesting operations that reduce harvesting efficiency, increase foreign matter in the harvested product and reduce product quality. Since the primary introduction of weeds to new areas takes place mainly through seeds, understanding the seed ecology is required. Also, studying the ecology of seed germination of various species is very useful to recognize and explain plant evolution and ecological adaptation (Baskin et al., 2004Baskin CC, Milberg P, Andersson L, Baskin JM. Germination ecology of seeds of the annual weeds Capsella bursa-pastoris and Descurainia sophia originating from high northern latitudes. Weed Res. 2004;44(2):60-8.). Biological and ecological information, specifically germination ecology of a specific weeds is necessary to develop long-term management strategies (Mennan and Ngouajio, 2006Mennan H, Ngouajio M. Seasonal cycles in germination and seedling emergence of summer and winter population of catch weed bedstraw and wild mustard. Weed Sci. 2006;54(1):114-20. ; Nakamura and Hossain, 2009Nakamura I, Hossain MA. Factors affecting the seed germination and seedling emergence of redflower ragleaf (Crassocephalum crepidioides). Weed Biol Manag. 2009;9(4):315-22.; Ali et al., 2013Ali HH, Tanveer A, Nadeem MA, Asghar HN, Javid MM. Germination ecology of Rhynchosia capitata: an emerging summer weed in Asia. Planta Daninha. 2013;31(2):249-57.; Tanveer et al., 2014Tanveer A, Sibtain M, Javaid MM, Ali HH. Germination ecology of wild onion: a rainfed crop weed. Planta Daninha. 2014;32(1):69-80. ). Peters et al. (2000Peters NCB, Atkins HA, Brain P. Evidence of differences in seed dormancy among populations of Bromus sterilis. Weed Res. 2000;40(5):457-78.) reported that knowledge of the influence of environmental factors on the germination and emergence behavior of seeds would be helpful to prevent their invasion to new areas and determine its management practices.
Germination of weed seed influenced by many environmental factors, such as the temperature, pH, soil salinity and moisture (Koger et al., 2004Koger CH, Reddy KN, Ppston DH. Factors affecting seed germination, seedling emergence, and survival of texas weed (Caperonia palustris). Weed Sci. 2004;52(6):989-95. ; Chauhan et al., 2006aChauhan BS, Gill G, Preston C. African mustard (Brassica tournefortii) germination in southern Australia. Weed Sci. 2006a;54(5):891-7., bChauhan BS, Gill G, Preston C. Influence of environmental factors on seed germination and seedling emergence of Oriental mustard (Sisymbrium orientale). Weed Sci. 2006b;4(6):1025-31. ; Eslami et al., 2010Eslami SV, Afghani F, Mahmoodi S. Effects of some environmental factors on germination and longevity of downy brome (Bromus tectorum) seeds. Iran J Weed Sci. 2010;4(2):47-57. ; Altop et al., 2015Altop EK, Mennan H, Philippo CJ, Zandstra BH. Effect of the burial depth and environmental factors on the seasonal germination of bearded sprangletop (Leptochloa fusca [L.] Kunth ssp. fascicularis [Lam.] N. Snow). Weed Biol Manag. 2015;14(4):147-58.; Asgarpour et al., 2015Asgarpour R, Ghorbani R, Khajeh-hosseini M, Mohamadvand E. Germination of spotted spurge (Chamaesyce maculata) seeds in response to different environmental factors. Weed Sci. 2015;63(2):502-10.; Sadeghloo et al., 2013Sadeghloo A, Asghari J, Ghaderi-Far F. Seed germination and seedling emergence of velvetleaf (Abutilon theophrasti) and barnyardgrass (Echinochloa crus-galli). Planta daninha. 2013;31(2):259-66.). Seed germination under adverse environmental conditions such as drought and salinity stresses might enable weeds to take advantage of the limiting conditions of the growth of other species (Chauhan and Johnson, 2008Chauhan BS, Johnson DE. Germination ecology of southern crabgrass (Digitaria ciliaris) and India crabgrass (Digitaria longiflora): two important weeds of rice in tropics. Weed Sci. 2008;56(5):722-8. ).
Because of the increasing range of this weed distribution in soybean fields in Golestan province, we investigated different environmental factors including of temperature (constant, alternating and high temperatures), osmotic and salinity stresses, pH, burial depth and flooding duration on seed germination and seedling emergence of Ivy-leaved morning-glory.
MATERIALS AND METHODS
Experiments were conducted to evaluate the effect of constant temperature, alternating temperature, pH, salinity stress, drought stress, and high temperature pretreatment on germination, and the influence of planting depth and flooding on emergence of Ivy-leaved morning-glory.
Seed collection
Seeds collected from multiple plants through soybean fields of Gorgan (36o50’29” N, 54o26’36” E, 143 m above sea level) in fall 2013. They stored at 4 oC until experiment initiation. The mean 100 seed weight was 2.93 g. A preliminary test of germination indicated that seeds had dormancy. So, treatment of concentrated sulphuric acid (98% H2SO4) for 25 min was used to break seed dormancy.
General protocol for germination tests
Four replications of 25 seeds of Ivy-leaved morning-glory were placed in 9 cm petri dishes on a layer of Whatman No. 1 filter paper, moistened with either 5 mL distilled water or treatment solution when required. Germination tests were conducted in an incubator at 25 oC in darkness (optimum temperature for germination, data not shown), except for the experiment of the effect of temperature on seed germination. The seeds were checked daily for germination. Seeds were considered germinated when the radicle was 2 mm in length (Ghaderi-Far et al., 2010Ghaderi-Far F, Gherekhloo J, Alimagham SM. Influence of environmental factors on seed germination and seedling emergence of yellow sweet clover (Melilotus officinalis). Planta Daninha. 2010;28(3):436-69.; Maraghni et al., 2010Maraghni M, Gorai M, Naffati M. Seed germination at different temperatures and water stress levels, and seedling emergence from different depths of Ziziphus lotus. S Afr J Bot. 2010;76(3):453-9. ; Sing et al., 2012). Seed germination was counted until 14 d after the start of the experiment.
Effect of constant and alternating temperatures on germination
Seven constant temperatures (10, 15, 20, 25, 30, 35 and 40 oC) and eight fluctuating 12/12 h night/day temperatures (7.5/12.5, 10/15, 12.5/20, 15/25, 20/30, 25/37.5, 30/42.5, and 35/45 oC) in darkness were selected to determine the effect of temperature on germination.
Effect of high temperature pretreatment on germination
Seeds were subjected to high temperature for short duration to simulate the fire effects. Seven various temperatures (80, 100, 120, 130, 160, 200, and 240 oC) for 5 min were studied along with a control (no treatment). A drying oven was utilized for high temperature treatment of seeds. Each treatment consisted of four replicate petri dishes with 25 seeds. Seeds were then set to germinate using the optimum temperature of 25 oC.
Effect of osmotic and salinity stresses on germination
Osmotic solutions with 0, -0.2, -0.4, -0.6, -0.8, -1.0, -1.1, -1.2, and -1.4 MPa potentials were prepared by dissolving measured quantities of PEG 8000 in distilled water (Michel, 1983).
Salt stress on seed germination was evaluated by placing the seeds in dishes containing sodium chloride (NaCl) solutions of 0, -0.2, -0.4, -0.6, -0.8, -1.0, -1.1, -1.2, -1.4, -1.5, -1.8, and -2.0 MPa, which were prepared by dissolving 0, 2.35 , 4.70, 7.06, 9.41, 11.77, 14.12, 16.47, 18.83, 21.18 and 23.54 g NaCl in 1 L of distilled water.
Effect of pH on germination
The effect of pH on seed germination was evaluated by placing seeds in buffer solutions with pH 3 to 9 prepared according to Susko et al. (1999Susko DJ, Mueller JP, Spears JF. Influence of environmental factors on germination and emergence of Pueraria lobata. Weed Sci. 1999;47(5):585-8. ). Distilled water (pH=6.6) was used as the control.
Effect of burial depth on seedling emergence
Twenty-five seeds of Ivy-leaved morning-glory were buried in soil in 19 cm diam plastic pots at depths of 1, 3, 5, 7, 10, 13, 14, and 15 cm. The soil used was loam with an organic matter content of 0.95% and pH of 7.2. Pots were placed in outdoor from June 26 to July 16, 2015. The average minimum and maximum temperatures and the rainfall during the experiment were 23.5 oC, 35.5 oC, and 39.5 mm, respectively. Pots were irrigated throughout the study to provide optimal moisture for seed germination. Seedling emergence was recorded daily until the seedlings ceased to emerge about 20 d after planting. Emergence was defined as the appearance of a part of seedling on the soil surface.
Effect of flooding on seedling emergence
Twenty-five seeds were buried at 1 cm soil depth in each pot (16 cm diam. by 19 cm depth). According to the results of seed depth experiment, maximum germination was observed in seeds buried at this depth. The pots were filled with the same soil as in the burial depth experiment. Pots containing the seeds were placed inside larger plastic pots (20 cm diam. by 32 cm depth) to maintain 3 cm water above the soil.
Statistical Analysis
In all experiments, treatments were laid out in a completely randomized design with four replications. The germination percentage data were transformed prior to analysis to prevent variance heterogeneity, but this did not improve homogeneity. Therefore, non-transformed percentage germination data were applied for ANOVA and regression analysis. Data were subjected to analysis of variance and the treatment means were separated using LSD at P≤0.01. All the statistical analyses were conducted with SAS (version 9.1).
Regression analysis was utilized to determine the effect of osmotic and salinity stresses, planting depth, and high temperature pretreatment. Data were fitted to a three-parameter sigmoidal function sing Sigma Plot (version 11). The model fitted was as follows:
where G is the total germination (%) at different concentrations of salt and osmotic potential, high temperature, or burial depth; Gmax is the maximum germination (%), x50 is the concentration of salt and osmotic potential, high temperature or planting depth required for 50% inhibition of the maximum germination, and Grate indicates the slope.
RESULTS AND DISCUSSION
Effect of constant and alternating temperatures on germination
Germination percentage changes at different temperatures were illustrated in Figure 1. No germination occurred at 10 oC. Fifty-eight percent of seeds germinated at 15 oC. With increasing the temperature from 15 to 20 oC, germination increased, so that the highest germination (82%) was observed at 20 oC. Thereafter, seed germination reduced and the seed germination ranged from 25-78% for the temperature range of 25-35 oC. Seeds did not germinate at 40 oC (Figure 1A).

Effect of constant temperature (A) and alternating temperature (B) on germination of Ivy-leaved morning-glory seeds.
Maximum germination (94%) of Ivy-leaved morning-glory was observed at night/day temperature of 15/25 oC. However, there was no significant difference between 15/25 oC and 20/30 oC (Figure 1B). A substantial reduction in germination was observed at temperatures below 12.5/20 oC or above 30/42.5 oC. These results indicated that alternating temperatures could lead to a greater seed germination percentage than that at constant temperatures. These results were similar to those obtained by Nakamura and Hossain (2009Nakamura I, Hossain MA. Factors affecting the seed germination and seedling emergence of redflower ragleaf (Crassocephalum crepidioides). Weed Biol Manag. 2009;9(4):315-22.). They observed that germination of redflower ragleaf (Crassocephalum crepidioides) at alternating temperatures was higher than that at constant temperatures.
The results of experiment of Thullen and Keeley (1983Thullen RJ, Keeley PE. Germination, growth, and seed production of Ipomoea hederacea when planted at monthly intervals. Weed Sci. 1983;31(6):837-40. ) showed that ivy leaf morning-glory seed germinated the most at the night/day air temperature regime of 27/32 oC, but germination decreased as temperature increased up to 32/38 oC. No germination occurred at 10/15 oC. Oliveira and Norsworthy (2006Oliveira MJ, Norsworthy JK. Pitted morning-glory (Ipomoea lacunosa) germination and emergence as affected by environmental factors and seeding depth. Weed Sci. 2006;54(5):910-6.) reported that the optimum germination of Pitted morning-glory (Ipomoea lacunose) was between 20 and 25 oC. The highest germination of Tall morning-glory (Ipomoea purpurea) was observed at alternating temperatures of25/15 oC to 30/20 oC (Singh et al., 2012Singh M, Ramirez AHM, Sharma SD, Jhala AJ. Factors affecting the germination of tall morning-glory (Ipomoea purpurea). Weed Sci. 2012;60(1):64-8.). Chauhan and Abugho (2012Chauhan BS, Abugho SB. Threelobe morning-glory (Ipomoea triloba) germination and response to herbicides. Weed Sci. 2012;60(2):199-204.) stated that maximum germination of Three lobe morning-glory (Ipomoea triloba) was obtained in complete darkness at night/day temperatures of 15/25 oC and 25/35 oC, and the germination decreased to <10% and <20% at below 20/12.5 oC and above 35/25 oC, respectively.
Effect of high temperature pretreatment on germination
A three-parameter sigmoidal model described the effect of heating on seed germin ability. Germination was greater than 80% up to temperatures of 100 oC for 5 min, but the increasing temperature from 100 oC to 120 oC decreased severely germination of Ivy-leaved morning-glory, with no germination at >130 oC. On the basis of the fitted model, a temperature required for 50% reduction of maximum germination was estimated 119.63 oC (Figure 2). This suggests that high temperatures caused by fire can destroy part of the seeds. So, burning of crop residue after harvesting to prepare the land for planting the next crop in some areas could be effective to control this invasive weed by removing some seeds in the soil surface.

Vermeire and Rinella (2009Vermeire LT, Rinella MJ. Fire alters emergence of invasive plant species from soil surface-deposited seeds. Weed Sci. 2009;57(3):304-10.) in the study of fire effect on emergence of invasive species of leafy spurge (Euphorbia esula), Japanese brome (Bromus japonicas), Russian knapweed (Acroptilon repens), and spotted knapweed (Centaurea maculosa) observed that emergence was reduced from 79 to 88% in comparison to unburned treatment. Seed germination of Horse purslane (Trianthema portulacastrum) reduced up to 50% when exposed to temperature of 119 oC for 5 min (Lee et al., 2011Lee J, Chauhan BS, Johnson DE. Germination of fresh horse purslane (Trianthema portulacastrum) seeds in response to different environmental factors. Weed Sci. 2011;59(4):495-499. ).
Effect of osmotic stress on germination
Osmotic stress significantly affect seed germination of Ivy-leaved morning-glory, as provided a good fit by three-parameter sigmoidal model (Figure 3). Germination declined from 85%to 25% as osmotic potential dropped from 0 to -1.1 MPa and no germination was occurred at ≥-1.2 MPa. Germination decreased by 50% of the maximum at an osmotic potential of -1.03 MPa. The results show that this weed has tolerant to low water potential in the germination stage.

Three lobe morning-glory germination was reduced to 6% (from 90% in the non-treated control) at an osmotic potential of -0.6 MPa, and it was completely inhibited at -0.8 MPa (Chauhan and Abugho, 2012Chauhan BS, Abugho SB. Threelobe morning-glory (Ipomoea triloba) germination and response to herbicides. Weed Sci. 2012;60(2):199-204.).The germination of Tall morning-glory was very low (15%) at an osmotic stress of -0.3 and -0.4 MPa, and no germination was observed above that (Singh et al., 2012Singh M, Ramirez AHM, Sharma SD, Jhala AJ. Factors affecting the germination of tall morning-glory (Ipomoea purpurea). Weed Sci. 2012;60(1):64-8.). Germination of Pitted morning-glory (Oliveira and Norsworthy, 2006Oliveira MJ, Norsworthy JK. Pitted morning-glory (Ipomoea lacunosa) germination and emergence as affected by environmental factors and seeding depth. Weed Sci. 2006;54(5):910-6.) decreased with an increase in drought stress. Seed germination of Ipomoea asarifolia declined linearly with decreasing osmotic potential, and germination ceased at osmotic potential <-0.8 MPa (Dias-Filho, 1996Dias-Filho MB. Germination and emergence of Stachytarpheta cayennensis and Ipomoea asarifolia. Planta Daninha. 1996;14(2):118-26. ). This suggests that morning-glory species had varying sensitivity to osmotic stress. The results of our study indicated that Ivy-leaved morning-glory was more tolerant to water stress than other weed species of morning-glory. These results were similar to those reported by Crowley and Buchanan (1978Crowley RH, Buchanan GA. Competition of four morning-glory (Ipomoea spp.) species with cotton (Gossypium hirsutum). Weed Sci. 1978;26(5):484-8.).
Effect of salinity stress on germination
Osmotic potential caused by sodium chloride, ranging from 0 to -1 MPa, did not influence germination of the seeds of Ivy-leaved morning-glory. This species had excellent tolerance to salinity with 86 and 46% germination at 0 and -1.6 MPa, respectively. The osmotic potential due to NaCl required for 50% reduction of maximum germination (x50) was estimated -1.6 MPa (Figure 4), indicating that the inhibitory effect on seed germination was larger in PEG treatment than in NaCl treatment. Thus, this exotic species could pose an invasion threat to saline soil habitats.

Germination of Three lobe morning-glory occurred under a wide range of salt concentration, as seeds germinated 96-99% across a NaCl range of 0 to 250 mM (Chauhan and Abugho, 2012Chauhan BS, Abugho SB. Threelobe morning-glory (Ipomoea triloba) germination and response to herbicides. Weed Sci. 2012;60(2):199-204.).
Ninety percent of Tall morning-glory seeds germinated in distilled water, whereas germination dropped to less than 40% at a salt concentration of 50 mM, and complete inhibition of its germination occurred at 80 mM (Singh et al., 2012Singh M, Ramirez AHM, Sharma SD, Jhala AJ. Factors affecting the germination of tall morning-glory (Ipomoea purpurea). Weed Sci. 2012;60(1):64-8.).
Effect of pH on germination
Germination of Ivy-leaved morning-glory seeds was significantly influenced by pH (Figure 5). Maximum (89%) and minimum (9%) germination were observed at pH 5 and 8, respectively. There was no significant difference in germination between the pH levels of 4 to 7. However, treatments of pH above 7 significantly decreased seed germination compared to control in which germination was 85%. No germination was observed at pH>8. This suggests that acidic and neutral conditions were favorable for germination of Ivy-leaved morning-glory. Most Gorgan soils fall in the acidic-to-neutral range, although some soils have pH values greater than 7. Hence, soil pH factor may affect the distribution of this weed.

The highest Pitted morning-glory germination was observed at pH 3 with optimal germination in a range from 6 to 8 of buffered pH solutions (Oliveira and Norsworthy, 2006Oliveira MJ, Norsworthy JK. Pitted morning-glory (Ipomoea lacunosa) germination and emergence as affected by environmental factors and seeding depth. Weed Sci. 2006;54(5):910-6.). In contrast, maximum germination of Tall morning-glory (75%) was occurred at pH 6 with 31% germination at pH 5, and there was no germination at pH 4 (Singh et al., 2012Singh M, Ramirez AHM, Sharma SD, Jhala AJ. Factors affecting the germination of tall morning-glory (Ipomoea purpurea). Weed Sci. 2012;60(1):64-8.). The pH range for the germination of different morning-glory species in studies show that soil pH is a limiting factor for germination of these weed species.
Effect of burial depth on seedling emergence
Ivy-leaved morning-glory emergence as a function of a range of planting depths is presented in Figure 6. Emergence was at its maximum for seeds planted at depths of 1 to 10 cm, and dramatically decreased with increasing the depth of burial to 45% at 13 cm. No emergence observed at a planting depth of 15 cm. The burial depth which caused 50% inhibition of the maximum emergence was 12.93 cm (Figure 6).

Since less than 1% of the light can penetrate greater than 4 mm soil deep (Benvenuti et al., 2001Benvenuti S, Macchia M, Miele S. Quantitative analysis of emergence of seedlings from buried weed seeds with increasing soil depth. Weed Sci. 2001;49(4):528-35. ), the results explained that Ivy-leaved morning-glory seeds are not a light requirement for emergence. Hence, this species is able to emerge even after the closure of canopy of crop.
Investigation of unemerged seeds at the end of the experiment (14 d) showed that they had decayed (fatal germination). It seems that seeds buried in depths of ≥13 cm germinated, but the seedling died before reaching the soil surface. Considering the fatal germination at these depths, burial of Ivy-leaved morning-glory seeds to a 15 cm, or possibly greater, depth with tillage operations may be an appropriate method of reducing Ivy-leaved morning-glory seed banks through fatal germination.
Effect of flooding on seedling emergence
Flooding had a suppressive effect on the emergence of Ivy-leaved morning-glory. Only 9% of seeds produced seedlings when flooding duration was for three days, whilst under normal (no flooding) conditions, emergence was 96%. Seeds of Ivy-leaved morning-glory buried in flooded soil for over three days failed to emerge (Figure 7). These findings indicated that Ivy-leaved morning-glory was sensitive to flooding. The results of the present study agree with similar studies showing that emergence of other Morning-glory species [such as Palm leaf morning-glory (Ipomoea wrightii), Pitted morning-glory and Tall morning-glory] were also inhibited at flooding conditions (Gealy, 1998Gealy D. Differential response of palm leaf morning-glory (Ipomoea wrightii) and pitted morning-glory (Ipomoea lacunosa) to flooding. Weed Sci. 1998;46(2):217-47. ; Singh et al., 2012Singh M, Ramirez AHM, Sharma SD, Jhala AJ. Factors affecting the germination of tall morning-glory (Ipomoea purpurea). Weed Sci. 2012;60(1):64-8.). It suggests that this species is not capable to be established in flooded or low drained soils. According to the impact of flooding on the emergence of this plant, flooding can be a suitable method for the management of this weed.

In conclusion, the results of our study indicated that environmental conditions had substantial effects on seed germination and emergence of Ivy-leaved morning-glory. This species is a tropical plant that its germination occurred from temperatures of 15 oC to 35 oC. However, seeds germinated greater at alternating temperatures. This weed had excellent tolerance to salinity and drought stresses. Therefore, these two environmental factors are not limiting factors of this species spread in dry farmlands of semi-arid regions in Iran. Seed germination was affected by pH. The results suggest that Ivy-leaved morning-glory tends to germinate better in acidic soils compared with neutral-to-alkaline soil environments. All seeds buried at 1-10 cm depths were able to emerge, and no germinable seeds were found at depths greater than 15 cm. So, burying seeds deep enough (>15 cm) can be beneficial to prevent morning-glory emergence. However, subsequent tillage operations must be at less depths to avoid transferring seeds near to the soil surface. Seedling emergence was inhibited by flooding >3 d. Thus, this technique could be exploited in the management of this weed. High temperatures also decreased seed germination of this species; therefore, residue burning can be effective in reducing the population of the seed bank.
REFERENCES
- Ali HH, Tanveer A, Nadeem MA, Asghar HN, Javid MM. Germination ecology of Rhynchosia capitata: an emerging summer weed in Asia. Planta Daninha. 2013;31(2):249-57.
- Altop EK, Mennan H, Philippo CJ, Zandstra BH. Effect of the burial depth and environmental factors on the seasonal germination of bearded sprangletop (Leptochloa fusca [L.] Kunth ssp. fascicularis [Lam.] N. Snow). Weed Biol Manag. 2015;14(4):147-58.
- Anonymous. Ipomoea hederacea (Convolvulaceae): the global compendium of weeds [acesso em: 10 mar 2018]. Disponível em: Disponível em: http://www.hear.org/gcw/html/autogend/species/10267.HTM
» http://www.hear.org/gcw/html/autogend/species/10267.HTM - Asgarpour R, Ghorbani R, Khajeh-hosseini M, Mohamadvand E. Germination of spotted spurge (Chamaesyce maculata) seeds in response to different environmental factors. Weed Sci. 2015;63(2):502-10.
- Baskin CC, Milberg P, Andersson L, Baskin JM. Germination ecology of seeds of the annual weeds Capsella bursa-pastoris and Descurainia sophia originating from high northern latitudes. Weed Res. 2004;44(2):60-8.
- Benvenuti S, Macchia M, Miele S. Quantitative analysis of emergence of seedlings from buried weed seeds with increasing soil depth. Weed Sci. 2001;49(4):528-35.
- Chauhan BS, Gill G, Preston C. African mustard (Brassica tournefortii) germination in southern Australia. Weed Sci. 2006a;54(5):891-7.
- Chauhan BS, Gill G, Preston C. Influence of environmental factors on seed germination and seedling emergence of Oriental mustard (Sisymbrium orientale). Weed Sci. 2006b;4(6):1025-31.
- Chauhan BS, Abugho SB. Threelobe morning-glory (Ipomoea triloba) germination and response to herbicides. Weed Sci. 2012;60(2):199-204.
- Chauhan BS, Johnson DE. Germination ecology of southern crabgrass (Digitaria ciliaris) and India crabgrass (Digitaria longiflora): two important weeds of rice in tropics. Weed Sci. 2008;56(5):722-8.
- Crowley RH, Buchanan GA. Competition of four morning-glory (Ipomoea spp.) species with cotton (Gossypium hirsutum). Weed Sci. 1978;26(5):484-8.
- Culpepper AS, Gimenez AE, York AC, Batts RB, Wilcut JW. Morning-glory (Ipomoea spp.) and large crabgrass (Digitaria sanguinalis) control with glyphosate and 2,4-DB mixtures in glyphosate-resistant soybean (Glycine max). Weed Technol. 2001;15(1):56-61.
- Dias-Filho MB. Germination and emergence of Stachytarpheta cayennensis and Ipomoea asarifolia Planta Daninha. 1996;14(2):118-26.
- Eslami SV, Afghani F, Mahmoodi S. Effects of some environmental factors on germination and longevity of downy brome (Bromus tectorum) seeds. Iran J Weed Sci. 2010;4(2):47-57.
- Ghaderi-Far F, Gherekhloo J, Alimagham SM. Influence of environmental factors on seed germination and seedling emergence of yellow sweet clover (Melilotus officinalis). Planta Daninha. 2010;28(3):436-69.
- Gealy D. Differential response of palm leaf morning-glory (Ipomoea wrightii) and pitted morning-glory (Ipomoea lacunosa) to flooding. Weed Sci. 1998;46(2):217-47.
- Koger CH, Reddy KN, Ppston DH. Factors affecting seed germination, seedling emergence, and survival of texas weed (Caperonia palustris). Weed Sci. 2004;52(6):989-95.
- Lee J, Chauhan BS, Johnson DE. Germination of fresh horse purslane (Trianthema portulacastrum) seeds in response to different environmental factors. Weed Sci. 2011;59(4):495-499.
- Maraghni M, Gorai M, Naffati M. Seed germination at different temperatures and water stress levels, and seedling emergence from different depths of Ziziphus lotus S Afr J Bot. 2010;76(3):453-9.
- Mennan H, Ngouajio M. Seasonal cycles in germination and seedling emergence of summer and winter population of catch weed bedstraw and wild mustard. Weed Sci. 2006;54(1):114-20.
- Nakamura I, Hossain MA. Factors affecting the seed germination and seedling emergence of redflower ragleaf (Crassocephalum crepidioides). Weed Biol Manag. 2009;9(4):315-22.
- Oliveira MJ, Norsworthy JK. Pitted morning-glory (Ipomoea lacunosa) germination and emergence as affected by environmental factors and seeding depth. Weed Sci. 2006;54(5):910-6.
- Pahlevani AH, Sajedi S. Alerting occurrence of several noxious weeds and invasive plants in arable lands in Iran. Rostaniha. 2011;12 (2):129-34.
- Peters NCB, Atkins HA, Brain P. Evidence of differences in seed dormancy among populations of Bromus sterilis Weed Res. 2000;40(5):457-78.
- Sadeghloo A, Asghari J, Ghaderi-Far F. Seed germination and seedling emergence of velvetleaf (Abutilon theophrasti) and barnyardgrass (Echinochloa crus-galli). Planta daninha. 2013;31(2):259-66.
- Singh BP, Singh S. Chemical investigation of seed of Ipomoea hederacea and its biological activity. J Chem Pharm Res. 2012;4(2):1441-8.
- Singh M, Ramirez AHM, Sharma SD, Jhala AJ. Factors affecting the germination of tall morning-glory (Ipomoea purpurea). Weed Sci. 2012;60(1):64-8.
- Susko DJ, Mueller JP, Spears JF. Influence of environmental factors on germination and emergence of Pueraria lobata Weed Sci. 1999;47(5):585-8.
- Tanveer A, Sibtain M, Javaid MM, Ali HH. Germination ecology of wild onion: a rainfed crop weed. Planta Daninha. 2014;32(1):69-80.
- Tenaglia D. Photographs and descriptions of the flowering and non-flowering plants of Missouri [acesso em: 10 nov 2017]. Disponível em: Disponível em: http://www.missouriplants.com/
» http://www.missouriplants.com/ - Thullen RJ, Keeley PE. Germination, growth, and seed production of Ipomoea hederacea when planted at monthly intervals. Weed Sci. 1983;31(6):837-40.
- Vermeire LT, Rinella MJ. Fire alters emergence of invasive plant species from soil surface-deposited seeds. Weed Sci. 2009;57(3):304-10.
- Ul-Haq MZ, De-Feo V, Riaz M. Ipomea hederaceaJacq: a medicinal herb with promising health benefits. Molecules. 2012;17(11):13132-45.
Publication Dates
-
Publication in this collection
17 Apr 2020 -
Date of issue
2020
History
-
Received
24 Apr 2018 -
Accepted
01 Nov 2018

 Vertical bars represent standard error of the means.
Vertical bars represent standard error of the means.
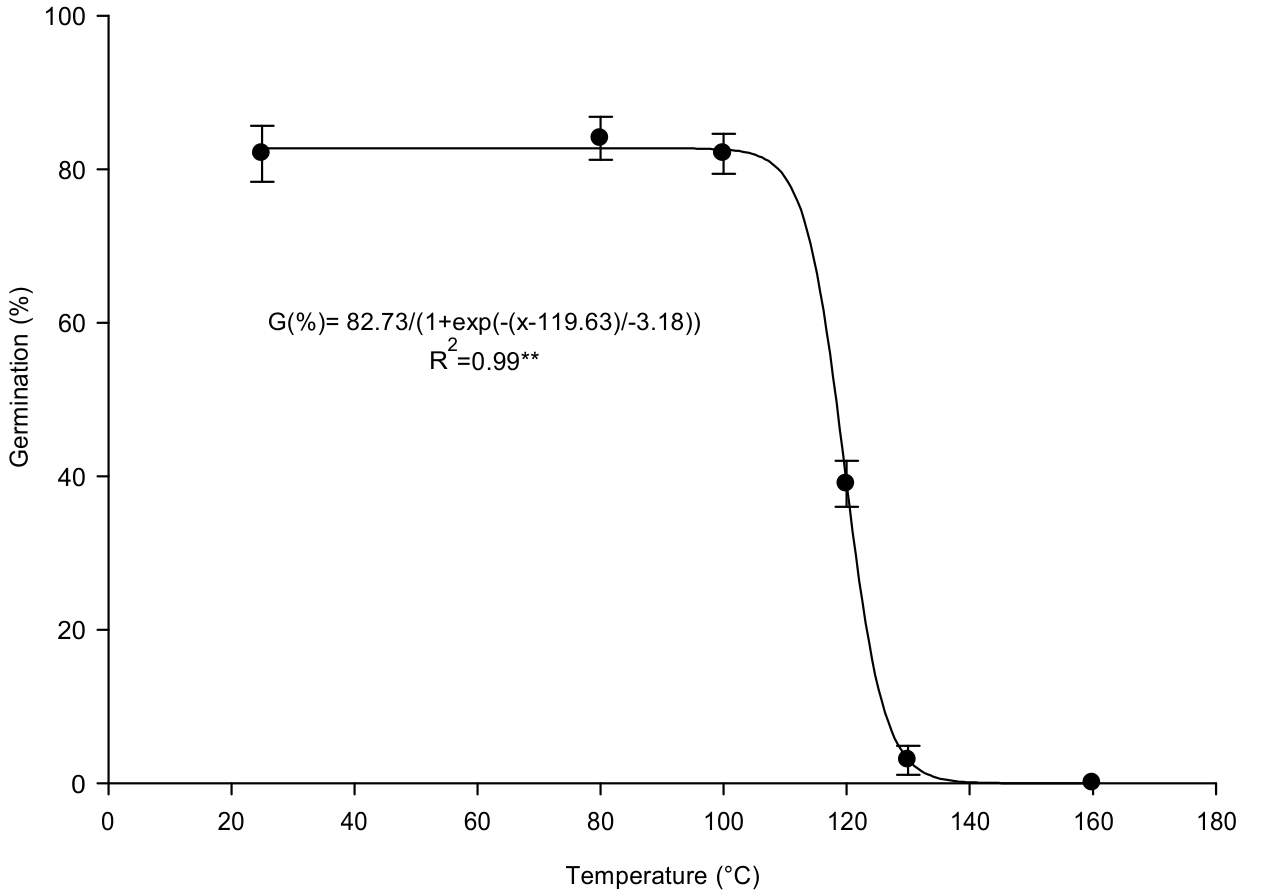 Vertical bars represent standard error of mean.
Vertical bars represent standard error of mean.
 Vertical bars represent standard error of mean.
Vertical bars represent standard error of mean.
 Vertical bars represent standard error of mean.
Vertical bars represent standard error of mean.
 Vertical bars represent standard error of mean.
Vertical bars represent standard error of mean.
 Vertical bars represent standard error of mean.
Vertical bars represent standard error of mean.
 Vertical bars represent standard error of mean.
Vertical bars represent standard error of mean.