Resumos
Estudos de biologia floral e dos animais visitantes, em anos diferentes, foram feitos em duas populações (Brotas e Campinas) de jurubeba, um arbusto invasor neotropical. As flores, do tipo aberto, têm odor muito suave. Pétalas de côr violeta-pálido, contrastando com as anteras amarelas, formam um conjunto visualmente atrativo. O pólen é a única recompensa para os visitantes. A espécie é alógama. Uma média de 19% das flores em ambas as populações apresentaram estilete curto e somente flores de estilete longo formaram frutos, indicando a existência de andromonoicia funcional. As taxas de frutificação em condições naturais diferiram entre Brotas (43%) e Campinas (17%). A frutificação por polinização manual foi de 46% em Brotas e nula em Campinas, provavelmente devido a um longo período de seca. A regularidade da microsporogênese e a alta taxa de viabilidade dos grãos de pólen mostraram a normalidade do processo de reprodução sexuada. Anteras poricidas e outras características florais de S. paniculatum estão associadas à síndrome da polinização vibrátil, que requer abelhas especializadas para a retirada de pólen. Os principais polinizadores foram Oxaea flavescens, Bombus morio, e Xylocopa frontalis na população de Campinas e Augochloropsis sp 1 e Augochloropsis sp 2 na de Brotas.
Studies on floral biology and visitors were done over different years in two populations (Brotas and Campinas) of "jurubeba," a common neotropical invader shrub. The flowers are dish-shaped and have a mellow odour. Pale-violet petals contrasting with yellow anthers make a visually attractive set. Pollen is the only reward for visitors. The species is alogamous. On average, 19% of the flowers in both populations had short styles; only long-styled flowers generated fruits, thus presenting a functional andromonoecism. Fruiting ratios in natural conditions differed between Brotas (43%) and Campinas (17%). The fruiting ratio due to manual polinization was 46% in Brotas and nihil in Campinas, this probably being due to a long dry period. Both microsporogenesis regularity and pollen high viability indicate that the process of sexual reproduction is normal. Poricide anthers and other flower features of S. paniculatum are associated with the buzz pollination syndrome, which requires bees with a specific behavior for pollen removal. The main pollinators are Oxaea flavescens, Bombus morio, and Xilocopa frontalis in the Campinas population, and Augochloropsis sp 1 e Augochloropsis sp 2 in the Brotas population.
Solanum; andromonoecism; buzz pollination
Biologia floral e reprodução de Solanum paniculatum L. (Solanaceae) no estado de São Paulo, Brasil
ELIANA REGINA FORNI-MARTINS1 1 . Departamento de Botânica, Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6109, 13083-970 Campinas, SP, Brasil. , MARCIA CRISTINA MENDES MARQUES2 1 . Departamento de Botânica, Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6109, 13083-970 Campinas, SP, Brasil. e MARISTERRA R. LEMES3 1 . Departamento de Botânica, Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6109, 13083-970 Campinas, SP, Brasil.
(recebido em 24/04/97; aceito em 03/02/98)
ABSTRACT - (Floral biology and reproduction of Solanum paniculatum L. (Solanaceae) in the state of São Paulo, Brazil). Studies on floral biology and visitors were done over different years in two populations (Brotas and Campinas) of "jurubeba," a common neotropical invader shrub. The flowers are dish-shaped and have a mellow odour. Pale-violet petals contrasting with yellow anthers make a visually attractive set. Pollen is the only reward for visitors. The species is alogamous. On average, 19% of the flowers in both populations had short styles; only long-styled flowers generated fruits, thus presenting a functional andromonoecism. Fruiting ratios in natural conditions differed between Brotas (43%) and Campinas (17%). The fruiting ratio due to manual polinization was 46% in Brotas and nihil in Campinas, this probably being due to a long dry period. Both microsporogenesis regularity and pollen high viability indicate that the process of sexual reproduction is normal. Poricide anthers and other flower features of S. paniculatum are associated with the buzz pollination syndrome, which requires bees with a specific behavior for pollen removal. The main pollinators are Oxaea flavescens, Bombus morio, and Xilocopa frontalis in the Campinas population, and Augochloropsis sp 1 e Augochloropsis sp 2 in the Brotas population.
RESUMO - (Biologia floral e reprodução de Solanum paniculatum L. (Solanaceae) no estado de São Paulo, Brasil). Estudos de biologia floral e dos animais visitantes, em anos diferentes, foram feitos em duas populações (Brotas e Campinas) de jurubeba, um arbusto invasor neotropical. As flores, do tipo aberto, têm odor muito suave. Pétalas de côr violeta-pálido, contrastando com as anteras amarelas, formam um conjunto visualmente atrativo. O pólen é a única recompensa para os visitantes. A espécie é alógama. Uma média de 19% das flores em ambas as populações apresentaram estilete curto e somente flores de estilete longo formaram frutos, indicando a existência de andromonoicia funcional. As taxas de frutificação em condições naturais diferiram entre Brotas (43%) e Campinas (17%). A frutificação por polinização manual foi de 46% em Brotas e nula em Campinas, provavelmente devido a um longo período de seca. A regularidade da microsporogênese e a alta taxa de viabilidade dos grãos de pólen mostraram a normalidade do processo de reprodução sexuada. Anteras poricidas e outras características florais de S. paniculatum estão associadas à síndrome da polinização vibrátil, que requer abelhas especializadas para a retirada de pólen. Os principais polinizadores foram Oxaea flavescens, Bombus morio, e Xylocopa frontalis na população de Campinas e Augochloropsis sp 1 e Augochloropsis sp 2 na de Brotas.
Key words - Solanum, andromonoecism, buzz pollination
Introdução
O gênero Solanum possui mais de 1700 espécies (Willis 1973), sendo bem representado na América tropical, incluindo o Brasil (Sendtner 1846). Na biologia floral e reprodutiva de espécies do gênero são caracteres comuns a polinização por abelhas, a deiscência poricida das anteras e a predominância de alogamia (Buchmann et al. 1977, Symon 1979). A retirada de pólen de anteras poricidas requer comportamento especial das abelhas, sendo a polinização destas flores denominada de polinização vibrátil (Buchmann et al. 1977). Embora muitas espécies de Solanum tenham flores monóclinas típicas, há considerável variação de formas sexuais, incluindo andromonoicismo, androdioicismo e dioicismo (Anderson 1979, Coleman & Coleman 1982, Oliveira Filho & Oliveira 1988, Anderson & Symon 1989).
Solanum paniculatum L., vulgarmente conhecida como jurubeba, ocorre em toda a América tropical (Leitão-Filho et al. 1975). A ela são atribuídas propriedades medicinais, sendo popularmente utilizada no tratamento da icterícia, da hepatite crônica e de febres intermitentes (Pio Corrêa 1969), além de usos culinários (Zurlo & Brandão 1990). É também considerada uma planta invasora, que ocupa os mais variados tipos de solo (Leitão-Filho et al. 1975). Grant (1975) atribuiu às plantas invasoras algumas características típicas, como a capacidade de colonização rápida de ambientes abertos (inclusive por ação antrópica) e a reprodução predominantemente autogâmica, o que lhes confere alta uniformidade genética em nível populacional, não descartando, entretanto, surtos esporádicos de recombinação.
O presente trabalho, desenvolvido em duas populações de S. paniculatum, apresenta e discute aspectos relacionados à sua biologia floral, aos visitantes florais e ao sistema de reprodução.
Material e métodos
As observações sobre S. paniculatum foram feitas em duas populações, ambas no estado de São Paulo: uma no Centro Experimental do Instituto Agronômico de Campinas (22o 54'S e 47o 05W, 674 m) - população I - e outra nas proximidades do Viveiro Municipal de Brotas (22o 08'S e 48o 50'W, 470 m) - população II - nos períodos de setembro a novembro de 1985 e outubro a novembro de 1991, respectivamente. Ambas as populações encontravam-se em ambientes alterados. A população I compreendia cerca de 250 indivíduos, divididos em dois grupos distantes 80 m entre si e circundados por campos de cultura agrícola. A população II era composta por aproximadamente 25 indivíduos, próximos a uma floresta ripícola. Foram registradas informações sobre o hábito das plantas, a morfologia das flores, o desenvolvimento dos frutos, o sistema reprodutivo e o comportamento dos visitantes florais.
A cor das flores foi determinada através da comparação com um catálogo de cores (Kornerup & Wanscher 1963). Para a determinação dos pigmentos florais, as flores foram colocadas em contato com vapores de amoníaco (Hess 1983). As regiões de absorção de ultravioleta foram detectadas através do teste com FeCl3 diluído em éter sulfúrico 1% (Vogel 1950). A ocorrência de glândulas de odor foi determinada por meio do uso de vermelho-neutro diluído em água (pH 7) na proporção 1 g : 1000 ml (Vogel 1962).
O número cromossômico e a regularidade do processo meiótico (microsporogênese) foram determinados por meio da fixação de botões florais em Carnoy (etanol : ácido acético na proporção 3:1), sendo as anteras esmagadas em carmin acético (Medina & Conagin 1964). Para a análise da viabilidade de pólen, cerca de 1000 grãos de pólen, provenientes de flores com estilete longo e curto, de cada população, foram corados com carmin acético (Medina & Conagin 1964). As observações da microsporogênese e da viabilidade de pólen foram feitas em microscópio óptico. A receptividade dos estigmas florais foi verificada com a aplicação de água oxigenada 10 vol. (Zeisler 1938).
Para a determinação do sistema de reprodução, botões florais em pré-antese foram envolvidos com sacos de papel, fazendo-se a emasculação nos testes para autopolinização e polinização cruzada. Posteriormente, foram feitas polinizações manuais com pólen da mesma flor ou do mesmo indivíduo (autopolinização) e com pólen de indivíduos diferentes (polinização cruzada), distanciados pelo menos três metros entre si. Foi testada a fertilidade de flores com estilete longo e curto, que foram utilizadas ora como doadoras ora como receptoras de pólen. As flores polinizadas manualmente foram novamente encobertas com sacos de papel, para impedir contatos posteriores com visitantes. Na população II também foi testada a ocorrência de polinização espontânea, sendo as flores envolvidas em sacos de papel e não polinizadas manualmente. A ocorrência de apomixia foi verificada em botões emasculados e ensacados durante quatro dias. Algumas flores foram marcadas e mantidas intactas e desprotegidas para observar o desenvolvimento de frutos em condições naturais.
Os estudos sobre o sistema reprodutivo foram complementados com observações do crescimento do tubo polínico nos estiletes nas flores da população II. Pistilos dos dois tipos florais foram acondicionados em placas de ágar, onde foram feitas polinizações controladas, com pólen proveniente de flores de estilete curto ou longo, da mesma planta ou de plantas diferentes. Foram utilizados cinco pistilos para cada tipo de cruzamento. Os pistilos foram fixados em FAA70 após 24 e/ou 48 horas da realização das polinizações. Posteriormente, foram analisados sob microscopia de fluorescência, segundo a técnica de Martin (1959).
O comportamento dos visitantes florais foi observado visualmente, em diferentes horários (das 6:30 às 17:00 h), sendo complementado com a análise de fotografias. Espécimes de visitantes foram coletados para posterior identificação e para obtenção de medidas do comprimento médio do corpo de cada espécie.
As exsicatas do material botânico foram depositadas no Herbário do Departamento de Botânica da Universidade Estadual de Campinas (UEC 17.935, população I e UEC 26.876, população II).
Resultados
Características das plantas - As plantas de Solanum paniculatum são perenes, arbustivas, atingindo até 2 m de altura, podendo ser classificadas como nanofanerófitas no sistema de formas de vida de Raunkiaer (Mueller-Dombois & Ellenberg 1974). Observou-se a rebrota após a poda (por roçada) do ramo principal quase no nível do solo. Possuem inflorescências em forma de panículas terminais, nas quais as flores atingem a maturidade da base para o ápice. Observou-se floração durante o período de setembro a novembro, mas registros de herbário mostram que o período de floração no estado de São Paulo pode ser maior, indo até fevereiro.
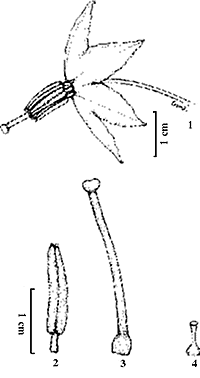
Características das flores - As flores possuem cinco pétalas unidas, de cor violeta-pálido (17 A 3), de acordo com Kornerup & Wanscher (1963). A corola é pigmentada por antocianina. As flores apresentam cinco anteras amarelas, poricidas, vistosas e tubulosas, que se dispõem de tal modo a formar um cone ao redor do estigma (figuras 1 e 2); o estigma é capitado e o estilete pode ser longo ou curto (figuras 3 e 4), sendo verde em ambos os casos. O ovário das flores com estilete curto é menor que o das flores com estilete longo (figuras 3 e 4), porém ambos são multiovulados.
Figuras 1-4. Morfologia floral de Solanum paniculatum. 1. Flor monóclina, com estilete longo; 2. Estame; 3. Gineceu de uma flor com estilete longo; 4. Gineceu de flor com estilete curto.
A antese ocorre em torno das 05:00 h e, após aproximadamente uma hora, os poros das anteras se abrem e expõem o pólen pulverulento. As flores permanecem abertas por um a dois dias, período em que os estigmas ainda se apresentam receptivos.
Detectou-se que as pétalas, os estames (exceto conectivos) e o estigma absorvem luz ultravioleta. As flores possuem odor adocicado muito suave. Glândulas ativas, provavelmente de odor (osmóforos), foram encontradas nas pétalas (exceto nas nervuras), estigmas, ápices das anteras, cálice e pedúnculo das flores.
Há duas formas florais: uma com estilete longo (L), com comprimento maior que o das anteras e estigma exposto; e outra com estilete curto (C), localizado abaixo do nível das anteras, sem exposição do estigma. Apesar da diferença no comprimento do estilete, as anteras e corolas dos dois tipos florais apresentaram dimensões semelhantes nas duas populações (tabela 1). Não há um padrão definido quanto à posição ou à proporção de flores de estilete longo ou curto em uma inflorescência ou em um indivíduo. Em média, 19% das flores amostradas apresentaram estilete curto.
Tabela 1. Valores médios (mm) do diâmetro da corola, do comprimento do cone das anteras e do comprimento do estilete em flores de Solanum paniculatum das populações I (Campinas) e II (Brotas). C-flor de estilete curto, L-flor de estilete longo (± desvio padrão).
Tanto flores com estilete curto como as de estilete longo apresentaram grãos de pólen viáveis (acima de 87%), não havendo grandes diferenças de viabilidade entre as duas populações. Encontrou-se regularidade no processo meiótico. O número cromossômico (n) foi igual a 12.
Sistema reprodutivo - Flores de estilete curto nunca produziram frutos, mesmo em condições naturais. A formação de frutos ocorreu somente em flores com estilete longo em duas condições: 1) em condições naturais, em ambas as populações, e 2) em flores de estilete longo após polinização cruzada com pólen proveniente de flores L ou C, apenas na população II. Não houve produção de frutos após autopolinização ou emasculação (tabela 2).
Tabela 2. Testes sobre o sistema reprodutivo de Solanum paniculatum, nas populações I (Campinas) e II (Brotas). C - flores de estilete curto, L - flores de estilete longo
A análise de pistilos, previamente polinizados e fixados, indicou: 1) nas autopolinizações, após 24 horas, já se podia observar grãos de pólen germinados e tubos polínicos em início de desenvolvimento; nas 24 horas seguintes, os tubos polínicos haviam crescido muito pouco, limitando-se à região do estilete próxima ao estigma; 2) nas polinizações cruzadas, após 24 horas, os tubos polínicos alcançaram o ovário e penetraram nos óvulos das flores com estilete longo; nas flores com estilete curto, os tubos polínicos cresceram apenas até o início do estilete.
Visitantes - Foram observadas oito espécies de abelhas visitando flores de S. paniculatum na população I e cinco espécies na população II (tabela 3). Em ambas as populações, as visitas das abelhas iniciavam-se a partir das 8:00 h e estendiam-se até, no máximo, às 16:00 h. O período de maior atividade de visitantes era entre 10:00 e 14:00 h. Centris sp 2 (população I) restringia suas visitas ao período entre 10:00 e 12:00 h.
Tabela 3. Abelhas visitantes de Solanum paniculatum nas populações I (Campinas) e II (Brotas) e seus respectivos comprimentos do corpo (mm) e freqüência de visitas: (+++) muito freqüente, (++) freqüente, (+) pouco freqüente.
Na população I as abelhas mais comumente observadas foram Oxaea flavescens (Oxaeidae), Bombus morio (Apidae), Xylocopa frontalis (Anthophoridae) e Exomalopsis sp. (Anthophoridae), nesta ordem.
Oxaea flavescens aproximava-se da flor frontalmente e pousava diretamente sobre o cone das anteras, no qual se agarrava com as patas. Em seguida, recurvava o abdome sobre a flor, contatando o estigma. Neste momento a abelha realizava movimento vibratório, liberando o pólen, que era depositado nas partes ventrais do tórax e do abdome. Habitualmente, um grande número de flores do mesmo indivíduo ou de indivíduos diferentes de S. paniculatum era visitado seqüencialmente por esta abelha. Entre uma visita e outra, a abelha apresentou o comportamento de limpeza e transferência de pólen.
Bombus morio e Xylocopa frontalis apresentaram comportamento de aproximação às flores, de pouso, de contacto com o estigma e de movimento vibratório semelhante ao descrito para Oxaea flavescens. Na maioria das flores, B. morio pousava, efetuava movimento vibratório, segurava-se na corola ou nas anteras e realizava a limpeza do abdome, transferindo o pólen para as corbículas. Em outras flores, a abelha pousava e efetuava apenas a vibração ou apenas o comportamento de limpeza, voando em seguida para outra flor.
Exomalopsis sp. pousava diretamente sobre as anteras, caminhava em direção aos poros das anteras e efetuava movimento vibratório. Após este procedimento da abelha, era possível verificar pólen caído sobre as pétalas.
Bombus atractus, Centris sp 1 e Centris sp 2 recolhiam pólen de S. paniculatum também por movimentos vibratórios, como descrito em Oxaea flavescens, porém a freqüência das visitas era menor do que a observada para as demais abelhas. Foi observada uma espécie de Colletidae deslocando-se e coletando pólen caído sobre as anteras.
Na população II, as abelhas mais freqüentes foram, em seqüência, Augochloropsis sp 1, Augochloropsis sp 2 (Halictidae) e Megamalopsis sp. (Anthophoridae). As duas espécies de Augochloropsis apresentaram comportamento de visita muito semelhante entre si. Ambas aproximavam-se da flor e dirigiam-se ao cone das anteras. Com as patas anteriores e, ocasionalmente, com as medianas, agarravam-se a uma antera (algumas vezes, duas), recurvavam o corpo sobre o poro e executavam o movimento vibratório, que podia se repetir por duas ou três vezes. Em seguida, deslocavam-se em direção a outra antera, onde repetiam o movimento. Durante o deslocamento, a abelha contatava várias vezes o estigma com o abdome. A coleta de pólen era realizada nas cinco anteras, mas, se as mesmas já estivessem vazias, as abelhas retiravam-se logo após os primeiros movimentos vibratórios. A limpeza do abdome era feita freqüentemente, após a coleta em uma ou duas anteras.
Megamalopsis sp. pousava na flor na região do cone das anteras ao qual se alinhava perpendicularmente, de modo que seu abdome contatava os poros. O movimento vibratório era breve e uma grande quantidade de pólen aderia ao corpo da abelha.
O comportamento de visita de Bombus morio às plantas de S. paniculatum na população II foi semelhante ao descrito para a população I, no entanto a freqüência de visitas foi bem menor (duas visitas). Apenas um indivíduo de Exomalopsis exomalopsis foi observado e, ao pousar na flor, caminhou sobre a corola, provavelmente à procura de pólen.
Além das abelhas, foram observados outros insetos visitando flores de S. paniculatum. Na população I, foi observada a presença de uma espécie de Meloidae (Coleoptera) que danificava corola, estames e estilete. Na população II vários indivíduos de Brachygastra lecheguana (Vespidae, Hymenoptera) foram freqüentemente observados alimentando-se de partes de botões florais, ocasionando danos. Também se encontraram besouros (Chrisomelidae, Coleoptera) caminhando sobre a flor de S. paniculatum.
Discussão
As flores de Solanum paniculatum podem ser classificadas como do tipo aberto (Faegri & Van der Pijl 1971), sendo actinomorfas, não tubulosas e tendo órgãos sexuais agrupados no centro. O pólen é a única recompensa aos visitantes florais. O androceu tem características de flores-pólen oligândricas (Vogel 1978): poucas anteras, grandes, vistosas, com deiscência poricida e produzindo pólen pulverulento em grande quantidade, mas liberado em pequenas porções por movimentos vibratórios dos visitantes.
Além destas características, outras, como o período de floração relativamente longo, a antese ao amanhecer, a duração de cada flor (dois dias), a produção diária de relativamente poucas flores, a presença de estigma receptivo e pólen disponível nas primeiras horas do dia, a cor da corola e das anteras poricidas e os grãos de pólen pequenos e esbranquiçados, indicam que S. paniculatum é uma típica planta com síndrome de polinização vibrátil (Buchmann 1983). O diâmetro reduzido dos grãos de pólen dos dois tipos florais (tabela 2) está dentro da variação de diâmetro apresentada para flores com esta síndrome de polinização (Buchmann 1983).
A absorção de raios ultravioleta em toda a corola e nas anteras, bem como a pigmentação da corola (antocianina) de S. paniculatum parecem estar associadas ao padrão de reconhecimento das flores por parte das abelhas (Vogel 1983). Entretanto, Coleman & Coleman (1982) mostraram que a retirada das corolas de S. palinacanthum causou pequena diminuição na freqüência de visitas, ao contrário do que ocorreu com a retirada das anteras, indicando que o padrão de coloração das anteras tem importante efeito atrativo para as abelhas.
Aromas florais fortes e adocicados produzidos por substâncias voláteis estão usualmente presentes em flores com síndrome de polinização vibrátil, como Amourexia, Cassia e Solanum, mas também podem estar ausentes (Buchmann 1983). É possível que os osmóforos nas flores (corola, ápices de anteras, cálice e pedúnculo floral) emitam odores suaves. Em S. palinacanthum, as anteras e, presumivelmente, o pólen exalam odores, que são tidos como atrativos florais (Coleman & Coleman 1982).
O gênero Solanum exibe várias formas florais, havendo espécies monóicas com flores monóclinas e outras com variações referentes à andromonoicia, androdioicia e até dioicia. Nos indivíduos andromonóicos, androdióicos ou dióicos, ocorrem muitas vezes flores morfologicamente semelhantes, porém funcionalmente distintas (Anderson 1979, Coleman & Coleman 1982, Oliveira-Filho & Oliveira 1988). Na seção Torva, à qual S. paniculatum pertence, já foram descritas espécies andromonóicas e monóicas (Symon 1979).
Os testes sobre o sistema reprodutivo permitem concluir que S. paniculatum é uma espécie andromonóica, em que as flores L têm androceu e gineceu funcionais e as flores C são funcionalmente masculinas, apesar de apresentarem óvulos (tabela 2). Apesar da amostragem ser pequena em alguns testes e do insucesso em alguns deles, especialmente na população I, os resultados foram confirmados pela análise de pistilos, pois os tubos polínicos alcançaram os óvulos apenas em flores de estilete longo. Oliveira-Filho & Oliveira (1988) estudaram a espécie S. lycocarpum e também relataram a ocorrência de óvulos em flores funcionalmente masculinas. Diferentemente do observado em S. paniculatum, Oliveira-Filho & Oliveira (1988) relataram a penetração de tubos polínicos nos óvulos de flores masculinas de S. lycocarpum, porém estas eram sempre abortadas. Já em S. palynacanthum, que também é andromonóica (Coleman & Coleman 1982), os grãos de pólen nem sequer germinaram em estigmas de flores funcionalmente masculinas. Os resultados aqui obtidos para S. paniculatum confirmam a derivação da andromonoicia a partir do hermafroditismo no gênero Solanum (Oliveira-Filho & Oliveira 1988), em um processo em que não houve supressão de alguns caracteres.
Solanum paniculatum é uma espécie alógama sem apomixia (tabela 2). No gênero Solanum há espécies autógamas e alógamas (Gardé 1959, Grun 1961, Whalen & Anderson 1981). As espécies alógamas possuem sistema de incompatibilidade gametofítico (Pandey 1960, Heslop-Harrison 1975). A auto-incompatibilidade é a condição ancestral em Solanum, porém existem alguns casos no gênero em que é secundária (Whalen & Anderson 1981). Em Solanum, a auto-incompatibilidade parece estar restrita ao subgênero Potatoe e a algumas espécies do subgênero Leptostemonum, ao qual S. paniculatum pertence. Até o momento, na Seção Torva, a incompatibilidade genética é citada apenas para S. paniculatum, enquanto que S. hispidum (Baksh & Iqbal 1978) e S. torvum (Hossain 1973) são autocompatíveis. Anderson & Symon (1989) sugeriram que a andromonoicia reduz a autogamia em várias espécies de Solanum da Austrália.
A alogamia indica alta variabilidade genética em S. paniculatum. A autogamia, comum em plantas invasoras herbáceas, confere pequena variabilidade genética em nível populacional e capacidade de formar uma população rapidamente e com adaptação imediata às condições do ambiente (Grant 1975). Entretanto, alguns grupos de plantas invasoras perenes, do tipo hemicriptófitas (brotam do sistema subterrâneo no nível do solo) e caméfitas (brotam de uma base caulinar aérea geralmente abaixo de 50 cm), são, em geral, alógamas e constituídas de populações relativamente variáveis geneticamente (Ehrendorfer 1965). Apesar de S. paniculatum ser uma espécie nanofanerófita, a técnica de controle através de roçadas no nível do solo pode ter selecionado uma estratégia semelhante à de plantas hemicriptófitas e caméfitas pela possibilidade de rebrota em época favorável, após a roçada. Assim, poderia ser vantajoso à espécie manter alta variabilidade através do cruzamento entre plantas, que lhes conferiria considerável flexibilidade às condições do ambiente em anos consecutivos. Espécies pioneiras de florestas tropicais, perenes, também são predominantemente alógamas (Bawa 1974).
Na população I, a baixa porcentagem de produção de frutos em condições naturais (17%) e a total ausência no teste com polinização cruzada (tabela 2) poderiam ser explicadas pelo excepcional e longo período de seca que ocorreu no ano de 1985 em todo estado de São Paulo (Camargo et al. 1986). Além disso, alguns dias com altas temperaturas devem ter agravado a deficiência hídrica no solo, implicando na queda de flores.
Outra hipótese que poderia explicar a não formação de frutos na população I seria a ocorrência de distúrbios meióticos, o que levaria à formação de gametas não funcionais. Entretanto, o processo meiótico nas diversas células pareceu bastante regular, tanto no pareamento como na disjunção dos cromossomos. O número cromossômico obtido (n=12) confirma contagens anteriores para outras populações desta espécie, nos estados da Bahia (Coleman & Smith 1969) e de São Paulo (Coleman 1982). A alta viabilidade de pólen (acima de 86%, tabela 2), tanto em flores C como em flores L, confirma indiretamente a regularidade da meiose.
As características morfológicas e comportamentais apresentadas pelas abelhas visitantes de flores de Solanum paniculatum coincidem com as relacionadas à síndrome da polinização vibrátil (Bowers 1975, Buchmann et al. 1977, Symon 1979, Coleman & Coleman 1982, Buchmann 1983).
A ocorrência de polinizadores distintos (exceto Bombus morio, mas com freqüência de visitas diferente) e de tamanhos diferentes nas duas populações (tabela 3) pode ser explicado pelo fato de que nas proximidades da população II ocorriam outras espécies de Solanum (S. mauritianum e S. ciliatum), cujas flores são maiores e mais vigorosas que as de S. paniculatum, que também eram visitadas por abelhas grandes como Bombus morio. A competição das três espécies de Solanum pelos polinizadores parece ser diminuída pelo deslocamento preferencial de abelhas grandes (como Bombus morio e outras) para espécies com flores maiores, com maior quantidade de pólen, enquanto as abelhas menores (Augochloropsis sp 1 e Augochloropsis sp 2) procuram flores menores, como as de S. paniculatum.
Agradecimentos - Aos Profs. Luiz Fernando Rödder e Moreira Aguiar, Pe. Jesus Santiago Moure e à Profa. Maria Cristina Almeida pelas identificações dos Hymenoptera, ao Prof. João Augusto Alves Meira e ao Dr. Condorcet Aranha pela identificação do material botânico e à Dra. Marlies Sazima e Dr. Ivan Sazima pela orientação durante a execução do trabalho.
Referências bibliográficas
ANDERSON, G.J. 1979. Dioecious Solanum species of hermaphroditic origin is an example of a broad convergence. Nature 282:836-838.
ANDERSON, G.J. & SYMON, D.E. 1989. Functional dioecy and andromonoecy in Solanum. Evolution 43:204-219.
BAKSH, S. & IQBAL, M. 1978. Floral features of Solanum macranthum Dun. with special reference to stylar heteromorphism and intercrossability. Flora 167:423-431.
BAWA, K.S. 1974. Breeding systems of tree species of a lowland tropical community. Evolution 28:85-92.
BOWERS, K.A.W. 1975. The pollination of Solanum rostratum (Solanaceae). Am. J. Bot. 62:633-638.
BUCHMANN, S.L. 1983. Buzz pollination in angiosperms. In Handbook of experimental pollination biology (C.E. Jones & R.J. Litter, eds). Van Nostrand & Reinhold, New York, p.73-113.
BUCHMANN, S.L., JONES, C.E. & COLIN, L.J. 1977. Vibratile pollination of Solanum douglassii and S. xanti (Solanaceae) in Southern California. Wasmann J. Biol. 35:1-25.
CAMARGO, M.B.P., ALFONSI, R.R., ORTOLANI, A.A., PEDRO-JÚNIOR, M.J., BRUNINI, O. & CHIAVEGA-TO, O.M.D.P. 1986. A seca de 1985 no estado de São Paulo. Agronômico 38:31-44.
COLEMAN, J.R. 1982. Chromossome numbers of angiosperms collected in the stat of São Paulo. Rev. Brasil. Genet. 5:533-549.
COLEMAN, J.R. & COLEMAN, M.A. 1982. Reproductive biology of an andromonoecious Solanum. (S. palinacan-thum Dunal). Biotropica 14:69-75.
COLEMAN, J.R. & SMITH, L.B. 1969. Chromossome numbers of some Brazilian angiosperms. Rhodora 71:548-551.
EHRENDORFER, F. 1965. Dispersal mechanisms, genetic systems, and colonizing abilites in some flowering plant families. In The genetics of colonizing species (H.G. Baker. & G.L. Stebbins, eds.). Academic Press, New York, p.331-351.
FAEGRI, K. & VAN DER PIJL, L. 1971. The principles of pollination ecology. Pergamon Press, New York.
GARDÉ, N.M. 1959. Mechanisms of species isolation in tuberous Solanum. Agron. Lusit. 21:19-42.
GRANT, V. 1975. Genetics of flowering plants. Columbia University Press, New York.
GRUN, P. 1961. Early stages in the formation of internal barriers to gene exchange between diploid species of Solanum. Am. J. Bot. 48:79-89.
HESLOP-HARRISON, J. 1975. Incompatibility and the pollen-stigma interaction. Ann. Rev. Plant Physiol. 26:403-425.
HESS, D. 1983. Die Blüte. Eugen Ulmer, Stuttgart.
HOSSAIN, F.L.S. M. 1973. Observations on stylar heteromorphism in Solanum torvum Sw. (Solanaceae). Bot. J. Linn. Soc. 66:291-301.
KORNERUP, A. & WANSCHER, J.H. 1963. Taschenlexikon der Farben. Ed. Musterschmidt, Zurich.
LEITÃO FILHO, H.F., ARANHA, C. & BACCHI, O. 1975. Plantas invasoras de culturas no estado de São Paulo. Vol. 2. HUCITEC e AGIPLAN, São Paulo.
MARTIN, F.W. 1959. Staining and observing pollen tubes in the style by means of fluorescence. Stain Technology 37:125.
MEDINA, D.M. & CONAGIN, C.H.T.M. 1964. Técnica citológica. Publicação nº 1610, Instituto Agronômico, Campinas.
MUELLER-DOMBOIS, H. & ELLENBERG, D. 1974. Aims and methods of vegetation ecology. Wiley, New York.
OLIVEIRA-FILHO, A.T. & OLIVEIRA, L.C.A. 1988. Biologia floral de uma população de Solanum lycocarpum St. Hil. (Solanaceae) em Lavras, MG. Revta brasil. Bot. 11:23-32.
PANDEY, K.K. 1960. Evolution of gametophytic and sporophytic systems of self-incompatibility in angiosperms. Evolution 14:98-115.
PIO CORRÊA, M. 1969. Dicionário das plantas úteis do Brasil e das exóticas cultivadas. Vol 4. Ministério da Agricultura, IBDF. Rio de Janeiro.
SENDTNER, O. 1846. Solanaceae. In Flora Brasiliensis (C.F.P. von Martius, ed.). F. Fleischer, Lipsiae. Vol 10.
SYMON, D.E. 1979. Sex forms in Solanum (Solanaceae) and the role of pollen collecting insects. In The biology and taxonomy of the Solanaceae (J.G. Hawkes, R.N. Lester & A.D. Skelding, eds.). Academic Press, London, p.385-397.
VOGEL, S. 1950. Farbwechsel und Zeichnungsmuster bei blüten. Öesterr. Bot. Z. 97:44-100.
VOGEL, S. 1962. Die Duftdrüsen der Orchideen. Akad. Wiss. Lit. Abh-Math.-Naturwiss-Kl. 10:603-763.
VOGEL, S. 1978. Evolutionary shifts from reward to deception in pollen flowers. In The pollination of flower by insects (A.J. Richards, ed.). Linnean Society Symposium Series. Academic Press, London. Vol. 6:89-96.
VOGEL, S. 1983. Ecophysiology of zoophilic pollination. In Physiological plant ecology III. (O.L. Lange, P.S. Nobel, C.B. Osmond & H. Ziegler, eds.). Springer-Verlag, Berlin, p. 560-623.
WHALEN, M.D. & ANDERSON, G.J. 1981. Distribution of gametophytic self-incompatibility and infrageneric classification in Solanum. Taxon 30:761-767.
WILLIS, J.C. 1973. A dictionary of flowering plants and ferns. University Press, Cambridge.
ZEISLER, M. 1938. Über die Abgrenzung der eigentlichen Narbenflche mit Hilf von Reaktionen. Beih. bot. Zbl. 58:308-318.
ZURLO, C. & BRANDÃO, M. 1990. As ervas comestíveis: descrição, ilustração e receitas. Editora Globo, São Paulo.
2. Departamento de Botânica, Setor de Ciências Biológicas, Universidade Federal do Paraná, Caixa Postal 19031, 81531-970 Curitiba, PR, Brasil.
3. Departamento de Ecologia, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478, 69011-970 Manaus, AM, Brasil.
- ANDERSON, G.J. 1979. Dioecious Solanum species of hermaphroditic origin is an example of a broad convergence. Nature 282:836-838.
- ANDERSON, G.J. & SYMON, D.E. 1989. Functional dioecy and andromonoecy in Solanum Evolution 43:204-219.
- BAKSH, S. & IQBAL, M. 1978. Floral features of Solanum macranthum Dun. with special reference to stylar heteromorphism and intercrossability. Flora 167:423-431.
- BAWA, K.S. 1974. Breeding systems of tree species of a lowland tropical community. Evolution 28:85-92.
- BOWERS, K.A.W. 1975. The pollination of Solanum rostratum (Solanaceae). Am. J. Bot. 62:633-638.
- BUCHMANN, S.L., JONES, C.E. & COLIN, L.J. 1977. Vibratile pollination of Solanum douglassii and S. xanti (Solanaceae) in Southern California. Wasmann J. Biol. 35:1-25.
- CAMARGO, M.B.P., ALFONSI, R.R., ORTOLANI, A.A., PEDRO-JÚNIOR, M.J., BRUNINI, O. & CHIAVEGA-TO, O.M.D.P. 1986. A seca de 1985 no estado de Săo Paulo. Agronômico 38:31-44.
- COLEMAN, J.R. 1982. Chromossome numbers of angiosperms collected in the stat of Săo Paulo. Rev. Brasil. Genet. 5:533-549.
- COLEMAN, J.R. & COLEMAN, M.A. 1982. Reproductive biology of an andromonoecious Solanum (S. palinacan-thum Dunal). Biotropica 14:69-75.
- COLEMAN, J.R. & SMITH, L.B. 1969. Chromossome numbers of some Brazilian angiosperms. Rhodora 71:548-551.
- EHRENDORFER, F. 1965. Dispersal mechanisms, genetic systems, and colonizing abilites in some flowering plant families. In The genetics of colonizing species (H.G. Baker. & G.L. Stebbins, eds.). Academic Press, New York, p.331-351.
- FAEGRI, K. & VAN DER PIJL, L. 1971. The principles of pollination ecology. Pergamon Press, New York.
- GARDÉ, N.M. 1959. Mechanisms of species isolation in tuberous Solanum Agron. Lusit. 21:19-42.
- GRANT, V. 1975. Genetics of flowering plants. Columbia University Press, New York.
- GRUN, P. 1961. Early stages in the formation of internal barriers to gene exchange between diploid species of Solanum Am. J. Bot. 48:79-89.
- HESLOP-HARRISON, J. 1975. Incompatibility and the pollen-stigma interaction. Ann. Rev. Plant Physiol. 26:403-425.
- HESS, D. 1983. Die Blüte. Eugen Ulmer, Stuttgart.
- HOSSAIN, F.L.S. M. 1973. Observations on stylar heteromorphism in Solanum torvum Sw. (Solanaceae). Bot. J. Linn. Soc. 66:291-301.
- KORNERUP, A. & WANSCHER, J.H. 1963. Taschenlexikon der Farben. Ed. Musterschmidt, Zurich.
- LEITĂO FILHO, H.F., ARANHA, C. & BACCHI, O. 1975. Plantas invasoras de culturas no estado de Săo Paulo. Vol. 2. HUCITEC e AGIPLAN, Săo Paulo.
- MARTIN, F.W. 1959. Staining and observing pollen tubes in the style by means of fluorescence. Stain Technology 37:125.
- MEDINA, D.M. & CONAGIN, C.H.T.M. 1964. Técnica citológica. Publicaçăo nş 1610, Instituto Agronômico, Campinas.
- MUELLER-DOMBOIS, H. & ELLENBERG, D. 1974. Aims and methods of vegetation ecology. Wiley, New York.
- OLIVEIRA-FILHO, A.T. & OLIVEIRA, L.C.A. 1988. Biologia floral de uma populaçăo de Solanum lycocarpum St. Hil. (Solanaceae) em Lavras, MG. Revta brasil. Bot. 11:23-32.
- PANDEY, K.K. 1960. Evolution of gametophytic and sporophytic systems of self-incompatibility in angiosperms. Evolution 14:98-115.
- PIO CORRĘA, M. 1969. Dicionário das plantas úteis do Brasil e das exóticas cultivadas. Vol 4. Ministério da Agricultura, IBDF. Rio de Janeiro.
- SENDTNER, O. 1846. Solanaceae. In Flora Brasiliensis (C.F.P. von Martius, ed.). F. Fleischer, Lipsiae. Vol 10.
- SYMON, D.E. 1979. Sex forms in Solanum (Solanaceae) and the role of pollen collecting insects. In The biology and taxonomy of the Solanaceae (J.G. Hawkes, R.N. Lester & A.D. Skelding, eds.). Academic Press, London, p.385-397.
- VOGEL, S. 1950. Farbwechsel und Zeichnungsmuster bei blüten. Öesterr. Bot. Z. 97:44-100.
- VOGEL, S. 1962. Die Duftdrüsen der Orchideen. Akad. Wiss. Lit. Abh-Math.-Naturwiss-Kl. 10:603-763.
- VOGEL, S. 1978. Evolutionary shifts from reward to deception in pollen flowers. In The pollination of flower by insects (A.J. Richards, ed.). Linnean Society Symposium Series. Academic Press, London. Vol. 6:89-96.
- VOGEL, S. 1983. Ecophysiology of zoophilic pollination. In Physiological plant ecology III. (O.L. Lange, P.S. Nobel, C.B. Osmond & H. Ziegler, eds.). Springer-Verlag, Berlin, p. 560-623.
- WHALEN, M.D. & ANDERSON, G.J. 1981. Distribution of gametophytic self-incompatibility and infrageneric classification in Solanum. Taxon 30:761-767.
- WILLIS, J.C. 1973. A dictionary of flowering plants and ferns. University Press, Cambridge.
- ZEISLER, M. 1938. Über die Abgrenzung der eigentlichen Narbenflche mit Hilf von Reaktionen. Beih. bot. Zbl. 58:308-318.
- ZURLO, C. & BRANDĂO, M. 1990. As ervas comestíveis: descriçăo, ilustraçăo e receitas. Editora Globo, Săo Paulo.
Datas de Publicação
-
Publicação nesta coleção
21 Dez 1998 -
Data do Fascículo
Ago 1998
Histórico
-
Aceito
03 Fev 1998 -
Recebido
24 Abr 1997