Resumos
(Aspectos estruturais da comunidade arbórea em remanescentes de floresta estacional decidual, em Corumbá, MS, Brasil). Um estudo comparativo da estrutura da comunidade arbórea entre dois remanescentes de floresta decídua foi conduzido em diferentes altitudes: florestas deciduais de terras baixas (FEDTB) e submontana (FEDSM), localizados em Corumbá, MS, Centro Oeste brasileiro. Amostraram-se indivíduos arbóreos com CAP ³ 15 cm, utilizando-se método de quadrantes. Foram demarcados 80 pontos em FEDTB e em FEDSM 78 pontos foram distribuídos por altitude: 180 m (18 pontos), 220, 260 e 300 m de altitude (20 pontos cada). Em FEDTB foram amostradas 34 espécies, sendo que Calycophyllum multiflorum Griseb., Ceiba pubiflora (A. St.-Hil.) K. Schum. e Anadenanthera colubrina (Vell.) Brenan apresentaram os maiores valores de importância. Na área de FEDSM foram amostradas 33 espécies, sendo que Anandenanthera colubrina, Ceiba pubiflora e Acosmium cardenasii H. S. Irwin & Arroyo foram as mais importantes. O padrão de distribuição das espécies variou ao longo do gradiente altitudinal. Em ambas as áreas, o índice de diversidade de Shannon foi de 2,9 e a Equitabilidade de 0,8, onde as famílias mais representativas foram Fabaceae (8 spp.) e Rubiaceae (4 spp). As florestas estudadas apresentaram baixa densidade de indivíduos por hectare em relação a outros estudos, exceto na área a 300 m. A comunidade arbórea apresentou 22,46% de indivíduos perfilhados e em fases iniciais de sucessão. O dossel variou de 6 a 12 m, com indivíduos emergentes de até 18 m. Estes remanescentes representam fonte de diversidade biológica para a região do Pantanal sendo importantes componentes dos corredores naturais da região.
composição florística; fitossociologia; floresta decídua; gradiente altitudinal
(Structural features of the arboreal community in remnants of dry forest in Corumbá, MS, Brazil). The structure of the arboreal community of two types of dry forest was compared in an altitudinal gradient of lowland (FEDTB) and submontane (FEDSM) dry forests, in Corumbá, MS, Midwest Brazil. Tree species with circumference at breast height ³ 15 cm were sampled, using the point centered quarter method. In the FEDTB 80 points were demarcated, and in FEDSM 78 points were distributed in the following altitudes: 180 m (18 points), 220, 260 and 300 m (20 points each). In FEDTB 34 species were sampled; Calycophyllum multiflorum Griseb., Ceiba pubiflora (A. St.-Hil.) K. Schum. and Anadenanthera colubrina (Vell.) Brenan have the highest VI values. In FEDSM, 33 species were sampled, and Anandenanthera colubrina, Ceiba pubiflora and Acosmium cardenasii H. S. Irwin & Arroyo were the most important. The distribution pattern of the species varied along the altitudinal gradient. In both forests Shannon's diversity and equitability indexes were 2.9 and 0.8, respectively, and the most representative families were Fabaceae (8 species) and Rubiaceae (4 species). The studied forests have low density of individuals in relation to other studies, except for the track at 300 m. About 22.46% of the individuals of arboreal community had basal ramifications and were in the initial phases of succession. The canopy varied from 6 to 12 m, and emergent individuals of up to 18 m. These remnants represent a source of biodiversity for the Pantanal, being an important part of the natural forest corridors of the region.
altitudinal gradient; deciduous forest; floristic compostition; phytosociology
ARTIGO DE REVISÃO
Aspectos estruturais da comunidade arbórea em remanescentes de floresta estacional decidual, em Corumbá, MS, Brasil1 1 Parte da dissertação de mestrado da primeira autora, Programa de Pós-Graduação em Biologia Vegetal da Universidade Federal de Mato Grosso do Sul, Campo Grande, MS, Brasil. 2 Autor para correspondência: geraldodamasceno@gmail.com
Michele Soares de LimaI; Geraldo Alves Damasceno-JúniorI,2 1 Parte da dissertação de mestrado da primeira autora, Programa de Pós-Graduação em Biologia Vegetal da Universidade Federal de Mato Grosso do Sul, Campo Grande, MS, Brasil. 2 Autor para correspondência: geraldodamasceno@gmail.com ; Marcel Okamoto TanakaII
IUniversidade Federal de Mato Grosso do Sul, Centro de Ciências Biológicas e de Saúde, Departamento de Biologia, Caixa Postal 549, 79070-900 Campo Grande, MS, Brasil
IIUniversidade Federal de São Carlos, Departamento de Hidrobiologia, Caixa Postal 676, 13565-905 São Carlos, SP, Brasil
ABSTRACT
(Structural features of the arboreal community in remnants of dry forest in Corumbá, MS, Brazil). The structure of the arboreal community of two types of dry forest was compared in an altitudinal gradient of lowland (FEDTB) and submontane (FEDSM) dry forests, in Corumbá, MS, Midwest Brazil. Tree species with circumference at breast height ³ 15 cm were sampled, using the point centered quarter method. In the FEDTB 80 points were demarcated, and in FEDSM 78 points were distributed in the following altitudes: 180 m (18 points), 220, 260 and 300 m (20 points each). In FEDTB 34 species were sampled; Calycophyllum multiflorum Griseb., Ceiba pubiflora (A. St.-Hil.) K. Schum. and Anadenanthera colubrina (Vell.) Brenan have the highest VI values. In FEDSM, 33 species were sampled, and Anandenanthera colubrina, Ceiba pubiflora and Acosmium cardenasii H. S. Irwin & Arroyo were the most important. The distribution pattern of the species varied along the altitudinal gradient. In both forests Shannon's diversity and equitability indexes were 2.9 and 0.8, respectively, and the most representative families were Fabaceae (8 species) and Rubiaceae (4 species). The studied forests have low density of individuals in relation to other studies, except for the track at 300 m. About 22.46% of the individuals of arboreal community had basal ramifications and were in the initial phases of succession. The canopy varied from 6 to 12 m, and emergent individuals of up to 18 m. These remnants represent a source of biodiversity for the Pantanal, being an important part of the natural forest corridors of the region.
Key words: - altitudinal gradient, deciduous forest, floristic compostition, phytosociology
RESUMO
(Aspectos estruturais da comunidade arbórea em remanescentes de floresta estacional decidual, em Corumbá, MS, Brasil). Um estudo comparativo da estrutura da comunidade arbórea entre dois remanescentes de floresta decídua foi conduzido em diferentes altitudes: florestas deciduais de terras baixas (FEDTB) e submontana (FEDSM), localizados em Corumbá, MS, Centro Oeste brasileiro. Amostraram-se indivíduos arbóreos com CAP ³ 15 cm, utilizando-se método de quadrantes. Foram demarcados 80 pontos em FEDTB e em FEDSM 78 pontos foram distribuídos por altitude: 180 m (18 pontos), 220, 260 e 300 m de altitude (20 pontos cada). Em FEDTB foram amostradas 34 espécies, sendo que Calycophyllum multiflorum Griseb., Ceiba pubiflora (A. St.-Hil.) K. Schum. e Anadenanthera colubrina (Vell.) Brenan apresentaram os maiores valores de importância. Na área de FEDSM foram amostradas 33 espécies, sendo que Anandenanthera colubrina, Ceiba pubiflora e Acosmium cardenasii H. S. Irwin & Arroyo foram as mais importantes. O padrão de distribuição das espécies variou ao longo do gradiente altitudinal. Em ambas as áreas, o índice de diversidade de Shannon foi de 2,9 e a Equitabilidade de 0,8, onde as famílias mais representativas foram Fabaceae (8 spp.) e Rubiaceae (4 spp). As florestas estudadas apresentaram baixa densidade de indivíduos por hectare em relação a outros estudos, exceto na área a 300 m. A comunidade arbórea apresentou 22,46% de indivíduos perfilhados e em fases iniciais de sucessão. O dossel variou de 6 a 12 m, com indivíduos emergentes de até 18 m. Estes remanescentes representam fonte de diversidade biológica para a região do Pantanal sendo importantes componentes dos corredores naturais da região.
Palavras-chave: - composição florística, fitossociologia, floresta decídua, gradiente altitudinal
INTRODUÇÃO
As florestas estacionais deciduais ocorrem na forma de disjunções florestais em regiões tropicais com precipitação anual abaixo de 2.000 mm e período de cinco a seis meses de estação seca, com precipitação total menor que 1.000 mm (Pennington et al. 2000), apresentando indivíduos com valores baixos na estatura e área basal (Linares-Palomino et al. 2003). Na América do Sul, são encontradas na forma de manchas e encraves remanescentes, de um período glacial frio e seco ocorrido entre o Pleistoceno e Holoceno (Prado & Gibbs 1993). Estas vegetações constituíam um corredor contínuo formado pelo núcleo da caatinga nordestina, núcleo das Misiones, que abrange a confluência dos Rios Paraguai e Paraná, e a depressão do Rio Uruguai, em Misiones (Argentina) e Santa Catarina (Brasil), e o terceiro núcleo Piedmont, que se extende de Santa Cruz de La Sierra na Bolívia a Tucumán e sierras ao noroeste da Argentina (Prado & Gibbs 1993). As formações deciduais localizadas nas morrarias de entorno do Município de Corumbá ocorrem na zona delimitada pelo núcleo Misiones, na fronteira com a Bolívia. Em todas estas áreas, ocorrem tipos vegetacionais complexos, dependendo do clima local, do substrato e do gradiente topográfico (Pennington et al. 2000).
Os fatores determinantes de vegetações decíduas, como sazonalidade da precipitação, características físicas do solo e topografia, são escassamente documentados e dificultam análises comparativas com outras áreas (Mooney et al. 1995). Poucos trabalhos florísticos e fitossociológicos são dirigidos a estas formações, devido em parte pela sua pobreza em espécies e a maior atenção conferida à rápida fragmentação de florestas tropicais úmidas (Mooney et al. 1995). Esta condição é lamentável porque as florestas sazonalmente secas vêm sofrendo grandes taxas de desmatamento para uso humano e possuem importante valor potencial como banco genético de espécies, sendo ecossistemas conhecidos apenas pela sua distribuição nos neotrópicos (Ratter et al. 1988, Mooney et al. 1995).
Poucos trabalhos abordam as variações da vegetação ao longo de um gradiente altitudinal em matas decíduas. Alguns estudos indicam que a altitude (Vásquez & Givnish 1998), a topografia e o substrato (Prance & Schaler 1982, Salis et al. 2004, White & Hood 2004) afetam claramente a abundância e distribuição de espécies lenhosas, indicando que estes fatores influenciam fortemente nos padrões da vegetação. Uma característica muito presente observada nestas formações é a grande predominância de espécies anemocóricas, cuja dispersão é facilitada pela deciduidade foliar das espécies na época de estiagem (Pennington et al. 2000). Porém, há poucos estudos focados na variação florística e estrutural das florestas decíduas de terras baixas e submontana em gradiente altitudinal, bem como nas estratégias de dispersão das espécies na estrutura vertical.
O rápido processo de urbanização, a ocupação ilegal dos morros, a retirada de calcário e construção de rodovias sem um plano de recomposição vêm agravando a taxa de desmatamento da vegetação no Município de Corumbá, e o desconhecimento da importância desta vegetação se torna um agravante, pois o microclima proporcionado pela cobertura florestal ajuda a reduzir os efeitos da radiação solar e as altas temperaturas. Com o objetivo de subsidiar futuros estudos de conservação das espécies de florestas deciduais, e auxiliar na compreensão da distribuição e dinâmica deste ecossistema, este estudo buscou avaliar como variam a composição florística e estrutural da vegetação arbórea, assim como as estratégias de dispersão das espécies levantadas em cada estrato, em dois remanescentes de floresta estacional decidual de terras baixas e submontana.
Material e métodos
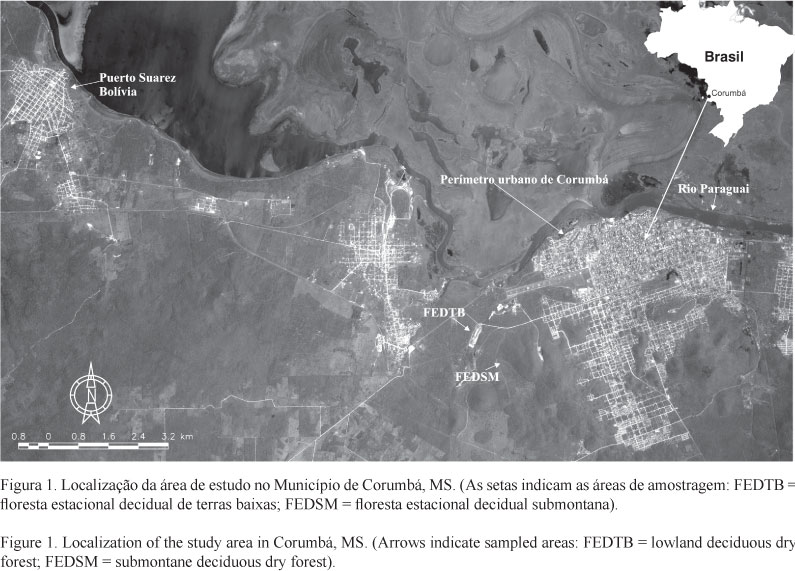
Áreas de estudo - As áreas de estudo estão localizadas cerca de 3 km a oeste do perímetro urbano do Município de Corumbá (19°01' S e 57°40' W), Estado de Mato Grosso do Sul, abrangendo uma vegetação de sopé (55 ha) e encosta (105 ha) que distam cerca de 200 metros entre si (figura 1). A primeira área, classificada como floresta estacional decidual de terras baixas - FEDTB (IBGE 1992), situa-se a 19°01'36,0" S e 57°41'03,4" W, com altitude máxima de 150 m acima do nível do mar, possui relevo plano a levemente ondulado, com poucos aglomerados rochosos de calcário e com uma camada de solo mais espessa. Encontra-se sobre solo do tipo rendzina de textura média a média pouco cascalhenta em fase endopedregosa, com relevo suave ondulado, declividade inferior a 2%, e altitude que varia entre 90 e 150 m (Spera et al. 1997, Okida & Anjos 2000). A segunda área é classificada como floresta estacional decidual submontana - FEDSM (IBGE 1992), está situada a 19°01'38,1" S e 57°40'57,2" W, com o topo atingindo 300 m acima do nível do mar. O solo constitui-se de cambissolo eutrófico chernozenico de textura média, com forte afloramento rochoso; sua declividade está entre 20% e 40%, com cota altimétrica que varia de 150 a 300 m (Spera et al. 1997, Okida & Anjos 2000).
O clima é do tipo Awa, segundo classificação de Köppen (1948), caracterizada por clima tropical de altitude, megatérmico (a temperatura média do mês mais frio é superior a 18 °C), com inverno seco e chuvas no verão, sendo que a média anual para a temperatura e precipitação registrada no período de 1975 a 1996 por Soriano (1997) foi de 25,1 °C e 1.070 mm, respectivamente. As estações são bem definidas: uma de outubro a janeiro, na qual as temperaturas médias absolutas mensais oscilam entre 26 °C e 28 °C, com máxima absoluta que pode chegar aos 40 °C; a outra estação, com temperaturas amenas, ocorre de maio a agosto, com temperaturas variando entre 21 °C e 25 °C, atingindo valores mais baixos nos meses de junho e julho, quando a mínima absoluta pode chegar a 0 °C. A maior concentração de chuvas ocorre de outubro a março, alcançando 68% do total anual, os meses mais secos correspondem ao período de junho a agosto, com uma precipitação menor que 7% da precipitação total anual (Soriano 1997).
Coleta dos dados - A amostragem foi realizada no mês de março de 2006, durante o período chuvoso, sendo utilizado o método de quadrantes centrados (Brower & Zar 1984). Foram amostrados todos os indivíduos arbóreos e arbustivos com pelo menos um dos troncos com CAP (circunferência à 1,3 m do solo) maior ou igual a 15 cm. Amostras dos indivíduos, férteis ou não, foram coletadas para proceder à identificação botânica. Em FEDTB, quatro amostras foram demarcadas aleatoriamente, por meio de picadas no meio da vegetação, com distância mínima de 100 metros entre si. A partir de 15 metros de distância da borda da mata, foram estabelecidos 20 pontos em cada amostra, distanciados 10 metros entre si, totalizando 80 pontos. Na área de FEDSM, com auxílio de um altímetro, também foram estabelecidas quatro amostras a partir da base do morro: a 180 metros (com 18 pontos), 220, 260 e 300 metros de altitude (20 pontos cada), totalizando 78 pontos demarcados. As três primeiras amostras se encontram em solo pouco profundo com grandes afloramentos carbonatados e declividade entre 10% e 20%. No topo do morro, um suave platô, encontra-se a amostra a 300 metros, caracterizado pela ocorrência de alguns vales úmidos de solo mais profundo, com vegetação densa e viçosa.
Para cada indivíduo amostrado anotou-se a espécie, o CAP (circunferência do caule a 1,3 m. acima do solo), a altura e a distância do indivíduo até o ponto. O material coletado foi identificado por comparação com exemplares já existentes e identificados no Herbário do Departamento de Ciências do Ambiente (CPAN/UFMS) e com auxílio de bibliografia especializada, sendo posteriormente incorporado à coleção do mesmo Herbário. A nomeação dos táxons seguiu o sistema de classificação do Angiosperm Phylogeny Group (APG II 2003). Os exemplares coletados férteis encontram-se depositados no herbário COR.
Os parâmetros fitossociológicos utilizados, baseados em Martins (1993), foram: densidade absoluta (DA) e relativa (DR), dominância absoluta (DoA) e relativa (DoR), freqüência absoluta (FA) e relativa (FR), Índice de Valor de Importância (VI) e Índice de Valor de Cobertura (VC). Os cálculos foram realizados com uso do programa FITOPAC 1.0 (Shepherd 1994). Foram calculados também os índices de diversidade de Shannon (Brower & Zar 1984) e Equitabilidade (Pielou 1975). A curva do coletor foi obtida para ambas as áreas, listando-se o número de indivíduos amostrados para cada espécie obtida, aleatoriamente (Pielou 1975).
Para analisar a existência de variação na distribuição das espécies, utilizou-se a análise de componentes principais (PCA) obtendo-se uma matriz de número de indivíduos para as quatro amostras de floresta decidual submontana, situadas em gradiente altitudinal, e para a floresta decidual de terras baixas, que foi decomposta em quatro amostras de 20 pontos para equilibrar a análise. No PCA foi utilizada a matriz de co-variância, a fim de que os dados quantitativos tivessem maior peso no resultado final. Para complementar a avaliação das associações florísticas entre as amostras, realizou-se análise de agrupamento utilizando-se o coeficiente de Bray-Curtis, por este ser mais sensível aos dados quantitativos, e o método de agrupamento de média de grupo (UPGMA). As análises foram feitas utilizando-se os programas PC-ORD for Windows, versão 3.17 (McCune & Mefford 1997) e FITOPAC 1.0 (Shepherd 1994).
Para a análise da distribuição diamétrica das espécies, transformou-se o CAP de cada indivíduo para diâmetro, utilizando-se a fórmula:
Diam = CAP π-1
a seguir, utilizou-se a fórmula para cálculo mínimo das classes e sua amplitude, proposto por Spiegel (1977).
O cálculo para a taxa de perfilhamento só foi obtido para as espécies que apresentaram no mínimo 10 indivíduos amostrados, considerando conjuntamente as duas áreas. Para o delineamento dos estratos florestais, a amplitude e a mediana das alturas foram representadas em um gráfico apenas para as espécies com no mínimo três indivíduos amostrados. A classificação das síndromes de dispersão foi efetuada de acordo com van der Pijl (1972).
Resultados
Composição florística e diversidade - Nas duas áreas foram amostrados 632 indivíduos arbóreos e arbustivos, distribuídos em 42 espécies, 36 gêneros e 22 famílias (tabela 1). A riqueza observada para as duas áreas de floresta de terras baixas e submontana foi de 34 e 33 espécies, respectivamente. A curva de coletor indicou uma diminuição no incremento de espécies a partir de 20 pontos amostrados, na qual constava cerca de 81% e 73% do total de espécies amostradas em floresta de terras baixas e submontana, respectivamente. As amostragens obtidas foram consideradas satisfatórias para comparar a composição florística e a estrutura das duas florestas. A área com remanescente de floresta decidual de terras baixas apresentou índice de diversidade de Shannon H' = 2,905 e Equitabilidade de Pielou J' = 0,824, enquanto que a área com floresta decidual submontana apresentou diversidade de Shannon H' = 2,933 e Equitabilidade de Pielou J' = 0,839 (tabela 2). No geral, as famílias mais representativas foram Fabaceae (oito espécies), Rubiaceae (quatro espécies), Malvaceae, Apocynaceae, Euphorbiaceae, Brassicaceae e Nyctaginaceae com três espécies cada (tabela 1). Os gêneros com maior número de espécies foram Aspidosperma (Apocynaceae), Capparis (Brassicaceae) e Acacia (Fabaceae), com três espécies cada. A família Fabaceae, a mais representativa em número de espécies (sete em cada área), constituiu 24,7% e 34,3% dos indivíduos amostrados em floresta decidual de terras baixas e submontana, respectivamente. Rubiaceae e Malvaceae (três espécies cada) também foram bastante abundantes, constituindo cerca de 28,4% e 8,4% em floresta de terras baixas, e em floresta submontana, 16% e 9,9%, respectivamente. Em ambas as áreas, a maior porcentagem de Fabaceae foi devido à elevada quantidade de indivíduos de Anadenanthera colubrina, Acosmium cardenasii e Acacia paniculata. Rubiaceae foi caracterizada por Calycophyllum multiflorum e Simira corumbensis, mas a primeira foi encontrada em grande quantidade em floresta submontana, constituindo a maior população desta área.
Ordenamento dos dados e similaridade entre amostras - A distribuição das espécies variou ao longo do gradiente altitudinal. Os autovalores do eixo 1 e 2 explicaram cerca de 42,7% e 20% da variação total dos dados, respectivamente (figura 2). O eixo 1 indica uma separação entre as áreas em dois grupos distintos, influenciadas principalmente pela variação do gradiente altitudinal. As amostras situadas a 180 (SM1), 220 (SM2) e 260 metros (SM3) de altitude se assemelham pela abundância de algumas espécies como Anadenanthera colubrina e Myracrodruon urundeuva e a ocorrência rara de Amburana cearensis e Cereus bicolor, enquanto que a amostra situada a 300 m (SM4) exibiu uma composição de espécies variável e distinta das demais amostras em floresta decidual submontana, porém, floristicamente semelhante à floresta decidual de terras baixas. Apenas uma amostra em terras baixas denominada TB4 foi diferente de todas as demais, apresentando a maior variação em relação ao eixo 2, devido principalmente à grande predominância de indivíduos de Capparis retusa e à ocorrência exclusiva de um único indivíduo de Ximenia americana. A análise de agrupamento (figura 3) confirma a similaridade florística entre a amostra a 300 m (SM4) com a floresta decidual de terras baixas, e a segregação desta com as amostras em floresta decidual submontana. A análise cofenética entre as áreas foi de 0,75.
Caesalpinia pluviosa e Sterculia striata foram as únicas espécies com mais de três indivíduos amostrados exclusivamente em floresta de terras baixas. Outras ocorreram somente em floresta submontana, como Bougainvillea infesta, Cereus bicolor e Saccelium brasiliense. Algumas áreas apresentaram espécies exclusivas, porém com poucos indivíduos amostrados, como Pereskia sacharosa e Ficus calyptroceras na amostra a 260 m, e Capparis prisca, Cedrela fissilis e Randia armata em amostra a 300 m. Algumas espécies mostraram-se abundantes em ambas florestas deciduais, como Acacia paniculata, Acosmium cardenasi, Anadenanthera colubrina, Aspidosperma pyrifolium, Aspidosperma quirandy, Calycophyllum multiflorum,Capparis retusa, Ceiba pubiflora, Cnidoscolus cnicodendron, Pseudobombax cf. marginatum, Simira corumbensis e Tabebuia impetiginosa.
Fisionomia e estrutura horizontal - Em floresta decidual de terras baixas, a densidade total foi de 916 indivíduos ha-1, com intervalo de confiança de 820 a 1.020 indivíduos ha-1, e uma estimativa de 32,14 m2 de área basal por hectare; na área de floresta decidual submontana, a densidade foi de 963 indivíduos ha-1, com intervalo de confiança de 867 a 1.070 indivíduos ha-1, com 38,31 m2 de área basal por hectare. Em gradiente altitudinal, a amostra a 300 m apresentou uma densidade total de 1.130 indivíduos ha-1, superior às outras amostras.
As espécies que obtiveram maior VI na área de floresta de terras baixas (tabela 3) foram Calycophyllum multiflorum (40,12), Ceiba pubiflora (35,15), Anadenanthera colubrina (29,25), Tabebuia impetiginosa (26,68), Simira corumbensis (23,15) e Acosmium cardenasii (20,64), que juntas, somaram 58,33% do VI total. O alto VI de Calycophyllum multiflorum se deve a maior freqüência relativa, mas se for analisado apenas o índice de cobertura, Ceiba pubiflora aparece em primeiro lugar, devido a alta dominância relativa (29,00) em relação a C. multiflorum (8,81). As outras espécies somam 35,98% do total. Do ponto de vista de contribuição de VI por família, Fabaceae (24,49%), Rubiaceae (20,94%) e Malvaceae (17,24%) somaram 62,67% do VI total.
Em floresta submontana (tabela 4), Anadenanthera colubrina (50,80) e Ceiba pubiflora (43,33) somaram 31,37% do VI total, devido ao alto valor de dominância relativa, apesar de C. pubiflora apresentar um baixo valor de freqüência relativa. Em relação ao VC, A. colubrina e C. pubiflora mantiveram-se nas mesmas posições. As famílias Fabaceae (29,21%), Malvaceae (20,78%) e Rubiaceae (11,52%) também foram as mais importantes nas florestas de encosta.
Em floresta decidual submontana, foi observada uma variação na importância das espécies, de acordo com o gradiente altitudinal. Anadenanthera colubrina aparece entre as principais espécies com maior VI nos trechos em 180, 220 e 260 metros de altitude, principalmente devido à grande quantidade de indivíduos, seguido por Acosmium cardenasii e Ceiba pubiflora. Esta última, apesar da sua baixa densidade nos trechos de 180 e 220 metros, apresentou VI alto devido à sua grande área basal, decrescendo no trecho a 260 metros. A 300 metros de altitude, A. colubrina cede lugar como espécie mais importante a C. pubiflora e Simira corumbensis, este último devido à alta densidade na área.
As comunidades vegetais nas florestas deciduais de terras baixas e submontana apresentaram uma distribuição das classes de diâmetro com grande predominância de indivíduos com diâmetro abaixo de 10 cm. Ao se comparar a figura 4 da distribuição diamétrica nas duas vegetações citadas com os dados de dominância e freqüência relativas, se observa que as duas áreas são compostas, na maioria, de indivíduos com 4 a 8 cm de diâmetro. Anadenanthera colubrina, Acosmium cardenasii e Simira corumbensis foram as que apresentaram menores valores de diâmetro em todas as áreas amostradas, sendo que na amostra a 300 m as espécies Calycophyllum multiflorum e Aspidosperma pyrifolium também incrementam o valor nas menores classes de diâmetro.
Perfilhamento " Uma característica marcante da comunidade lenhosa dos dois tipos de floresta decidual foi a grande quantidade de árvores perfilhadas, principalmente em Bougainvillea infesta e Jacaratia corumbensis (figura 5), seguidos de Capparis retusa, Cnidoscolus cnicodendron, Simira corumbensis e Aspidosperma pyrifolium, apresentando cada um mais de 30% dos indivíduos perfilhados. Cerca de 22,46% apresentaram perfilhamento, considerando os 632 indivíduos amostrados, com pelo menos dois perfilhos a 1,30 m à altura do solo. Os valores para a área de floresta de terras baixas (20,93%) e para submontana (24,03%) também se aproximaram desta taxa. Considerando apenas as espécies que apresentaram pelo menos 10 indivíduos, a taxa de perfilhamento variou de 4,16% a 90,90% por espécie, com média de 32% (figura 5).
Estrutura vertical e estratégias de dispersão " Nas duas áreas de floresta decídua foram identificados três estratos: sub-bosque (2,5 a 5 m de altura), composto principalmente por árvores jovens e arbustos como Simira corumbensis, Acacia paniculata, Capparis retusa, Urera baccifera e Allophylus pauciflorus; o dossel (6 a 12 metros), pouco contínuo e composto pela maioria das espécies amostradas, dos quais destacam-se principalmente Acosmium cardenasii, Calycophyllum multiflorum, Cnidoscolus cnicodendron, Aspidosperma pyrifolium e Jacaratia corumbensis; e o estrato emergente (13 a 18 metros), com a predominância de espécies como Tabebuia impetiginosa, Anadenanthera colubrina, Ceiba pubiflora, Pseudobombax cf. marginatum, Myracrodruon urundeuva e Enterolobium contortisiliquum (figura 6A, B).
A maioria das espécies ocorrentes nas duas florestas deciduais apresentou síndrome de dispersão anemocórica, com sua maior predominância no estrato superior da comunidade, abrangendo 100% das espécies emergentes da floresta decidual submontana (figura 6C, D). Por sua vez, a maior parte das espécies zoocóricas se concentrou no subdossel, tanto em floresta de terras baixas (26%) quanto em submontana (13%).
Discussão
A riqueza observada para as duas áreas de floresta estacional decidual de terras baixas e submontana (34 e 33 espécies, respectivamente) aproxima-se dos valores observados para outros remanescentes de floresta decídua estudados na região de Corumbá por Damasceno-Júnior & Bezerra (2004) e Salis et al. (2004) (tabela 2). Entretanto, está abaixo do valor de 50-70 espécies com DAP > 2,5 cm em amostras de 0,1 ha, proposto por Gentry (1995) para florestas estacionais deciduais, que são mais pobres em espécies, comparadas às florestas úmidas semi-sempreverdes (100-150 espécies) e florestas pluviais (200-250 espécies). Em algumas áreas onde se encontram fragmentos deste tipo de floresta, a riqueza pode chegar a mais de 100 espécies, como encontrado por Werneck et al. (2000) na região da Mata da Zilda (114 espécies lenhosas) no Município de Perdizes, MG, na qual a composição florística deve-se em grande parte à fase de sucessão em que a floresta se encontra, ocasionada por perturbações pela retirada seletiva de madeira.
Apesar de a curva do coletor indicar uma nítida tendência de estabilização, deve-se ressaltar que este método é controverso, uma vez que em florestas tropicais as complexas associações entre os componentes tornam a riqueza destes habitats variável no espaço-tempo; assim observa-se que o incremento no número de espécies em relação ao esforço amostral tende ao infinito, à medida em que se aumenta o tamanho amostral (Schilling & Batista 2008). Este fato foi constatado para algumas espécies como Jatropha weddelliana e Sapium cf. hasslerianum, que foram observadas em ambas as áreas, mas entraram apenas na relação de espécies da floresta estacional de terras baixas.
Em geral, a riqueza específica tende a diminuir com o aumento da altitude, como constatado por Silva & Scariot (2003) em uma área de floresta decidual sobre afloramento calcário na Bacia do Rio Paraná, cuja riqueza encontrada é menor do que em florestas deciduais de terras baixas (Scariot & Sevilha 2000). Em um estudo de campo conduzido por Damasceno-Júnior no planalto residual do Urucum, em Corumbá, MS, observou-se que a diversidade da vegetação diminui à medida que se avança em um gradiente altitudinal de 100 a 800 m, na qual as florestas decíduas da base da morraria foram mais ricas em espécies do que as florestas semideciduais das altitudes elevadas (dados não publicados). O mesmo padrão foi constatado por Vázquez & Givnish (1998) na Reserva de Sierra de Manantlán, México, onde a riqueza da floresta decídua diminuiu com o aumento da altitude. Diferentemente dos padrões acima citados, foi observada pouca variação no número de espécies nas amostras levantadas no presente estudo, devido em parte à relativa proximidade das áreas e ao baixo gradiente altitudinal entre as amostras.
Florestas estacionais deciduais na região de Piracicaba, SP (Ivanauskas & Rodrigues 2000), no Triângulo Mineiro, MG (Werneck et al. 2000), em fragmento de floresta estacional de terras baixas em Macaíba, RN (Cestaro & Soares 2004) e sobre afloramento calcário em Goiás (Silva & Scariot 2004) apresentaram H' que variou de 3,0 a 3,7, maiores que a diversidade encontrada em ambas florestas deciduais de Corumbá, no presente estudo. Apesar disso, o índice de Equitabilidade de 80% indica que os indivíduos arbóreo-arbustivos distribuem-se aleatoriamente na região.
A grande predominância da família Fabaceae, seguida por Rubiaceae, Malvaceae, Apocynaceae, Euphorbiaceae, Brassicaceae e Nyctaginaceae, bem como a ocorrência de espécies de ampla distribuição, como Anadenanthera colubrina, Myracrodruon urundeuva e Tabebuia impetiginosa coincidem com os resultados obtidos por Ratter et al. (1988) e Salis et al. (2004), cujos estudos também foram conduzidos em floresta decídua de Corumbá, MS. Gentry (1995) afirma que espécies componentes do grupo das Leguminosas (família com grande presença de espécies com frutos e sementes anemocóricos), assim como Euphorbiaceae (família predominantemente autocórica) e Brassicaceae, são bem representadas nestes tipos florestais. O forte estresse hídrico presente em vegetações deciduais limitaria a ocorrência de outras espécies sem mecanismos de compensação à esta restrição, estando assim ausentes ou mal representadas nesses ambientes. Diferenças na hidrologia, estrutura do solo ou exposição à luz solar direta podem influenciar a sobrevivência de plântulas e árvores, selecionando espécies capazes de tolerar esses habitats (Webb et al. 1999).
Algumas espécies foram exclusivas em determinadas amostras, como Pereskia sacharosa e Ficus calyptroceras em amostra a 260 m, e Capparis prisca, Cedrela fissilis e Randia armata em amostra a 300 m. Entretanto, esta particularidade pode ser efeito da insuficiência de indivíduos amostrados na tomada de dados. Espécies abundantes e de ampla ocorrência nas áreas como Acosmium cardenasii, Anadenanthera colubrina, Cnidoscolus cnicodendron, Myracrodruon urundeuva, Pseudobombax cf. marginatum e Tabebuia impetiginosa parecem ser indiferentes à presença de afloramentos rochosos. Killeen et al. (1998) estudando uma vegetação decídua em Las Trancas, em Chiquitania, na Bolívia, constataram, também por análise de PCA, que A. cardenasii, A. colubrina, T. impetiginosa, Cedrela fissilis e P. cf. marginatum foram consideradas características de áreas de vegetação decídua sob elevações.
Os valores de densidade absoluta e área basal encontrados nas florestas deciduais de terras baixas e submontana estão abaixo da média encontrada em estudos semelhantes (tabela 2) e podem ser reflexos da metodologia empregada no presente estudo, que exclui indivíduos mais jovens da amostragem. Outra explicação seria que, em florestas deciduais de encosta, com grande afloramento rochoso, o ambiente dificulta a fixação do sistema radicular e a exploração de recursos, o que resulta na seleção das espécies com vantagens competitivas suficientes para permanecer neste ambiente. Para o valor de densidade obtido na amostra a 300 m, o terreno pouco inclinado, com ocorrência de alguns vales que permite a maior deposição de solo favorece o desenvolvimento e a ocupação sucessiva das espécies, principalmente daquelas pouco tolerantes a ambientes com afloramento rochoso.
A predominância de Anadenanthera colubrina, Ceiba pubiflora, Calycophyllum multiflorum, Simira corumbensis, Tabebuia impetiginosa e Pseudobombax cf. marginatum, evidenciada nos valores de freqüência, densidade e dominância absolutas, difere dos resultados encontrados em estudos feitos em outras formações deciduais (Killeen et al. 1998, Damasceno-Júnior & Bezerra 2004). Acosmium cardenasii, espécie predominante nestes estudos, não foi muito representativa nos trechos altitudinais, variando em 5% a 11% no VI.
Os indivíduos que apresentaram maiores valores de dominância relativa foram representados por espécies como Pseudobombax cf. marginatum e Ceiba pubiflora ("paineiras" ou "barrigudas"), que possuem o caule com elevado estoque hídrico, pelas cactáceas com caule suculento e fotossintetizante, e por outras espécies com xilopódio, como Jacaratia corumbensis (Caricaceae). As diferenças apresentadas entre as florestas deciduais de terras baixas e submontana na distribuição dos valores de importância das espécies podem ser atribuídas às condições distintas de solo e topografia, favorecendo a predominância de algumas espécies em relação a outras em um tipo florestal específico. Em solos de encostas íngremes, que possuem boa drenagem e solo raso, espécies que toleram bem estresse à seca tendem a se sobressair em relação a espécies pouco tolerantes.
A floresta decidual submontana apresentou maior quantidade de indivíduos com 6 a 8 cm de diâmetro, devido à grande predominância de indivíduos jovens de Anadenanthera colubrina e outras de menor circunferência, como Acosmium cardenasii e Simira corumbensis. A distribuição típica em jota (J) invertido apresentada pela floresta decidual de terras baixas é atribuída ao grande número de arbustos adaptados a condições xerofíticas e de indivíduos jovens com diâmetro abaixo de 6 cm, como Calycophyllum multiflorum, Simira corumbensis, A. cardenasii e Cnidoscolus cnicodendron, que podem, portanto, constituir uma reserva para a continuidade da sucessão florestal. Esta característica decorre das condições extremas enfrentadas pelas espécies no início do seu desenvolvimento, como o solo raso e a deficiência hídrica, o que compromete o desenvolvimento da área basal. Nestas condições, espécies de matas secas calcárias alocam a maior parte dos recursos para o armazenamento de água no sistema radicular do que para a parte aérea (Mooney et al. 1995). Outras espécies, como Ceiba pubiflora e Pseudobombax cf. marginatum, desenvolvem mecanismos adaptativos à restrição hídrica, estocando reservas de água no seu caule, o que lhes confere grandes valores nas classes de circunferência.
A quantidade relativamente alta de indivíduos perfilhados (22%) se aproxima do valor encontrado por Ivanauskas & Rodrigues (2000) em uma floresta estacional decidual de planície em Piracicaba, SP, com 31% dos indivíduos com PAP ³ 15 cm apresentando perfilhamento; ao analisarem somente as espécies com pelo menos 10 indivíduos amostrados, a taxa de perfilhamento variou de 5% a 79%, com média de 33%. De acordo com Ceccon et al. (2006), a regeneração por brotamento direto de árvores adultas parece ser um padrão comum encontrado em florestas decíduas; devido à baixa sobrevivência das sementes neste ambiente, o brotamento vegetativo parece ser vantajoso para muitas espécies, mesmo aquelas que se mantém por sementes, porque os brotos vegetativos podem se beneficiar do sistema radical e da estocagem de metabólitos da planta-mãe.
Os estratos nas duas amostras de floresta decídua apresentaram alturas semelhantes encontradas em outros estudos, como Ivanauskas & Rodrigues (2000), Damasceno-Júnior & Bezerra (2004) e Salis et al. (2004). Acosmium cardenasii foi a principal espécie constituinte do dossel em estudo feito por Damasceno-Júnior & Bezerra (2004), em uma ilha na Lagoa do Castelo, Corumbá, que também possui vegetação decídua, ocorrendo também como espécie dominante do dossel de floresta decidual submontana, juntamente com Simira corumbensis. Entretanto, o dossel da floresta decidual de terras baixas se constituiu de outras espécies mais abundantes principalmente de Calycophyllum multiflorum e S. corumbensis, juntamente com A. cardenasii. Ratter et al. (1988) descreveram as áreas de morraria ao redor de Corumbá com vegetação decídua sobre afloramento calcário, cuja altura do dossel variou de 6 a 15 m, relativamente baixo se comparado ao dossel de florestas tropicais mais úmidas. Na região de Chiquitania, na Bolívia, a altura média de árvores amostradas em estudo conduzido por Killeen et al. (1998) foi de 16 m, e a maior altura registrada foi Schinopsis brasiliensis, com cerca de 30-35 m, um padrão muito superior se comparado à altura média de 8 m do total de espécies registrado nas duas florestas decíduas levantadas no presente estudo.
A predominância de espécies anemocóricas nas duas áreas já era esperada para este tipo de vegetação, com grande quantidade de espécies com esta característica no dossel e estrato emergente. Este padrão foi encontrado também por Killeen et al. (1998) para o componente arbóreo de uma vegetação decídua em Las Trancas, Bolívia, com predomínio de espécies anemocóricas no estrato emergente. Gentry (1995) enfatiza as duas principais tendências em florestas secas, comparadas às florestas úmidas e chuvosas: a grande prevalência de espécies lenhosas com floração curta e conspícua, polinizadas por insetos especialistas e com sementes anemocóricas. A escassez de espécies zoocóricas em florestas decíduas pode estar relacionada com a necessidade de maior investimento energético e hídrico por parte da planta-mãe na produção de frutos carnosos. Em espécies anemocóricas o investimento de energia na frutificação é baixo, uma vez que nestes habitats a estocagem de água e nutrientes é limitada e não há dependência por algum vetor animal (Janzen 1967). Na mata decídua da Lagoa do Castelo, em Corumbá, Damasceno-Júnior & Bezerra (2004) constataram que cerca de 50% das espécies eram zoocóricas e somente 33% anemocóricas, devido à grande quantidade de espécies zoocóricas de sub-bosque e dossel. O ar seco e os fortes ventos característicos em Corumbá, MS durante o período de estiagem poderiam facilitar o processo de secagem e maturação das vagens de espécies anemocóricas e autocóricas, como em Fabaceae (Leguminosae), Bignoniaceae e Malvaceae. (Janzen 1967). Além disso, a dispersão dos diásporos é facilitada pela deciduidade da vegetação durante o período mais seco do ano, contribuindo para o sucesso da dispersão das sementes anemocóricas longe da planta-mãe (Janzen 1967). Já a ocorrência de espécies zoocóricas nos estratos inferiores das duas florestas deciduais amostradas parece estar ligada à área de atuação dos animais dispersores, como observado por Frankie et al. (1974) e Morellato & Leitão-Filho (1992).
Algumas espécies típicas das formações estacionais deciduais, como Myracrodruon urundeuva e Anadenanthera colubrina, que são dominantes nessas formações, também estão presentes como espécies mais abundantes em vegetações semideciduais no Município de Corumbá (G.A. Damasceno-Júnior, dados não publicados). Estas espécies, juntamente com Commiphora leptophloeos, Amburana cearensis, Cnidoscolus cnicodendron e Coutarea hexandra possuem ampla distribuição no domínio da Caatinga (Lorenzi 2002a, b, Pott & Pott 1994). Das 67 espécies decíduas identificadas por Salis et al. (2004) em mata decídua ao redor de Corumbá, a maioria também se distribui em outras formações sazonais como o Chaco e Caatinga. Estes resultados concordam com o proposto por Prado & Gibbs (1993), que mencionam as formações residuais de mata seca de Corumbá como integrantes do Arco Pleistocênico, uma provável rota migratória de espécies remanescentes de mudanças climáticas ocorridas no Pleitoceno Superior, abrangendo a Caatinga e matas calcárias do sudeste brasileiro, até alcançar regiões de matas secas na Bolívia, Argentina e Peru. A coincidência encontrada na presença de alguns gêneros e famílias entre as diferentes florestas decíduas espalhadas na América Latina é explicada por Bigarella et al. (1975) como resultados de fenômenos ocorridos no Quaternário que propiciaram uma distribuição contínua dos taxa, sendo mais tarde isolados por disjunções decorrentes de mudanças climáticas desfavoráveis, com conseqüente diferenciação genética e especiação. Estes remanescentes são constituídos de elementos do Chaco, Caatinga e Cerrado, representando fonte de diversidade biológica para a região do Pantanal e certamente desempenham papel importante como parte dos corredores naturais de florestas da região.
Agradecimentos - Os autores agradecem ao Prof. Otávio Froehlich (UFMS) pela revisão dos textos em inglês, ao pesquisador da Embrapa-CPAP Luiz Alberto Pellegrin, do laboratório de geoprocessamento, pela estimativa das áreas dos remanescentes e disponibilização da imagem em satélite, e à Propp/UFMS pelo auxílio financeiro concedido.
(recebido: 06 de janeiro de 2010; aceito: 10 de junho de 2010)
- APG II. 2003. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG II. Botanical Journal of the Linnean Society 141:399-436.
- BIGARELLA, J.J., ANDRADE LIMA, D. & RIEHS, P.J. 1975. Considerações a respeito das mudanças paleoambientais na distribuição de algumas espécies vegetais e animais no Brasil. Annais da Academia Brasileira de Ciências 47:411-464.
- BROWER, J.E. & ZAR, J.H. 1984. Field and laboratory methods for general ecology. 2nd ed., C. Brown, Iowa.
- CECCON, E., HUANTE, P. & RINCÓN, E. 2006. Abiotic factors influencing tropical dry forests regeneration. Brazilian Archives of Biology and Technology 49:305-312.
- CESTARO, L.A. & SOARES, J.J. 2004. Variações florística e estrutural e relações fitogeográficas de um fragmento de floresta decídua no Rio Grande do Norte, Brasil. Acta Botanica Brasilica 18:203-218.
- DAMASCENO-JÚNIOR, G.A. & BEZERRA, M.A.O. 2004. Estudo fitossociológico em uma ilha fluvial na Lagoa do Castelo, Pantanal, Corumbá, MS. In Anais do IV Simpósio sobre recursos naturais e sócio-econômicos do Pantanal: sustentabilidade regional (B.M.A. Soriano, S.M. Salis, G. Mourão & L.A. Pelegrin, eds.). Embrapa Pantanal, Corumbá
- FRANKIE, G.W., BAKER, H.G. & OPLER, P.A. 1974. Comparative phenological studies of trees in tropical wet and dry forests in the lowlands of Costa Rica. Journal of Ecology 62:881-919.
- GENTRY, A.H. 1995. Diversity and floristic composition of neotropical dry forests. In Seasonally dry tropical forests (S.H. Bullock, A. Mooney & E. Medina, eds.). Cambridge University Press, Cambridge, p.146-194.
- IBGE. 1992. Manual técnico da vegetação brasileira. Fundação Instituto Brasileiro de Geografia e Estatística, Departamento de Recursos Naturais e Estudos Ambientais, Rio de Janeiro.
- IVANAUSKAS, N.M. & RODRIGUES, R.R. 2000. Florística e fitossociologia de remanescentes de floresta estacional decidual em Piracicaba, São Paulo, Brasil. Revista Brasileira de Botânica 23:291-304.
- JANZEN, D.H. 1967. Synchronization of sexual reproduction of trees within the dry season in Central America. Evolution 21:620-637.
- KILLEEN, T.J., JARDIM, A., MAMANI, F. & ROJAS, N. 1998. Diversity, composition and structure of a tropical semideciduous forest in the Chiquitanía region of Santa Cruz, Bolívia. Journal of Tropical Ecology 14:803-827.
- KÖPPEN, W.P. 1948. Climatología: con un estudio de los climas de la Tierra. Fondo de Cultura Económica, México.
- LINARES-PALOMINO, R., PENNINGTON, R.T. & BRIDGEWATER, S. 2003. The phytogeography of the seasonally dry tropical forests in Equatorial Pacific South America. Candollea 58:473-499.
- LORENZI, H. 2002a. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 4Ş ed., Plantarum, Nova Odessa, v.1.
- LORENZI, H. 2002b. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 2Ş ed., Plantarum, Nova Odessa, v.2.
- MARTINS, F.R. 1993. Estrutura de uma floresta mesófila. Série Teses, Editora da Universidade Estadual de Campinas, Campinas.
- MCCUNE, B. & MEFFORD, M.J. 1997. Multivariate analysis of ecological data: version 3.17. MjM Software, Gleneden Beach.
- MOONEY, H.A., BULLOCK, S.H. & MEDINA, E. 1995. Introduction. In Seasonally dry tropical forests (S.H. Bullock, A. Mooney & E. Medina, eds.). Cambridge University Press, Cambridge, p.1-8.
- MORELLATO, L.P.C. & LEITÃO FILHO, H.F. 1992. Padrões de frutificação e dispersão na Serra do Japi. In História natural da Serra do Japi: ecologia e preservação de uma área florestal no sudeste do Brasil (L.P.C. Morellato, org.). Editora da Universidade Estadual de Campinas, Campinas, p.112-141.
- OKIDA, R. & ANJOS, C.E. 2000. Geomorfologia. In Zoneamento ambiental da borda oeste do Pantanal: Maciço do Urucum e adjacências. (J.S.V. Silva, ed.). Embrapa, Brasília, p.47-54.
- PENNINGTON, R.T., PRADO, D.E. & PENDRY, C.A. 2000. Neotropical seasonally dry forests and Quaternary vegetation changes. Journal of Biogeography 27:261-273.
- PIELOU, E.C. 1975. Ecological diversity. John Wiley & Sons, New York.
- POTT, A. & POTT, V.J. 1994. Plantas do Pantanal. Embrapa Pantanal, Brasília.
- PRADO, D.E. & GIBBS, P. 1993. Patterns of species distributions in the dry seasonal forest of South America. Annals of Missouri Botanic Garden 80:902-927.
- PRANCE, G. & SCHALLER, G.B. 1982. Preliminary study of some vegetation types of the Pantanal, Mato Grosso, Brazil. Brittonia 34:228-251.
- RATTER, J.A., POTT, A., POTT, V.J., CUNHA, C.N. & HARIDASAN, M. 1988. Observation on wood vegetation types in the Pantanal at Corumbá, Brazil. Notes of the Royal Botanic Garden of Edinburgh 45:503-525.
- SALIS, S.M., SILVA, M.P., MATTOS, P.P., SILVA, J.S.V., POTT, V.J. & POTT, A. 2004. Fitossociologia de remanescentes de floresta estacional decidual em Corumbá, Estado do Mato Grosso do Sul, Brasil. Revista Brasileira de Botânica 27:671-684.
- SCARIOT, A. & SEVILHA, A.C. 2000. Diversidade, estrutura e manejo de florestas deciduais e as estratégias para a conservação. In Tópicos atuais em botânica (T.B. Cavalcanti & B.M.T. Walter, orgs.). Sociedade Botânica do Brasil/Embrapa Recursos Genéticos e Biotecnologia, Brasília, p.183-188.
- SCHILLING, A.C. & BATISTA, J.L.F. 2008. Curva de acumulação de espécies e suficiência amostral em florestas tropicais. Revista Brasileira de Botânica 31:179-187.
- SHEPHERD, G.J. 1994. Fitopac 1 - manual do usuário. Universidade Estadual de Campinas, Campinas.
- SILVA, L.A. & SCARIOT, A. 2003. Composição florística e estrutura da comunidade arbórea em uma floresta estacional decidual em afloramento calcário (Fazenda São José, São Domingos, GO, Bacia do Rio Paraná). Acta Botanica Brasilica 17:305-313.
- SILVA, L.A. & SCARIOT, A. 2004. Comunidade arbórea de uma floresta estacional decídua sobre afloramento calcário na bacia do Rio Paraná. Revista Árvore 28:61-67.
- SORIANO, B.M.A. 1997. Caracterização climática de Corumbá, MS. Boletim de Pesquisa 11. Embrapa-CPAP, Corumbá
- SPERA, S.T., TOSTO, S.G., CARDOSO, E.L. & OLIVEIRA, H. 1997. Levantamento de reconhecimento de alta intensidade dos solos e avaliação da aptidão agrícola das terras da borda oeste do Pantanal: Maciço do Urucum e adjacências, MS. Boletim de Pesquisa 9, Embrapa CPAP/Embrapa CNPS, Corumbá/Rio de Janeiro.
- SPIEGEL, M.R. 1977. Estatística. McGraw Hill do Brasil, São Paulo.
- VAN DER PIJL, L. 1972. Principles of dispersal in higher plants. 2nd ed. Springer, Berlin.
- VÁSQUEZ, G.J.A. & GIVNISH, T.J. 1998. Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantlán. Journal of Ecology 86:999-1020.
- WEBB, E.L., STANFIELD, B.J. & JENSEN, M.L. 1999. Effects of topography on rainforest tree community structure and diversity in American Samoa, and implications for frugivore and nectarivore populations. Journal of Biogeography 26:887-897.
- WERNECK, M.S., FRANCESCHINELLI, E.V. & TAMEIRÃO NETO, E. 2000. Mudanças na florística e estrutura de uma floresta decídua durante um período de quatro anos (1994-1998), na região do Triângulo Mineiro, MG. Revista Brasileira de Botânica 23:401-413.
- WHITE, D.A. & HOOD, C.S. 2004. Vegetation patterns and environmental gradients in tropical dry forest of the northern Yucatan Peninsula. Journal of Vegetation Science 15:151-160.
Datas de Publicação
-
Publicação nesta coleção
05 Out 2010 -
Data do Fascículo
Set 2010
Histórico
-
Recebido
06 Jan 2010 -
Aceito
10 Jun 2010