Abstract
2-Hydroxybutyric acid appears at high concentrations in situations related to deficient energy metabolism (e.g., birth asphyxia) and also in inherited metabolic diseases affecting the central nervous system during neonatal development, such as "cerebral" lactic acidosis, glutaric aciduria type II, dihydrolipoyl dehydrogenase (E3) deficiency, and propionic acidemia. The present study was carried out to determine the effect of 2-hydroxybutyric acid at various concentrations (1-10 mM) on CO2 production and lipid synthesis from labeled substrates in cerebral cortex of 30-day-old Wistar rats in vitro. CO2 production was significantly inhibited (30-70%) by 2-hydroxybutyric acid in cerebral cortex prisms, in total homogenates and in the mitochondrial fraction. We also demonstrated a significant inhibition of lipid synthesis (20-45%) in cerebral cortex prisms and total homogenates in the presence of 2-hydroxybutyric acid. However, no inhibition of lipid synthesis occurred in homogenates free of nuclei and mitochondria. The results indicate an impairment of mitochondrial energy metabolism caused by 2-hydroxybutyric acid, a fact that may secondarily lead to reduction of lipid synthesis. It is possible that these findings may be associated with the neuropathophysiology of the situations where 2-hydroxybutyric acid is accumulated.
lipid synthesis; energy metabolism; cerebral cortex; 2-hydroxybutyric acid
Braz J Med Biol Res, May 2001, Volume 34(5) 627-631 (Short Communication)
Inhibition of in vitro CO 2 production and lipid synthesis by 2-hydroxybutyric acid in rat brain
A.R. Silva, C. Ruschel, C. Helegda, A.T.S. Wyse, C.M.D. Wannmacher, M. Wajner and C.S. Dutra-Filho
Departamento de Bioquímica, Instituto de Ciências Básicas da Saúde, Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brasil
Text
Abstract

2-Hydroxybutyric acid appears at high concentrations in situations related to deficient energy metabolism (e.g., birth asphyxia) and also in inherited metabolic diseases affecting the central nervous system during neonatal development, such as "cerebral" lactic acidosis, glutaric aciduria type II, dihydrolipoyl dehydrogenase (E3) deficiency, and propionic acidemia. The present study was carried out to determine the effect of 2-hydroxybutyric acid at various concentrations (1-10 mM) on CO2 production and lipid synthesis from labeled substrates in cerebral cortex of 30-day-old Wistar rats in vitro. CO2 production was significantly inhibited (30-70%) by 2-hydroxybutyric acid in cerebral cortex prisms, in total homogenates and in the mitochondrial fraction. We also demonstrated a significant inhibition of lipid synthesis (20-45%) in cerebral cortex prisms and total homogenates in the presence of 2-hydroxybutyric acid. However, no inhibition of lipid synthesis occurred in homogenates free of nuclei and mitochondria. The results indicate an impairment of mitochondrial energy metabolism caused by 2-hydroxybutyric acid, a fact that may secondarily lead to reduction of lipid synthesis. It is possible that these findings may be associated with the neuropathophysiology of the situations where 2-hydroxybutyric acid is accumulated.
Key words: lipid synthesis, energy metabolism, cerebral cortex, 2-hydroxybutyric acid
2-Hydroxybutyric acid originates from 2-ketobutyrate which is a product of the catabolism of the amino acids 2-aminobutyrate, methionine, threonine, and homoserine (1,2). The conversion of 2-ketobutyrate to 2-hydroxybutyric acid occurs when the NADH/NAD+ ratio is increased in the cytosol (2), as observed in renal tissue biopsies after ischemia provoked by clamping the renal artery (3). High excretion of 2-hydroxybutyric acid also occurs in neonates with severe birth asphyxia causing encephalopathy (4), and in some "cerebral" organic acidemias such as glutaric aciduria type II (5) and dihydrolipoyl dehydrogenase (E3) deficiency which produces a combined deficiency of pyruvate, a-ketoglutarate and branched-chain a-keto acid dehydrogenases (6). Increased concentrations of 2-hydroxybutyric acid were also observed in cerebrospinal fluid (CSF) of patients presenting "cerebral" lactic acidosis (which may be due to biotinidase deficiency or to some mitochondriopathies), and propionic acidemia (7). These inherited metabolic disorders are clinically characterized by severe neurological manifestations such as mental retardation, convulsions, neuropsychomotor delay, ataxia and alteration of muscle tonus and by significant brain atrophy (8). Normal levels of 2-hydroxybutyric acid range from 11 to 86 µmol/l in CSF and from 8 to 80 µmol/l in plasma, whereas in these organic acidemias CSF levels may achieve 1.7 mM (7,9), reflecting its accumulation in the central nervous system (CNS). We recently reported preliminary results showing an inhibition of CO2 production by 2-hydroxybutyric acid (10).
Therefore, since 2-hydroxybutyric acid is present at high concentrations in the brain of patients with various neurodegenerative disorders (4-7,8) and lipid synthesis is crucial to the normal development and function of the CNS (e.g., myelination and synaptogenesis) (11), in the present study we investigated the influence of 2-hydroxybutyric acid on lipid synthesis in cerebral cortex of young rats. The effect of the acid on brain CO2 production was also determined.
All chemicals were purchased from Sigma Chemical Co., St. Louis, MO, USA, except for the radiolabeled compounds ([U-14C]-acetate and [1,5-14C]-citrate) which were purchased from Amersham International plc, Little Chalfont, UK. We used in the experiments a commercially available DL isomer mixture of 2-hydroxybutyric acid. Thirty-day-old Wistar rats bred in this laboratory were used. Rats had free access to a 20% protein commercial chow and water and were kept in a room on a 12-h light/dark cycle at a temperature of 24 ± 1oC. Animals were killed by decapitation without anesthesia, brains were removed immediately and the cortex was dissected onto an ice-cold glass plate.
Brain cortex was cut in two perpendicular directions to produce 400-µm wide prisms using a McIlwain chopper. Prisms were pooled and weighed.
Cerebral cortex was homogenized in (1:10, w/v) 0.32 M sucrose, 1 mM EDTA-K+, and 10 mM Tris-HCl buffer, pH 7.4.
The mitochondrial fraction and the homogenate free of nuclei and mitochondria (HFNM) were prepared as follows. The homogenate was centrifuged at 3,000 g for 10 min to remove unbroken cells and nuclei. The supernatant was centrifuged at 10,000 g for 20 min and the pellet from the second centrifugation containing the mitochondrial fraction was suspended in 2 ml of the above buffer. The supernatant from this centrifugation was used as HFNM.
The cerebral cortex prisms (50 mg) were added to small flasks (11 cm3) containing 0.5 ml Krebs-Ringer bicarbonate buffer, pH 7.4. The homogenates (300 µl), mitochondrial fractions (300 µl) or HFNM (300 µl) were added to 0.5 ml of an intracellular medium, pH 7.4, containing 4.0 mM KCl, 0.21 mM EDTA, 2.0 mM KH2PO4, 3.5 mM MgSO4, 59 mM NaCl, 70 mM Tris-HCl, pH 7.4, 1.2 mM ATP, and 0.16 mM NAD+ according to the method of McKenna et al. (12). Flasks were pre-incubated in a metabolic shaker at 37oC for 15 min (90 oscillations/min). After pre-incubation, 0.2 µCi [U-14C]-acetate or [1,5-14C]-citrate and the respective unlabeled substrates were added to the incubation medium at a final concentration of 0.5 mM. DL-2-hydroxybutyric acid (buffered to pH 7.4) was added to the incubation medium at a final concentration of 1.0 mM. The controls did not contain the acid. The flasks were gassed with a O2:CO2 (95:5) mixture and sealed with rubber stoppers and Parafilm M. Glass center wells containing a folded 65 mm/5 mm piece of Whatman No. 3 filter paper were hung from the stoppers. After 60 or 120 min of incubation at 37oC, 0.25 ml of 50% trichloroacetic acid was added to the medium and 0.1 ml of benzethonium hydroxide was added to the center wells with needles introduced through the rubber stopper. The flasks were left to stand for 30 min to complete CO2 trapping and then opened. The filter papers were removed and added to vials containing scintillation fluid, and radioactivity was measured (13). CO2 production was calculated as µmol produced per hour per mg protein.
Lipid synthesis from acetate was performed in the same incubation medium as used to measure CO2 production. After the flasks were opened, brain tissue was collected and homogenized and the lipids were extracted with chloroform-methanol (2:1) by centrifugation for 10 min at 5,000 g. Chloroform-methanol was evaporated overnight, scintillation fluid was added and radioactivity was measured (13). Lipid synthesis was calculated as µmol of total lipids produced from acetate per hour per mg protein.
Protein was measured by the method of Lowry et al. (14) using bovine serum albumin as standard. Data were analyzed by the paired Student t-test since our in vitro experimental design used samples (brain fractions) from the same animal in the control and in the test groups. One-way ANOVA followed by the Duncan multiple range test was also performed to test the effect of various concentrations of the acid.
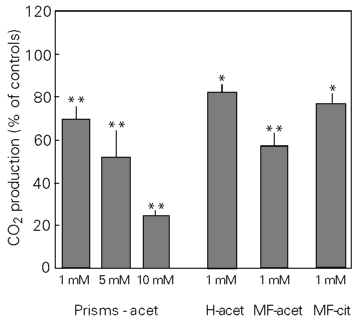
Figure 1 shows the in vitro CO2 production from 14C-acetate in prisms or homogenates and from 14C-acetate and 14C-citrate in the mitochondrial fraction of cerebral cortex in the presence of DL-2-hydroxybutyric acid. The results are expressed as mean ± SEM and correspond to percentage of controls. The control values were 129 ± 11.2 nmol h-1 g tissue-1. DL-2-hydroxybutyric acid inhibited CO2 production from acetate in prisms (F = 20.43; P<0.01) in a dose-dependent manner with maximal inhibition (70%) at 10 mM. The same effect occurred using homogenates (t = 4.70; P<0.05) and the mitochondrial fraction (t = 7.28; P<0.01). CO2 production in prisms was also inhibited by 10 mM DL-2-hydroxybutyric acid when 2-h incubation was performed (t = 17.88; P<0.01). In addition, CO2 production from citrate was also inhibited by 1 mM DL-2-hydroxybutyric acid in the mitochondrial fraction (t = 4.37; P<0.05).
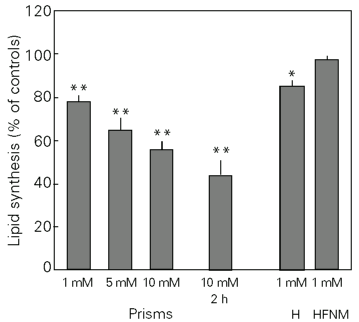
Figure 2 shows the effect of DL-2-hydroxybutyric acid on in vitro lipid synthesis from 14C-acetate by prisms, homogenates and HFNM of cerebral cortex. The results are expressed as mean ± SEM and correspond to percentage of controls. The control values were 164 ± 15.9 nmol h-1 g tissue-1. It can be seen that the acid significantly inhibited lipid synthesis (F = 24.02; P<0.01) in prisms in a dose-dependent manner with maximal inhibition (45%) at 10 mM. The same effect, but to a larger extent (60%), was verified when incubation with 10 mM DL-2-hydroxybutyric acid was carried out for 2 h (t = 14.53; P<0.01). We also observed inhibition of lipid synthesis in homogenates (t = 5.56; P<0.01), but not in HFNM (t = 0.80; P>0.05).
Although brain accumulation and excretion of 2-hydroxybutyric acid occurs in encephalopathy associated with hypoxia and ischemia and in some cerebral organic acidemias (4-7,9) in which the CSF concentrations of this metabolite are approximately 20- to 50-fold higher than in normal individuals, the effect of 2-hydroxybutyric acid on brain metabolism is virtually unknown.
We have recently presented preliminary results showing that 10 mM 2-hydroxybutyric acid inhibits CO2 formation in the brain (10). In the present study, we confirmed and extended these findings by demonstrating a dose-dependent effect of the acid on CO2 production from acetate and also from citrate, starting at concentrations as low as 1 mM, similar to those found in CSF of patients affected by various organic acidemias. Moreover, CO2 production was also strongly diminished by DL-2-hydroxybutyric acid in all brain preparations studied, but more prominent when mitochondrial preparations were used, a fact that suggests an intramitochondrial mechanism of action of 2-hydroxybutyric acid. In agreement with our results, Hammer and co-workers (15) reported that 14CO2 production from threonine is inhibited in isolated hepatocytes from domestic cats in the presence of 10 mM 2-hydroxybutyric acid in the incubation medium.
We also observed that DL-2-hydroxybutyric acid provokes a significant and dose-dependent inhibition of in vitro lipid synthesis in prisms and homogenates of cerebral cortex from young rats. However, when mitochondria were not present in the incubation medium (HFNM), no inhibition was detected, suggesting that mitochondria are critical for the inhibitory effect of DL-2-hydroxybutyric acid and/or that this inhibition cannot be possibly attributed to a direct action of the acid on the enzymes of the lipid synthesis pathway. Since energy is necessary to lipid synthesis, it is tempting to speculate that the impairment of lipid synthesis detected in our in vitro assays may be a consequence of the inhibitory effect of DL-2-hydroxybutyric acid on energy production (decreased CO2 production).
Some mechanisms can be discussed to explain our findings. A competition of 2-hydroxybutyric acid with acetate for the same monocarboxylic membrane transporter could occur (16,17), reducing the availability of the labeled substrate (acetate) to produce 14CO2 and lipids. However, this is probably not the case since the inhibition of CO2 production and lipid synthesis persisted when lysed membrane preparations (homogenates) were used. In addition, when labeled citrate, which crosses mitochondria through the tricarboxylic acid membrane carrier, was employed, CO2 production was still inhibited by DL-2-hydroxybutyric acid. An alternative mechanism could be the inhibition of acetyl CoA synthetase (EC 6.2.1.1), the enzyme that produces acetyl CoA from acetate. Again, this is unlikely since this enzyme is present in both mitochondria and cytosol (18), and 2-hydroxybutyric acid was not able to inhibit lipid synthesis when an HFNM preparation is used. 2-Hydroxybutyric acid could also sequester coenzyme A, leaving a lower amount of CoA for acetate and citrate binding. However, this would not explain the differential effect of the acid on lipid synthesis in distinct brain preparations.
It is difficult to extrapolate our findings to the human situations in which increased levels of 2-hydroxybutyric acid are found, but if the human condition is similar, reduction of brain energy metabolism and lipid synthesis may be associated with the pathophysiology of the neurological dysfunction found in these disorders.
Address for correspondence: C.S. Dutra Filho, Departamento de Bioquímica, ICBS, UFRGS, Rua Ramiro Barcelos, 2600, anexo 90035-003 Porto Alegre, RS, Brasil. Fax: +55-51-316-5535. E-mail: dutra@ufrgs.br
Research supported by PRONEX/FINEP, CNPq and FAPERGS. Received April 12, 2000. Accepted January 30, 2001.
- 1. Bremer J (1969). Pyruvate dehydrogenase, substrate specificity and product inhibition. European Journal of Biochemistry, 8: 535-540.
- 2. Landaas S (1975). The formation of 2-hydroxybutyric acid in experimental animals. Clinica Chimica Acta, 58: 23-32.
- 3. Niwa T, Yamamoto N, Asada H, Kawanishi A, Yokoyama M, Maeda K & Ohki T (1983). Ischemic change of organic acids in kidney. Journal of Chromatography, 272: 227-232.
- 4. Walker V & Mills GA (1992). Effects of birth asphyxia on urinary organic acid excretion. Biology of the Neonate, 61: 162-172.
- 5. Przyrembel H, Wendel U, Becker K, Bremer HJ, Bruinvis L, Ketting D & Wadman SK (1976). Glutaric aciduria type II: report on a previously undescribed metabolic disorder. Clinica Chimica Acta, 66: 227-239.
- 6. Kuhara T, Shinka T, Inoue Y, Matsumoto M, Yoshino M, Sakaguchi Y & Matsumoto I (1983). Studies of urinary organic acid profiles of a patient with dihydrolipoyl dehydrogenase deficiency. Clinica Chimica Acta, 133: 133-140.
- 7. Hoffmann GF, Meier-Augenstein W, Stöckler S, Surtees R, Rating D & Nyhan WL (1993). Physiology and pathophysiology of organic acids in cerebrospinal fluid. Journal of Inherited Metabolic Disease, 16: 648-669.
- 8. Brandt NJ (1984). Symptoms and signs in organic acidurias. Journal of Inherited Metabolic Disease, 7 (Suppl 1): 23-27.
- 9. Hoffmann GF, Gibson KM, Trefz FK, Nyhan WL, Bremer HJ & Rating D (1994). Neurological manifestations of organic acid disorders. European Journal of Pediatrics, 153 (Suppl 1): S94-S100.
- 10. Dutra-Filho CS, Wajner M, Wannmacher CMD, Gassen E, Candiago RH, Wilhems A & Malfussi HFC (1995). 2-Hydroxybutyrate and 4-hydroxybutyrate inhibit CO2 formation from labeled substrates by rat cerebral cortex. Biochemical Society Transactions, 23 (Suppl): 228S.
- 11. Edmond J, Higa TA, Korsak RA, Bergner EA & Lee WN (1998). Fatty acid transport and utilization for the developing brain. Journal of Neurochemistry, 70: 1227-1234.
- 12. McKenna MC, Bezold LI, Kimatian SJ & Tildon JT (1986). Competition of glycerol with other oxidizable substrates in rat brain. Biochemical Journal, 237: 47-51.
- 13. Silva AR, Ruschel C, Helegda C, Brusque AM, Wannmacher CMD, Wajner M & Dutra-Filho CS (1999). Inhibition of rat brain lipid synthesis in vitro by 4-hydroxybutyric acid. Metabolic Brain Disease, 14: 157-164.
- 14. Lowry OH, Rosebrough NJ, Farr AL & Randall RJ (1951). Protein measurement with the Folin phenol reagent. Journal of Biological Chemistry, 193: 265-275.
- 15. Hammer VA, Rogers QR & Freedland RA (1996). Threonine is catabolized by L-threonine 3-dehydrogenase and threonine dehydratase in hepatocytes from domestic cats (Felis domestica). Journal of Nutrition, 136: 2218-2226.
- 16. Oldendorf WH (1973). Carrier-mediated blood-brain barrier transport of short-chain monocarboxylic organic acids. American Journal of Physiology, 224: 1450-1453.
- 17. Jackson VN & Halestrap AP (1996). The kinetics, substrate, and inhibitor specificity of the monocarboxylate (lactate) transporter of rat liver cells determined using the fluorescent intracellular pH indicator, 2'-7'-bis (carboxyethyl)-carboxyfluorescein. Journal of Biological Chemistry, 271: 861-868.
- 18. Yip V, Carter JG, Pusateri ME, McDougal Jr DB & Lowry OH (1991). Distribution in brain and retina of four enzymes of acetyl CoA synthesis in relation to choline acetyl transferase and acetylcholine esterase. Neurochemical Research, 16: 629-635.
Correspondence and Footnotes
Publication Dates
-
Publication in this collection
19 Apr 2001 -
Date of issue
May 2001
History
-
Accepted
30 Jan 2001 -
Received
12 Apr 2000