Abstract
The duration of the intraerythrocytic cycle of Plasmodium is a key factor in the pathogenicity of this parasite. The simultaneous attack of the host red blood cells by the parasites depends on the synchronicity of their development. Unraveling the signals at the basis of this synchronicity represents a challenging biological question and may be very important to develop alternative strategies for therapeutic approaches. Recently, we reported that the synchrony of Plasmodium is modulated by melatonin, a host hormone that is synthesized only during the dark phases. Here we report that N-acetyl-serotonin, a melatonin precursor, also releases Ca2+ from isolated P. chabaudi parasites at micro- and nanomolar concentrations and that the release is blocked by 250 mM luzindole, an antagonist of melatonin receptors, and 20 mM U73122, a phospholipase C inhibitor. On the basis of confocal microscopy, we also report the ability of 0.1 µM melatonin and 0.1 µM N-acetyl-serotonin to cross the red blood cell membrane and to mobilize intracellular calcium in parasites previously loaded with the fluorescent calcium indicator Fluo-3 AM. The present data represent a step forward into the understanding of the signal transduction process in the host-parasite relationship by supporting the idea that the host hormone melatonin and N-acetyl-serotonin generate IP3 and therefore mobilize intracellular Ca2+ in Plasmodium inside red blood cells.
Malaria; Plasmodium; Calcium; Melatonin; Circadian rhythm; Host-parasite interaction
Braz J Med Biol Res, November 2003, Volume 36(11) 1583-1587
Melatonin and N-acetyl-serotonin cross the red blood cell membrane and evoke calcium mobilization in malarial parasites
C.T. Hotta, R.P. Markus and C.R.S. Garcia
Departamento de Fisiologia, Instituto de Biociências, Universidade de São Paulo, São Paulo, SP, Brasil
References
Abstract
The duration of the intraerythrocytic cycle of Plasmodium is a key factor in the pathogenicity of this parasite. The simultaneous attack of the host red blood cells by the parasites depends on the synchronicity of their development. Unraveling the signals at the basis of this synchronicity represents a challenging biological question and may be very important to develop alternative strategies for therapeutic approaches. Recently, we reported that the synchrony of Plasmodium is modulated by melatonin, a host hormone that is synthesized only during the dark phases. Here we report that N-acetyl-serotonin, a melatonin precursor, also releases Ca2+ from isolated P. chabaudi parasites at micro- and nanomolar concentrations and that the release is blocked by 250 mM luzindole, an antagonist of melatonin receptors, and 20 mM U73122, a phospholipase C inhibitor. On the basis of confocal microscopy, we also report the ability of 0.1 µM melatonin and 0.1 µM N-acetyl-serotonin to cross the red blood cell membrane and to mobilize intracellular calcium in parasites previously loaded with the fluorescent calcium indicator Fluo-3 AM. The present data represent a step forward into the understanding of the signal transduction process in the host-parasite relationship by supporting the idea that the host hormone melatonin and N-acetyl-serotonin generate IP3 and therefore mobilize intracellular Ca2+ in Plasmodium inside red blood cells.
Key words: Malaria, Plasmodium, Calcium, Melatonin, Circadian rhythm, Host-parasite interaction
Introduction
One of the puzzling questions concerning the development of malaria parasites is the search for the molecules which might be involved in signaling processes in a parasite that is not exposed to the extracellular milieu and therefore to body fluids. Processes such as cell cycle progression or arrest, cell mobility, and stage-specific gene expression are all required for parasite development in the intraerythrocytic stage (1). The parasite maturation process comprises an intraerythrocytic cycle where defined stages known as ring, trophozoite and schizont take place. The beginning of a new stage as well as cell division and synchronicity may require protein synthesis and phosphorylation which are signal-mediated processes.
Recent studies on the dynamics of calcium in malaria parasites also indicate similarities with mammalian cells, such as the existence of a thapsigargin-sensitive pool in the human malaria parasite Plasmodium falciparum (2) and in the rodent parasite P. chabaudi (3-6). In addition to the endoplasmic reticulum Ca2+ pool, the existence of acidic calcium pools in both parasites indicates that they possess multiple mechanisms to store calcium during their intraerythrocytic developmental stage (6). The putative signal which might be involved in emptying these Ca2+ pools in P. chabaudi has been reported to be the second messenger IP3, thus suggesting that these parasites utilize a calcium-mediated cell signaling. In this regard, we have reported that the parasite is surrounded by a high calcium environment within the red blood cell (7).
We have recently reported that the host hormone melatonin modulates circadian rhythms of malarial parasites by inducing calcium release from their intracellular stores. Melatonin was shown to be a key factor not only for the synchronicity of the maturation process of P. falciparum but also for the development of infection with P. chabaudi (8).
Melatonin is the hormone synthesized during the dark phase by the pineal gland and is responsible for the transduction of the environmental illumination of several physiological events that change diurnally in vertebrates (9), arthropods (10), higher plants (11), and dinoflagellates (12). The dependence of malaria infection on daylight is clear from the fact that changes in the host photoperiod also change the timing of the parasite cell cycle (13).
In P. chabaudi the rupture of infected blood cells and therefore the reinvasion of new red blood cells occur between midnight and 3:00 am (14,15) which coincides with the peak of melatonin levels in vertebrates. When mice are submitted to an inverted rhythm, the parasite cycle is also inverted (16) and schizogony occurs around midday (15). Interestingly, the pineal gland is thought to play a role in mediating photoperiodic control of growth and division synchrony in malaria parasites since pinealectomy in mice abolishes division and synchrony in the rodent malaria parasite P. berghei (13).
Melatonin and N-acetyl-serotonin (NAS) are thought to induce IP3 generation in different systems such as hamster melanoma cells (17), pigeon brain (18), chick brain (19), and dinoflagellates (20). The ability of IP3 to mobilize Ca2+ from intracellular stores and therefore to trigger many biological processes such as cell division is well known (21). Therefore, Ca2+ release by NAS receptor stimulation might play a role in parasite division.
Material and Methods
Parasite maintenance
P.chabaudi (clone F IP-Pc1) was maintained in female mice (BALB/c) by transfer of infection. The procedure for blood collection from mice and platelet removal has been previously described (6).
Intracellular calcium measurements
Melatonin- and NAS-induced Ca2+ release in P. chabaudi was determined in intact parasites. For these measurements, isolated parasites obtained from infected red blood cells after saponin lysis were washed twice in buffer A (116 mM NaCl, 5.4 mM KCl, 0.8 mM MgSO4, 5.5 mM D-glucose, and 50 mM Mops, pH 7.2) and resuspended at 108 parasites/ml in the same buffer containing probenecid, an inhibitor of organic anion transport. The procedure for labeling isolated parasites with Fluo-3 was previously described (8). A similar procedure was performed to load red blood cells infected with P. chabaudi.
All experiments were performed in the presence of protease inhibitors, leupeptin, pepstatin A, antipain and chymostatin at 20 µg/ml and benzamidine at 0.5 mM. Measurements were made at 37ºC with a Hitachi F-4500 spectrofluorimeter (Tokyo, Japan) with emission at 530 nm and excitation at 505 nm and a Zeiss LSM-510 confocal microscope (Oberkochen, Germany) using 488 nm excitation and 550 nm emission wavelength filters. Samples were observed on microscopy plates (MatTek Corp., Ashland, MA, USA) previously incubated with polylysine for 1 h and washed.
Results and Discussion
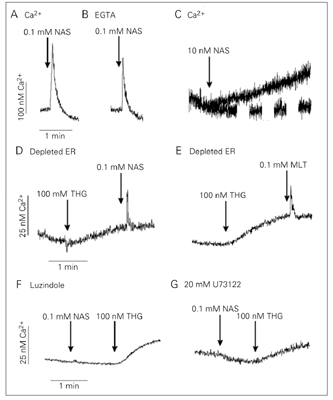
In the present study, we show that NAS elicits Ca2+ release from P. chabaudi parasites at the trophozoite stage in a Ca2+ medium (Figure 1). The amount of Ca2+ released by 0.1 mM NAS was 130 ± 5 nM (N = 3; Figure 1A) while melatonin elicited 80 ± 5 nM (N = 3) (8). The agonists still mobilized Ca2+ in the absence of extracellular Ca2+ (plus 2 mM EGTA), indicating a mobilization from intracellular stores (Figure 1B). A physiological concentration of NAS (10 nM) caused, after a brief lag phase, a slow increase in [Ca2+]i that reached a plateau about 60 ± 10 nM above the prestimulation level (Figure 1C). It is worth noting that the kinetics of the rise of [Ca2+]i with supramaximal doses was different from the rise induced by physiological concentrations.
The depletion of the endoplasmic reticulum Ca2+ pool by thapsigargin, a sarcoendoplasmic Ca2+ ATPase inhibitor, reduced the Ca2+ release induced by the addition of 0.1 mM NAS (Figure 1D) and 0.1 mM melatonin (Figure 1E), suggesting that both hormones induce Ca2+ release from a thapsigargin-sensitive pool.
Luzindole, an antagonist of melatonin receptors, blocked the NAS-induced Ca2+ release (Figure 1F). In addition, this Ca2+ release could also be blocked by the phospholipase C inhibitor, U73122 (Figure 1G). These data indicate that, in addition to melatonin (8), its precursor, NAS also stimulates the phospholipase C pathway which leads to Ca2+ release from the endoplasmic reticulum through a melatonin-binding site that can be blocked by luzindole.
To determine if melatonin and NAS are able to release calcium under physiological conditions, we labeled P. chabaudi-infected red blood cells with the fluorescent calcium indicator Fluo-3 AM and performed experiments similar to those described above. Indeed both 0.1 µM NAS and 0.1 µM melatonin induced a slow increase in Fluo-3 fluorescence, indicating the occurrence of intracellular calcium mobilization (Figure 2A,B).
Since we cannot determine whether calcium was mobilized in the red blood cell cytoplasm or inside the parasite by fluorescence spectrometry, we decided to use a confocal microscope to detect fluorescence only inside the parasite (Figure 2C-J).
Although malarial parasites are protected from the external medium by at least two membranes, the red blood cell membrane and the parasitophore vacuole (22), both melatonin and NAS are hydrophobic molecules, which enables them to signal parasites inside red blood cells by mobilizing their intracellular calcium (Figure 2K,L).
The results presented here provide new information related to the identification of molecules that might trigger the developmental processes in malaria parasites. We showed that a direct precursor of melatonin, NAS, is also able to mobilize intracellular calcium in Plasmodium.
In addition we showed that both hormones are able to cross at least two membranes, the red blood cell membrane and the parasitophore vacuole, before signaling the parasite. The hydrophobic nature of melatonin and NAS is an essential feature enabling them to act as signaling molecules in parasites that are not directly exposed to the extracellular milieu.
The fact that the host melatonin starts signal transduction pathways in these parasites shows a closer interaction between parasite and host. To adapt to the host, the parasite might utilizes the molecular host machinery for successful division and growth.
Ca2+ mobilization by N-acetyl-serotonin (NAS) in intact P. chabaudi parasites. Intact parasites at the trophozoite stage were loaded with Fluo-3 AM (see Material and Methods). The reaction medium contained 2 mM Ca2+ in panel A and 12 mM EGTA in panel B. Where indicated, 0.1 mM NAS, 0.1 mM melatonin (MLT) or 100 nM thapsigargin (THG) was added to parasite suspensions (A-G). Addition of 0.1 mM NAS to cells pretreated with 250 mM luzindole (F, 50 min) or with 20 mM U73122 (G, 1 min) did not elicit a Ca2+ response. However, the addition of thapsigargin released Ca2+. Results are typical of at least three independent cell preparations. ER = endoplasmic reticulum.
Ca2+ mobilization by melatonin and N-acetyl-serotonin (NAS) in red blood cells (RBC) infected with P. chabaudi parasites. Infected RBC at the trophozoite stage were loaded with Fluo-3 AM. The reaction medium contained 2 mM Ca2+.Where indicated, 0.1 µM melatonin (MLT) or 0.1 µM NAS was added to the cell suspensions. Experiments were performed with a Hitachi F-4500 spectrofluorometer (A,B) or a Zeiss LSM-510 confocal microscope (C-J). C and G, Phase contrast. Fluorescence of infected RBC before the addition of 0.1 µM MLT (D) or 0.1 µM NAS (H). E and I, Co-localization of the previous images. Fluorescence after the addition of 0.1 µM MLT (F) or 0.1 µM NAS (J). K,L, Fluorescence intensity (DF/F0) normalized as a function of time.
Address for correspondence: C.R.S. Garcia, Departamento de Fisiologia, Instituto de Biociências, USP, 05508-900 São Paulo, SP, Brasil. Fax: +55-11-3091-7594. E-mail: cgarcia@usp.br
C.R.S. Garcia was the recipient of a FAPESP fellowship. Received February 20, 2003. Accepted August 25, 2003.
- 1. Doerig CD (1997). Signal transduction in malaria parasites. Parasitology Today, 13: 297-313.
- 2. Varotti FP, Beraldo FH, Gazarini ML & Garcia CRS (2003). Plasmodium falciparum malaria parasites display a thapsigargin-sensitive Ca2+ pool. Cell Calcium, 33: 137-144.
- 3. Garcia CRS (1999). Calcium homeostasis and signaling in the blood-stage malaria parasite. Parasitology Today, 15: 488-491.
- 4. Garcia CRS, Dluzewski AR, Catalani LH, Burting R, Hoyland J & Mason WT (1996). Calcium homeostasis in intraerythrocytic malaria parasites. European Journal of Cell Biology, 71: 409-413.
- 5. Passos APD & Garcia CRS (1997). Characterization of Ca2+ transport activity associated with a non-mitochondrial calcium pool in the rodent malaria parasite P. chabaudi Biochemistry and Molecular Biology International, 42: 919-925.
- 6. Garcia CRS, Ann SE, Tavares ES, Dluzewski AR, Mason WT & Paiva FB (1998). Calcium acidic pools in intraerythrocytic malaria parasites. European Journal of Cell Biology, 76: 133-138.
- 7. Gazarini ML, Thomas AP, Pozzan T & Garcia CRS (2003). Calcium signalling in a low calcium environment: how the intracellular malaria parasite solves the problem. Journal of Cell Biology, 161: 103-110.
- 8. Hotta CT, Gazarini ML, Beraldo FH, Varotti FP, Lopes C, Markus RP, Pozzan T & Garcia CRS (2000). Calcium dependent modulation by melatonin of the circadian rhythm in malaria parasites. Nature Cell Biology, 2: 466-468.
- 9. Stankov B & Reiter RJ (1990). Melatonin receptors: current status, facts and hypotheses. Life Sciences, 46: 971-982.
- 10. Viven-Roels B, Prevet P, Beck O & Fevre-Montagne M (1995). Identification of melatonin in the compound eye of an insect, the locust (Locusta migratoria) by radioimmunoassay and gas chromatography mass spectrophotometry. Neuroscience Letters, 49: 153-157.
- 11. Dubbels R, Reiter RJ, Klenke E, Goebel A, Schnakenberg E, Ehlers C, Schiwara HW & Schloot W (1995). Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography mass-spectrometry. Journal of Pineal Research, 18: 28-31.
- 12. Pöggeler B & Hardeland R (1994). Detection and quantification of melatonin in a dinoflagellate Gonyaulax polyedra: solutions to the problem of methoxyndole destruction in non-vertebrate material. Journal of Pineal Research, 17: 1-10.
- 13. Arnold JD, Berger A & Martin DC (1969). The role of the pineal in mediating photo-periodic control of growth and division synchrony and capillary sequestration of Plasmodium berghei in mice. Journal of Parasitology, 55: 609-616.
- 14. Hawking F, Gammage K & Worms MJ (1972). The asexual and sexual circadian rhythms of Plasmodium vinckei chabaudi, of P. berghei and of P. gallinaceum Parasitology, 65: 189-201.
- 15. Cambie G, Landau I & Chabaud AG (1990). Niches horaires de trois espèces de Plasmodies coéxistant chez un rongeur de Centrafrique. Comptes Rendus de l'Academie des Sciences, 310: 183-188.
- 16. David PH, Hommel M, Bennichou JC, Eisen HAM & Pereira da Silva LH (1978). Isolation of malaria merozoites: release of Plasmodium chabaudi merozoites: release of Plasmodium chabaudi merozoites from schizonts bound to immobilized concanavalin. Proceedings of the National Academy of Sciences, USA, 75: 5081-5084.
- 17. Eison AS & Mullins UL (1993). Melatonin binding sites are functionally coupled to phosphoinositide hydrolysis in Syrian hamster RPMI1846 melanoma cells. Life Sciences, 53: PL393-PL398.
- 18. Mullins UL & Eison AS (1994). Pharmacologic characterization of melatonin-mediated phosphoinositide hydrolysis in pigeon brain. Journal of Pineal Research, 17: 33-38.
- 19. Popova JS & Dubocovich ML (1995). Melatonin receptor-mediated stimulation of phosphoinositide breakdown in chick brain slices. Journal of Neurochemistry, 64: 130-138.
- 20. Tsim ST, Wong JTY & Wong YH (1997). Calcium ion dependence and the role of inositol phosphates in melatonin-induced encystment of dinoflagellates. Journal of Cell Science, 110: 1387-1393.
- 21. Pozzan T, Rizzuto R, Volpe P & Mendonza PM (1996). Molecular and cellular physiology of intracellular Ca2+ stores. Physiological Reviews, 74: 595-636.
- 22. Kirk K (2001). Membrane transport in the malaria-infected erythrocyte. Physiological Reviews, 81: 495-537.
Correspondence and Footnotes

Publication Dates
-
Publication in this collection
22 Oct 2003 -
Date of issue
Nov 2003
History
-
Received
20 Feb 2003 -
Accepted
25 Aug 2003