Abstract
It has been reported that mental stress causes abnormality of spermiogram parameters. We investigated the effect of psychological stress on the L-arginine-nitric oxide (NO) pathway. Semen samples were collected from 29 healthy fourth semester medical students just before (stress) and 3 months after (non-stress) the final examinations. Psychological stress was measured by the State Anxiety Inventory questionnaire. After standard semen analysis, arginase activity and NO concentration were measured spectrophotometrically in the seminal plasma. Measurements were made in duplicate. During the stress period, sperm concentration (41.28 ± 3.70 vs 77.62 ± 7.13 x 10(6)/mL), rapid progressive motility of spermatozoa (8.79 ± 1.66 vs 20.86 ± 1.63%) and seminal plasma arginase activity (0.12 ± 0.01 vs 0.22 ± 0.01 U/mL) were significantly lower than in the non-stress situation, whereas seminal plasma NO (17.28 ± 0.56 vs 10.02 ± 0.49 µmol/L) was higher compared to the non-stress period (P < 0.001 for all). During stress there was a negative correlation between NO concentration and sperm concentration, the percentage of rapid progressive motility and arginase activity (r = -0.622, P < 0.01; r = -0.425, P < 0.05 and r = -0.445, P < 0.05, respectively). These results indicate that psychological stress causes an increase of NO level and a decrease of arginase activity in the L-arginine-NO pathway. Furthermore, poor sperm quality may be due to excessive production of NO under psychological stress. In the light of these results, we suggest that the arginine-NO pathway, together with arginase and NO synthase, are involved in semen quality under stress conditions.
Psychological stress; Arginase; Nitric oxide; Sperm motility; Sperm count
Braz J Med Biol Res, May 2006, Volume 39(5) 581-588
Effect of psychological stress on the L-arginine-nitric oxide pathway and semen quality
 Correspondence and Footnotes
Correspondence and Footnotes
 S. Eskiocak1, A.S. Gozen2, A. Taskiran1, A.S. Kilic2, M. Eskiocak3 and S. Gulen1
S. Eskiocak1, A.S. Gozen2, A. Taskiran1, A.S. Kilic2, M. Eskiocak3 and S. Gulen1
1Department of Biochemistry, 2Department of Urology, 3Department of Public Health, School of Medicine, Trakya University, Edirne, Turkey
 References
References
Correspondence and Footnotes
Correspondence and Footnotes
Correspondence and Footnotes
Abstract
It has been reported that mental stress causes abnormality of spermiogram parameters. We investigated the effect of psychological stress on the L-arginine-nitric oxide (NO) pathway. Semen samples were collected from 29 healthy fourth semester medical students just before (stress) and 3 months after (non-stress) the final examinations. Psychological stress was measured by the State Anxiety Inventory questionnaire. After standard semen analysis, arginase activity and NO concentration were measured spectrophotometrically in the seminal plasma. Measurements were made in duplicate. During the stress period, sperm concentration (41.28 ± 3.70 vs 77.62 ± 7.13 x 106/mL), rapid progressive motility of spermatozoa (8.79 ± 1.66 vs 20.86 ± 1.63%) and seminal plasma arginase activity (0.12 ± 0.01 vs 0.22 ± 0.01 U/mL) were significantly lower than in the non-stress situation, whereas seminal plasma NO (17.28 ± 0.56 vs 10.02 ± 0.49 µmol/L) was higher compared to the non-stress period (P < 0.001 for all). During stress there was a negative correlation between NO concentration and sperm concentration, the percentage of rapid progressive motility and arginase activity (r = -0.622, P < 0.01; r = -0.425, P < 0.05 and r = -0.445, P < 0.05, respectively). These results indicate that psychological stress causes an increase of NO level and a decrease of arginase activity in the L-arginine-NO pathway. Furthermore, poor sperm quality may be due to excessive production of NO under psychological stress. In the light of these results, we suggest that the arginine-NO pathway, together with arginase and NO synthase, are involved in semen quality under stress conditions.
Key words: Psychological stress, Arginase, Nitric oxide, Sperm motility, Sperm count
Introduction
Stress is one of the most important health and social problems. It is responsible for various diseases such as atherosclerosis and gastrointestinal system pathology (1,2). Although stress has been implicated as a co-factor in the severity and progression of a number of diseases, the potential role of stress in semen quality has received very little scientific attention. The prevalence of infertile couples is ~10-15% of all couples. The causes of infertility can be divided into four major categories: pure male factor, pure female factor, anomalies detected in both partners, and unexplained infertility (3). Psychological distress has often been suggested as a cause of unexplained infertility (4). Clinical observations and experimental studies have shown that stress suppresses the sexual/reproductive function (5). While the effects of psychological stress on females have been well studied (6,7), comparatively little is known about the impact of emotional stress on males. Male infertility accounts for 40% of all infertility problems (8). Previous studies have indicated that stress has a negative impact on various parameters associated with semen quality, including sperm concentration, motility and morphology (9-11). The degree of psychosocial stress has been found to be higher in infertile couples than in fertile controls (4,10). An inverse relationship between semen quality and psychological stress has been detected in infertile subjects undergoing in vitro fertilization (9,11).
Nitric oxide (NO) is a highly reactive free radical gas which has been shown to possess an extraordinary variety of biological functions. NO is synthesized from L-arginine by a family of isoenzymes known as the NO synthases (NOS, EC 1.14.13.39) (12). NOS has been found in the male reproductive system (13). NO has a dual function, being both a cytotoxic and necessary molecule for normal sperm production. Under physiological conditions, NO plays an important role in normal sperm production and motility. Low NO concentrations have been shown to enhance sperm motility (14), whereas high NO concentrations reduce it (15).
Arginase (EC 3.5.3.1) is an enzyme of the urea cycle that catalyzes the hydrolysis of L-arginine to urea and ornithine in liver. Extrahepatic arginase may play a role in reactions other than those of the urea cycle. In mammals, it is also found in the prostate, testis, epididymis, seminal plasma, and human sperm cells (16). Since arginase is an arginine-depleting enzyme, it is an important part of the cellular arginine regulatory system affecting NOS activity (17).
Most studies investigating the association between psychological stress and semen quality have lacked information on biochemical parameters and the effect of stress alone on semen quality. Infertility itself and/or its therapy may be stressors; on the other hand, stress may also cause infertility. For this reason, we investigated the effect of stress alone on the semen quality and L-arginine-NO pathway. We studied semen samples from medical students, because medical students in Turkey must complete an intensive program, especially in the 2nd year. Therefore, stress is very common among medical students during the course of their training and they are more stressed during final examinations.
Material and Methods
Patients and samples
The Institutional Ethics Committee approved the study according to the principles of the Declaration of Helsinki and informed written consent was obtained from each subject. All subjects were nonsmokers and none were currently taking any medication. Their cortisol, total testosterone, prolactin, follicle-stimulating hormone, and luteinizing hormone levels were within reference values (18.14 ± 3.98 µg/dL, 517.38 ± 95.45 ng/dL and 5.62 ± 2.06 ng/mL, 3.03 ± 0.91 and 2.84 ± 1.33 mIU/mL, respectively). Subjects with diabetes mellitus, renal and hepatic disease or hormonal dysfunction and those suffering from any acute infection were excluded. Oligospermic subjects (spermatozoa density <20 x 106/mL) were also excluded from the study in order to eliminate the possible negative effects of unknown additional pathologies and to limit the study only to healthy individuals. None of the subjects were exposed to any other stress factor within the last 3 months before the exam stress period. Subjects who were exposed to any other stress factor such as the death of a family member between periods were also excluded from the study. Semen samples were obtained from 29 healthy volunteers (age: 19.97 ± 0.98 years), fourth semester medical students, just before the final exam (stress period) and then again about 3 months (10.24 ± 0.87 weeks) later, after vacation (non-stress period). The 3-month interval is necessary because of the duration of the spermatogenic cycle and the sperm transit time in humans is approximately 74 days (18). No subjects performed exercise or received dietary therapy during the study period. Samples were collected at the clinic by masturbation into a sterile glass container, following sexual abstinence for at least 48 h. The abstinence time of each subject was the same for each sample collection. The mean abstinence time was 3 days (range: 2-4 days).
After semen liquefaction at 37ºC, spermiograms were obtained by the same two trained observers, as described earlier, according to the WHO guidelines (19). Spermiograms included semen volume (mL), sperm motility (%), sperm concentration (millions per mL), abnormal morphologic features (%), and pH. A phase-contrast microscope (Olympus, Tokyo, Japan) was used for semen analysis. Sperm concentration was measured in a Neubauer counting chamber. The sperm motility assay was performed exactly after 2 h during both the stress and non-stress collections. Sperm motility was classified into four categories: rapid progressive, borderline progressive, non-progressive, and immobility. The motility index as the indicator of motility quality was derived from the formula: % total progressive motility/100 x sperm count; the index is used to demonstrate progressive motile sperm concentration per mL of semen. Total progressive motility was defined as the percentage of rapid progressive motile plus borderline progressive motile spermatozoa. Morphology was measured by determining the percentage of abnormal forms in the sample. The Diff-Quick stain was used for the examination of morphological features. A total of 100 cells per sample were examined to determine the cells with normal morphology, which was characterized by a normal head, middle piece and tail.
Stress questionnaire
Psychological stress was measured by the State Anxiety Inventory, which is widely used for assessing state or acute anxiety (20). The State Anxiety Inventory, translated and validated by Oner and LeComple (21), was completed by all participants after collection of the semen specimen. The State Anxiety Inventory asks the subject to describe how he feels "right now" by responding to 20 questions with a 4-point response format from "not at all" (score 1) to "extremely" (score 4). Total scores range from 20 to 80, with higher scores indicating greater anxiety. This measure has been shown to have high reliability and good construct validity.
Biochemical analysis
Semen samples were centrifuged at 10,000 g for 15 min at +4ºC and seminal plasma was used for analysis. Nitrate (NO3-) and nitrite (NO2-) are the stable end products of NO. To estimate the amount of NO production, NO metabolites in the seminal plasma were measured by the method of Cortas and Wakid (22) based on the colorimetric reaction of Griess. Since very little or no NO2- is normally found in plasma, no attempt was made to differentiate between the amounts of NO3- and NO2- and therefore the results are reported as NO. Seminal plasma was preincubated with urease at 20ºC, pH 7.0, for the hydrolysis of endogenous urea. Arginase activity was then determined at 37ºC, pH 9.7, by the spectrophotometric method described by Geyer and Dabich (23). One unit of arginase activity is defined as one micromole of urea released per minute at 37ºC.
Chemicals
High purity grade reagents from Sigma (St. Louis, MO, USA) and Merck (Darmstadt, Germany) were used for NO and arginase measurements.
Data analysis
Data are reported as means ± SEM. Comparisons of the data between the stress and non-stress periods were made by the t-test for paired samples. When the data were not normally distributed, the Wilcoxon signed-rank test was used for analysis. Correlations between parameters at each stage were determined by the Pearson correlation test. A P value of less than 0.05 was considered to be statistically significant.
Results
The values of semen parameters, state anxiety scores, NO levels, and arginase activities of seminal plasma during the stress and non-stress periods are presented in Table 1. The mean semen volume and the mean semen pH were unchanged during both periods. The concentration and rapid progressive motility of spermatozoa in the stress situation were significantly lower than those found in the non-stress (P < 0.001 for both). The motility index during stress was significantly decreased compared to the non-stress situation (P < 0.001). The percentage of sperm with abnormal morphology tended to increase with stress, but this change was not significant. State anxiety scores were significantly higher during stress than during the non-stress period (P < 0.01). During stress, seminal plasma NO level was higher, whereas arginase activity of seminal plasma was lower, compared to the non-stress period (P < 0.001 for both).
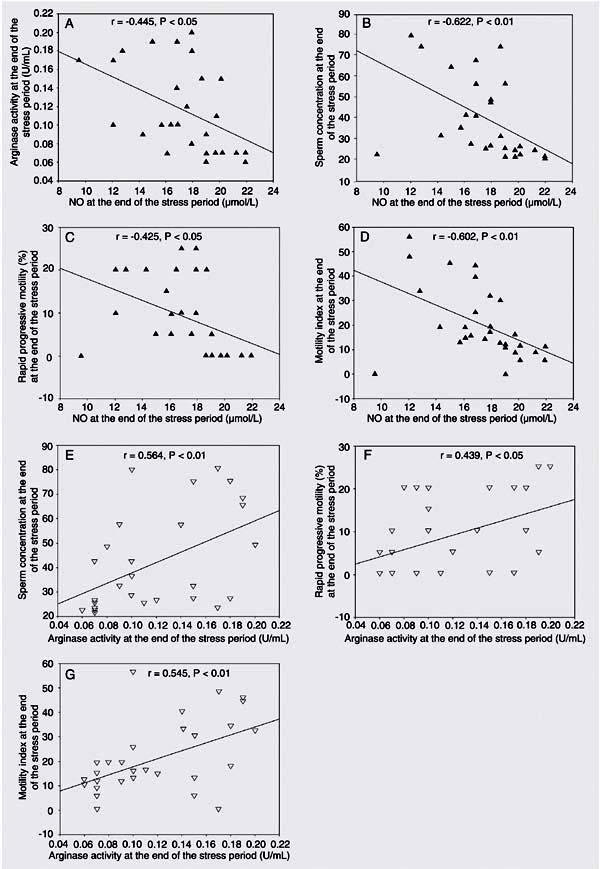
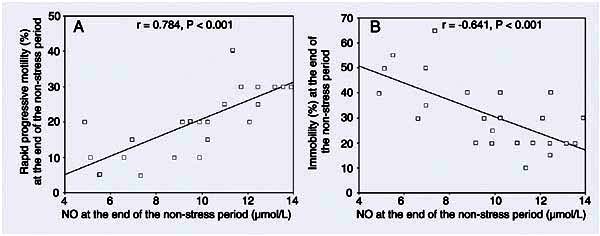
The correlations between parameters during both periods are illustrated in Figures 1 and 2. During the stress period, the significant increase in seminal plasma NO levels, but the significant decrease in seminal arginase activity were found to be related to poor sperm quality including concentration, rapid progressive motility and motility index. There was a negative correlation between the level of seminal plasma NO and arginase activity (r = 0.445, P < 0.05). During the non-stress period, there was a positive correlation between seminal NO level and rapid progressive motility (r = 0.784, P < 0.001). There was also a negative correlation between seminal NO level and the percentage of immobile sperm (r = -0.641, P < 0.001).
Stress period correlations between nitric oxide and arginase activity (A), sperm concentration (B), rapid progressive motility (C), motility index (D) and arginase activity and sperm concentration (E), rapid progressive motility (F), and motility index (G). The correlations in panels A, C, and F were calculated by the Pearson test, and the correlations in panels B, D, E and G by the Spearman test. r and P values are given at the top of each panel.
Non-stress period correlations between nitric oxide (NO) levels and rapid progressive motility and between NO levels and immobility. r and P values are given at the top of each panel. Panel A was calculated by the Pearson test and panel B by the Spearman test.
Semen quality, state anxiety scores, seminal plasma nitric oxide levels, and arginase activity determined in medical students during stress and non-stress periods.
Discussion
NO is a highly reactive, short-lived, lipophilic molecule with a half-life of a few seconds, which makes it difficult to measure it (12). Several research groups have studied the effect of NO on spermatogenesis and sperm function. NO reduces or inhibits sperm motility, while NOS inhibitors, which inhibit the formation of NO, prevent the reduction of sperm motility, indicating a role for the L-arginine-NO pathway in the modulation of sperm motility and motility survival (13,15). It has also been shown that low concentrations of NO may have some physiological role in fertilization through enhanced capacitation and zona pellucida binding (24). Donnelly et al. (25) found that sperm motility is affected negatively by both excessive and reduced endogenous NO production. They suggested that endogenous NO appears to have an important role in the maintenance of sperm motility, when carefully regulated. It has been reported that the abnormal morphology of spermatozoa is not associated with elevated NO production and that the effect of reactive oxygen species (ROS) on sperm membranes depresses sperm function. Aksoy et al. (26) reported that the production of NO is increased in patients with varicocele, and NO is negatively correlated with sperm motility, morphology and concentration.
In our study, seminal plasma NO levels were higher during stress than during the non-stress situation, and were negatively correlated with concentration, rapid progressive motility and motility index of spermatozoa. These data suggest that higher NO levels in seminal plasma are associated with poor sperm quality. During the non-stress period, there was a positive correlation between seminal plasma NO and progressive motility, whereas seminal plasma NO was negatively correlated with the percentage of immobile sperm. These data suggest that, under physiological conditions, low amounts of NO are necessary for normal sperm function. In a non-stress situation, NO may neutralize free radicals and thereby prevent the reduction of sperm motility mediated by ROS. However, excessive generation of NO under stress conditions may be toxic and could reduce sperm motility. Sperm motility is maintained by high levels of adenosine triphosphate. It is known that NO can reduce adenosine triphosphate levels in cells by inhibiting glycolysis and the electron-transport chain (27). Excessive NO also contributes to the formation of peroxynitrite, a highly toxic anion of peroxidation. Peroxynitrite is not a free radical because the unpaired electrons of NO and O2.- combine to form a new N-O bond in peroxynitrite, but it is a strong one- or two-electron oxidant and nitrating agent. Even at physiological pH, the relative stability of peroxynitrite allows it to diffuse for a considerable distance on a cellular scale, and even to cross cell membranes (28). Peroxynitrite can react rapidly with protein, lipids and DNA (29,30). Spermatozoa are highly susceptible to damage induced by ROS. This is due to the high content of polyunsaturated fatty acids within the plasma membranes and a low concentration of scavenging enzymes within the cytoplasm (8). There is growing evidence that oxidative stress significantly impairs sperm function and plays a major role in the etiology of male infertility. The production of abnormal levels of ROS is thought to be involved in many aspects of male infertility, with spermatozoa rendered dysfunctional by lipid peroxidation and altered membrane function, together with impaired metabolism, morphology, motility, and fertility. The incidence of spontaneous pregnancy is negatively correlated with the generation of ROS (31). Many studies have demonstrated the association of lipid peroxidation with middle piece abnormality, decreased sperm count and motility, and loss of the capacity of the spermatozoon to undergo the acrosome reaction and fertilization.
Elgun et al. (32) have shown that sperm cells from infertile men with oligospermia have a significantly higher arginase activity than controls. They reported a positive correlation between sperm motility and arginase activity in the infertile group in both seminal plasma and sperm cells. In the present study, seminal plasma arginase levels were lower during the stress period than in the non-stress situation. Furthermore, seminal plasma arginase activity correlated positively with sperm concentration, rapid progressive motility and motility index. These data suggest that a lower arginase activity in seminal plasma is associated with poor sperm quality.
The rate of NO production may be critically dependent on the availability of arginine. Reduced arginine availability may cause reduced NO synthesis. Arginine availability for NO synthesis is regulated by de novo arginine synthesis, cellular arginine transport and arginase activity (33). Recent studies have reported that arginase modulates NOS activity via arginine consumption (17,34). On the other hand, it has been reported that arginase is inhibited by the nitrite, a stable end product of NO (35). We demonstrated a negative correlation between seminal plasma NO level and arginase activity during stress. However, we did not find a correlation between these parameters during the non-stress situation. Our results suggest that the high level of NO is associated with arginase activity. Under physiological conditions, the concentration of circulating arginine is approximately 100 µM and the tissue concentration ranges from 100 to 1000 µM (17). The Km of the NOS isoforms for arginine lies between 2 and 20 µM, while the Km of arginase is approximately 1 mM. The 1000-fold difference in Km between arginase and NOS for arginine suggests that, under most conditions, arginase cannot compete with NOS for arginine. Depending on the local concentration of arginase, the enzyme can limit the availability of arginine for NO synthesis. The present study suggests that NO and the end products of NO are excessively produced during the stress period. In our opinion, arginase may be inhibited by the end products of NO.
The concentration of NO is important for sperm quality and it especially affects sperm motility. Under stress conditions, NO content of seminal plasma was negatively correlated with sperm quality parameters. Our data suggest that NO plus stress are interrelated and seminal plasma NO levels can affect sperm quality with consequences on fertilization. In the light of these evaluations, we may suggest that the arginine-NO pathway, together with arginase and NOS, may be involved in semen quality under stress conditions.
Received October 14, 2004. Accepted December 16, 2005.
- 1. Schellenbaum GD, Rea TD & Smith NL (2002). Mental stress-induced ischemia and all-cause mortality in patients with coronary artery disease. Circulation, 106: 183-184.
- 2. Swain MS (2000). Stress and the gastrointestinal tract. I. Stress and hepatic inflammation. American Journal of Physiology, 279: 1135-1138.
- 3. The ESHRE Capri Workshop Group (1996). Infertility revisited: the state of the art today and tomorrow. Human Reproduction, 11: 1779-1807.
- 4. Harrison RF, O'Moore RR & O'Moore AM (1986). Stress and fertility: some modalities of investigation and treatment in couples with unexplained infertility in Dublin. International Journal of Fertility, 31: 153-159.
- 5. Yoon H, Chung WS, Park YY et al. (2005) Effects of stress on female rat sexual function. International Journal of Impotence Research, 17: 33-38.
- 6. Hjollund NH, Jensen TK, Bonde JP et al. (1999). Distress and reduced fertility: a follow-up study of first-pregnancy planners. Fertility and Sterility, 72: 47-53.
- 7. Harlow CR, Fahy UM, Talbot WM et al. (1996). Stress and stress-related hormones during in vitro fertilization treatment. Human Reproduction, 11: 274-279.
- 8. Sharma RK, Pasqualotto FF, Nelson DR et al. (1999). The reactive oxygen species-total antioxidant capacity score is a new measure of oxidative stress to predict male infertility. Human Reproduction, 14: 2801-2807.
- 9. Pook M, Krause W & Rohrle B (1999). Coping with infertility: distress and changes in sperm quality. Human Reproduction, 14: 1487-1492.
- 10. Boivin J, Shoog-Svanberg A, Andersson L et al. (1998). Distress level in men undergoing intracytoplasmic sperm injection versus in vitro fertilization. Human Reproduction, 13: 1403-1406.
- 11. Clarke RN, Klock SC, Geoghegan A et al. (1999). Relationship between psychological stress and semen quality among in vitro fertilization patients. Human Reproduction, 14: 753-758.
- 12. Davies MG, Fulton GJ & Hagen PO (1995). Clinical biology of nitric oxide. British Journal of Surgery, 82: 1598-1610.
- 13. Zini A, O'Bryan MK & Schlegel PN (2001). Nitric oxide synthase activity in human seminal plasma. Urology, 58: 85-89.
- 14. Herrero MB, de Lamirande E & Gagnon C (2001). Tyrosine nitration in human spermatozoa: a physiological function of peroxynitrite, the reaction product of nitric oxide and superoxide. Molecular Human Reproduction, 7: 913-921.
- 15. Rosselli M, Dubey RK, Imthurn B et al. (1995). Effects of nitric oxide on human spermatozoa: evidence that nitric oxide decreases sperm motility and induces sperm toxicity. Human Reproduction, 10: 1786-1790.
- 16. Keskinege A, Elgun S & Yilmaz E (2001). Possible implications of arginase and diamine oxidase in prostatic carcinoma. Cancer Detection and Prevention, 25: 76-79.
- 17. Hallemeesch MM, Lamers WH & Deutz NE (2002). Reduced arginine availability and nitric oxide production. Clinical Nutrition, 21: 273-279.
- 18. Turek P (2000). Male infertility. In: Tanagho EM & McAninch JW (Editors), Smith's General Urology. McGraw-Hill Co., New York.
- 19. World Health Organization (1993). WHO Laboratory Manual for the Examination of Human Sperm and Semen Cervical Mucus Interaction. Cambridge University Press, New York.
- 20. Spielberger CD, Gorsuch RL & Lushene RE (1970). STAI Manual for the State-Trait Anxiety Inventory (Self-Evaluation Questionnaire). Consulting Psychologists Press, Palo Alto, CA, USA.
- 21. Oner N & LeComple A (1998). DurumLuluk-Süreklilik Kaygi Envanteri: El Kitabi. Boqaziçi Üniversitesi Matbaasi, Ästanbul, Turkey.
- 22. Cortas NK & Wakid NW (1990). Determination of inorganic nitrate in serum and urine by a kinetic cadmium-reduction method. Clinical Chemistry, 36: 440-443.
- 23. Geyer JW & Dabich D (1971). Rapid method for determination of arginase activity in tissue homogenates. Analytical Biochemistry, 39: 412-417.
- 24. Sengoku K, Tamate K, Yoshida T et al. (1998). Effects of low concentrations of nitric oxide on the zona pellucida binding ability of human spermatozoa. Fertility and Sterility, 69: 522-527.
- 25. Donnelly ET, Lewis SE, Thompson W et al. (1997). Sperm nitric oxide and motility: the effects of nitric oxide synthase stimulation and inhibition. Molecular Human Reproduction, 3: 755-762.
- 26. Aksoy H, Aksoy Y, Ozbey I et al. (2000). The relationship between varicocele and semen nitric oxide concentrations. Urological Research, 28: 357-359.
- 27. Brown GC & Borutaite V (2002). Nitric oxide inhibition of mitochondrial respiration and its role in cell death. Free Radical Biology and Medicine, 33: 1440-1450.
- 28. Beckman JS (2002). Protein tyrosine nitration and peroxynitrite. FASEB Journal, 16: 1144.
- 29. Sies H & Arteel GE (2000). Interaction of peroxynitrite with selenoproteins and glutathione peroxidase mimics. Free Radical Biology and Medicine, 28: 1451-1455.
- 30. Herrero MB & Gagnon C (2001). Nitric oxide: a novel mediator of sperm function. Journal of Andrology, 22: 349-356.
- 31. Aitken RJ, Irvine DS & Wu FC (1991). Prospective analysis of sperm-oocyte fusion and reactive oxygen species generation as criteria for the diagnosis of infertility. American Journal of Obstetrics and Gynecology, 164: 542-551.
- 32. Elgun S, Kacmaz M, Sen I et al. (2000). Seminal arginase activity in infertility. Urological Research, 28: 20-23.
- 33. Wu G & Morris SM (1998). Arginine metabolism: nitric oxide and beyond. Biochemical Journal, 336: 1-7.
- 34. Chang C (1998). Arginase modulates nitric oxide production in activated macrophages. American Journal of Physiology, 274: H342-H348.
- 35. Hrabak A, Bajor T, Temesi A et al. (1996). The inhibitory effect of nitrite, a stable product of nitric oxide (NO) formation, on arginase. FEBS Letters, 390: 203-206.
Correspondence and Footnotes

Publication Dates
-
Publication in this collection
20 Apr 2006 -
Date of issue
May 2006
History
-
Accepted
16 Dec 2005 -
Received
14 Oct 2004