Abstract
Multiple cell membrane alterations have been reported to be the cause of various forms of hypertension. The present study focuses on the lipid portion of the membranes, characterizing the microviscosity of membranes reconstituted with lipids extracted from the aorta and mesenteric arteries of spontaneously hypertensive (SHR) and normotensive control rat strains (WKY and NWR). Membrane-incorporated phospholipid spin labels were used to monitor the bilayer structure at different depths. The packing of lipids extracted from both aorta and mesenteric arteries of normotensive and hypertensive rats was similar. Lipid extract analysis showed similar phospholipid composition for all membranes. However, cholesterol content was lower in SHR arteries than in normotensive animal arteries. These findings contrast with the fact that the SHR aorta is hyporeactive while the SHR mesenteric artery is hyperreactive to vasopressor agents when compared to the vessels of normotensive animal strains. Hence, factors other than microviscosity of bulk lipids contribute to the vascular smooth muscle reactivity and hypertension of SHR. The excess cholesterol in the arteries of normotensive animal strains apparently is not dissolved in bulk lipids and is not directly related to vascular reactivity since it is present in both the aorta and mesenteric arteries. The lower cholesterol concentrations in SHR arteries may in fact result from metabolic differences due to the hypertensive state or to genes that co-segregate with those that determine hypertension during the process of strain selection.
Smooth muscle cell lipids; Membrane microviscosity; Spontaneously hypertensive rats; Lipid composition; Spin label; Electron spin resonance
Braz J Med Biol Res, September 2009, Volume 42(9) 844-853
Structural properties of lipid reconstructs and lipid composition of normotensive and hypertensive rat vascular smooth muscle cell membranes
T.R. Oliveira1, M.T. Lamy1, U.M. De Paula2, L.L. Guimarães3, M.S. Toledo3, H.K. Takahashi3, A.H. Straus3, C.J. Lindsey2 and  Correspondence and Footnotes
Correspondence and Footnotes
 T.B. Paiva2
T.B. Paiva2
1Departamento de Física Geral, Instituto de Física, Universidade de São Paulo, São Paulo, SP, Brasil
2Departamento de Biofísica, 3Departamento de Bioquímica, Escola Paulista de Medicina, Universidade Federal de São Paulo, São Paulo, SP, Brasil
 References
References
Correspondence and Footnotes
Correspondence and Footnotes
Correspondence and Footnotes
Abstract
Multiple cell membrane alterations have been reported to be the cause of various forms of hypertension. The present study focuses on the lipid portion of the membranes, characterizing the microviscosity of membranes reconstituted with lipids extracted from the aorta and mesenteric arteries of spontaneously hypertensive (SHR) and normotensive control rat strains (WKY and NWR). Membrane-incorporated phospholipid spin labels were used to monitor the bilayer structure at different depths. The packing of lipids extracted from both aorta and mesenteric arteries of normotensive and hypertensive rats was similar. Lipid extract analysis showed similar phospholipid composition for all membranes. However, cholesterol content was lower in SHR arteries than in normotensive animal arteries. These findings contrast with the fact that the SHR aorta is hyporeactive while the SHR mesenteric artery is hyperreactive to vasopressor agents when compared to the vessels of normotensive animal strains. Hence, factors other than microviscosity of bulk lipids contribute to the vascular smooth muscle reactivity and hypertension of SHR. The excess cholesterol in the arteries of normotensive animal strains apparently is not dissolved in bulk lipids and is not directly related to vascular reactivity since it is present in both the aorta and mesenteric arteries. The lower cholesterol concentrations in SHR arteries may in fact result from metabolic differences due to the hypertensive state or to genes that co-segregate with those that determine hypertension during the process of strain selection.
Key words: Smooth muscle cell lipids; Membrane microviscosity; Spontaneously hypertensive rats; Lipid composition; Spin label; Electron spin resonance
Introduction
Abnormalities in the cell membrane lipids are believed to underlie several defects including those that are strongly linked to hypertension, strokes and other cardiovascular diseases (1). Reduced membrane fluidity has been reported in arteries and or blood cells of patients with essential hypertension or in animal models of genetic hypertension (2) including spontaneously hypertensive rats (SHR) (3). In patients with essential hypertension as well as in SHR, fluorescence polarization studies have suggested increased microviscosity of erythrocytes and platelets as compared to normotensive controls (4). In agreement with these findings, using electron spin resonance (ESR) of spin labels, Tsuda et al. (5) confirmed decreased fluidity in erythrocytes of SHR. On the other hand, using fluorescence anisotropy, it was shown that erythrocyte membrane microviscosity does not correlate with blood pressure (6).
However, unlike genetically determined hypertension, secondary hypertension models, such as deoxycorticosterone-salt hypertension, two kidney-one clip Goldblatt hypertension and portal hypertension in rats, are not associated with demonstrable alterations in membrane fluidity (7) or lipid composition (8). This is consistent with the assumption that patients and animals with genetically determined hypertension have a predetermined generalized defect of the cell membrane matrix, in which the transport systems operate, that leads to an increase of bilayer microviscosity. This defect appears to be responsible for the multiple abnormalities of the membrane transport systems known to occur in genetic hypertension (9).
Although compelling, the assumption that increased microviscosity (including that of vascular smooth muscle cells, VSMC) is related to the hypertensive state can be contested by some arguments. First, SHR conducting vessels such as the aorta show decreased vascular reactivity (10), while resistance vessels show increased reactivity to vasoconstrictor agents (11), notwithstanding the purported changes in microviscosity or fluidity of myocyte membranes. Also, this hypothesis does not account for the effects of intracellular ion concentration on the viscoelastic properties of cell membranes. Tsuda and Nishio (2) reported increased viscosity of erythrocyte membranes with high intracellular calcium ion concentrations. Hence, measurements in whole cells do not account for the influence of intracellular ion concentrations on membrane viscosity and other properties (2). In addition, the erythrocyte membranes are not excitable and lack proteins associated with vascular reactivity. Moreover, investigations in cultured cells do not consider the cell density-dependent alterations of membrane properties (12,13) that occur in different stages of cell culture.
Another point is the choice of the Wistar-Kyoto (WKY) strain of normotensive rat as the sole control for SHR in most studies (14). Thus, comparison of SHR to other normotensive strains in addition to the WKY could provide a wider scope for the interpretation of the expected findings. Despite these arguments, the most conspicuous flaw of the hypothesis related to membrane fluidity and hypertension is that little concern is given to the relation between other biophysical and functional properties of smooth muscle membranes that determine vascular reactivity and thus peripheral resistance and a consequent hypertensive state.
In the present investigation, the structure of membranes reconstituted with lipids extracted from resistance (mesenteric) and conductance (aorta) vessels of SHR, normotensive Wistar rats (NWR), and WKY was studied via the ESR signal of nitroxide spin labels incorporated into the membranes. This technique has been extensively used to monitor the viscosity and polarity of the microenvironment where the probes are localized (see, for instance, Ref. 15, and references therein). The nitroxide ESR signal is sensitive to movements in the range of 10-7 to 10-11 s, therefore distinguishing regions of different packing. Moreover, due to the asymmetry of the nitroxide moiety, its ESR signal is also sensitive to bilayer order. Spin labeled phospholipids at different acyl chain positions were used to monitor the bilayer structure at different depths. These labels, similar to biological phospholipids, intercalate into the bilayer, monitoring the structure of the reconstituted lipid membranes.
Material and Methods
Reagents
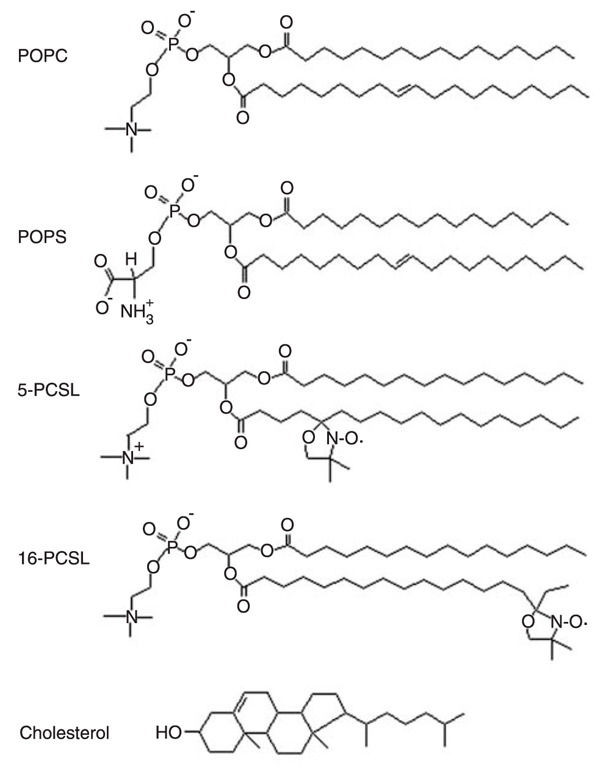
The phospholipids 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) and [1-palmitoyl-2-oleoyl-sn-glycero-3-(phospho-L-serine)] (POPS) cholesterol, and the spin labels 1-palmitoyl-2-(n-doxylstearoyl)-sn-glycero-3-phosphocholine (n-PCSL, n = 5 and 16) were purchased from Avanti Polar Lipids (USA). All products were used as received and their chemical structures are presented in Figure 1. Egg lecithin, (4-(2-hydroxyethyl)-1-piperizineethanesulfonic acid (HEPES), 1,4-piperazinediethane-sulfonic acid (Pipes) and ethylenediaminetetraacetic acid (EDTA) were purchased from Sigma (USA).
Animal handling
Experiments were carried out using 12- to 14-week-old (290-320 g) male Okamoto and Aoki (16) SHR, the normotensive WKY (16), and NWR from the Wistar Institute, Philadelphia, PA, USA, inbred at Escola Paulista de Medicina, Universidade Federal de São Paulo, SP, Brazil. The animals, housed 5 to a cage (60 x 40 x 16 cm), received food and water ad libitum. All experiments were carried out according to existing guidelines on ethics in animal care. The care, handling, housing, and research protocols conformed to the guiding principles for animal experimentation enunciated by the International Association for the Study of Pain and approved by the internal Ethics Committee of Universidade Federal de São Paulo, UNIFESP. Aorta and mesenteric arteries were collected by laparotomy immediately after sacrifice by cerebral concussion and immersed in cooled, pH-equilibrated-buffered Krebs solution for up to 15 min.
Phospholipid extraction and purification
After removal of adherent connective tissue, the aorta and mesenteric arteries collected from 15 to 20 animals per group were weighed, washed with cold phosphate-buffered aerated Krebs solution, pH 7.5, chopped, and minced in chloroform:methanol (2:1, v/v). Phospholipids were collected from supernatants after a 5-h incubation in chloroform:methanol under magnetic stirring and centrifugation at 1500 g for 20 min. The procedure was repeated three times and then twice with isopropanol:hexane:water (55:20:25, v/v/v) (17). The supernatants were pooled, dried under a stream of N2 at 37°C, and suspended in 5 mL chloroform:methanol solution. Non-lipid contaminants, gangliosides and neutral glycolipids with long carbohydrate chains were separated from the phospholipids by Folch's partitioning (18). The Folch lower phase was applied to a silica-gel column (10 x 60 mm) equilibrated with chloroform:methanol (9.5:0.5, v/v). Cholesterol was eluted from the column with 5 volumes of chloroform:methanol (9.5:0.5 and 9.0:1.0, v/v), and the phospholipids were eluted with 5 volumes of chloroform:methanol (7:3 and 2:8, v/v) and 5 volumes of isopropanol:hexane:water (55:20:25, v/v/v). The Folch lower phase was dried and analyzed for phospholipid content by high-performance thin layer chromatography (HPTLC) using chloroform:methanol:methylamine (63:35:10, v/v/v) as the mobile phase, and visualized as blue spots with the Dittmer-Lester reagent. Sterols were determined by HPTLC, developed in chloroform:ethyl ether:acetic acid (97:2.3:0.5, v/v/v), and visualized using the copper acetate reagent. The non-polar phospholipids were measured by HPTLC. Cholesterol was removed from Folch's lower phase phospholipid extracts by silica-gel column chromatography with a 2-step chloroform/methanol elution. The lipids of different fractions were dried under a stream of N2 at 37°C and analyzed by HPTLC on silica gel 60 plates (Merck, Germany).
Phospholipid and cholesterol quantification
Total phospholipid concentrations were assayed as described (19). Aorta and mesenteric artery phospholipid composition and cholesterol content were determined by HPTLC. After staining the HPTLC plates, phospholipids were quantified by densitometry at 525 nm (Shimadzu CS9301, Japan). The commercially available phospholipid standards phosphatidylinositol (PI) and phosphatidylcholine (PC) from Matreya Inc. (Pleasant Gap, USA) and phosphatidylethanolamine (PE), sphingomyelin (SM), phosphatidylserine (PS), and cholesterol from Sigma-Aldrich Corporation (USA) were used as 1-mg/mL solutions.
Gas chromatograph-mass spectrometry
Purified phospholipid samples (20 µg) were dried in Pyrex tubes with Teflon-lined screw caps. Methanolysis was carried out in 1.0 mL 1 M HCl in anhydrous methanol (MeOH/HCl) and heated at 80°C for 16 h. The cooled methanolysates were partitioned three times with equal volumes of hexane. The pooled hexane phases containing fatty acid methyl esters (FAMEs) were dried under N2 at 37°C and resuspended in 100 µL hexane. Gas chromatography-mass spectrometry (GC-MS) analysis was performed at a temperature gradient of 40 to 300°C at 5°C/min followed by a 5-min period at 300°C using a CP-Sil 8 CB column in a CP-3800/1200L mass spectrometer (Varian Inc., USA), with splitless injection. All derivatives were identified by their retention times and mass spectra were compared to their authentic standards and to published data.
Lipid dispersion preparation
A film was formed from the chloroform/methanol solution of the Folch lower phase extracted lipids (phospholipids and cholesterol) with spin labels. The optimal spin label concentrations displaying absence of spin-spin interactions were determined to be 0.6 mol% for the 5-PCSL and 0.2 mol%, for the 16-PCSL. The preparations were dried under a stream of N2 and left under reduced pressure for 2 h to remove any trace of organic solvents. Dispersions were prepared with film hydration by the addition of a buffered Krebs solution, pH 7.4, and vortexed. The final lipid dispersion (10 mM) was loaded onto a capillary for ESR measurements. The egg lecithin reference membrane was made as 10 mM dispersions prepared in HEPES buffer, pH 7.4. The POPC or POPS films were hydrated with 20 mM Pipes, 1 mM EDTA, 150 mM NaCl, 0.002% NaN3, pH 7.4 buffer. Both samples contain the same total lipid concentration (16.4 mg/mL). Cholesterol-saturated membranes of POPC and POPS were also prepared (molar ratio of 50 and 33 mol% relative to total lipid, in POPC and POPS, respectively) (20).
ESR spectroscopy
ESR measurements were performed with a Bruker EMX spectrometer (Germany). The sample temperature was controlled within 0.2°C by a Bruker BVT-2000 variable temperature device. The temperature was checked with a Fluke 51 K/J thermometer (USA) with the probe placed just above the cavity. The sample temperature varied from 5 to 55°C. To ensure thermal equilibrium, before each scan, the sample was left at the desired temperature for at least 5 min. The ESR data were acquired immediately after sample preparation. Field-modulation amplitude of 1 G and microwave power of 10 mW were used. The magnetic field was measured with a Bruker ER 035 NMR Gaussmeter. For the highly anisotropic spectra of 5-PCSL, the isotropic hyperfine splitting was calculated from the equation (21):

[1]
where A// (= Amax) is the maximum hyperfine splitting directly measured in the spectrum (Figure 2), and

[2]
where 2Amin is the measured inner hyperfine splitting (Figure 2) and Axx, Ayy and Azz are the principal values of the hyperfine tensor for doxylpropane (22). Effective order parameters, Seff, were calculated from the equation (23):

[3]
where
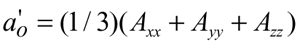
Results and Discussion
Phospholipid and cholesterol quantification
The lipid composition of all membrane preparations used for ESR spectrum studies was quantified. Phospholipid concentrations were the same in resistance and conductance vessels of the hypertensive and normotensive rat strains. The figures ranged from 60 to 40 mol% among the different preparations. On the other hand, cholesterol levels were significantly higher in the arteries of the normotensive rat strains, ranging from 40 to 70 mol% in resistance and conductance vessels of the normotensive strains, while the same vessels of the hypertensive rats had (2 ± 1) x 10 mol% of cholesterol (Table 1). Also, no significant differences were observed in the amount of PE, PC, SM, PS, or PI in the mesenteric or aorta arteries among the 3 rat strains, as shown in Figure 1. In the mesenteric arteries from WKY animals, there was possibly a somewhat lower content of SM and a larger content of PS than in the aorta from the same rat strain.
Despite the biophysical and pharmacological differences between conductance and resistance vessel in membrane resting potentials and vascular reactivity to vasoconstrictor agents or blood pressure (24,25), no significant differences were observed in the concentrations of bulk phospholipid headgroups between the aorta and mesenteric vessels regardless of rat strain. Apparently, the low cholesterol levels of the SHR arteries do not relate to vascular reactivity or directly to the hypertensive state since this lipid had low expression in both resistance and conductance vessels that are hyper- and hyporeactive to vasoconstrictor agents.
Phospholipid fatty acid composition
Mass spectrometry determination of fatty acid composition as methylester (FAMES) derivatives showed similar lipid composition in both aorta and mesenteric artery lipid extracts from hypertensive SHR, normotensive WKY animals or NWR. Stearic and palmitic acids were the major saturated fatty acid components while arachidonic and oleic acids were the main unsaturated fatty acids in the VSMC membranes from the arteries of the 3 rat strains (Table 2). There was no difference in fatty acid composition between the aorta and mesenteric arteries and between any of the three rat strains. However, both the aorta and mesenteric arteries of SHR showed a lower proportion of palmitic acid and a higher proportion of arachidonic acid than the same arteries from normotensive WKY animals or NWR. In contrast to the present findings, decreased levels of linoleic, arachidonic and eicosapentaenoic acids were reported in arteries of stroke-prone hypertensive rats (SHRSP) (26). However, in agreement with our results, a higher proportion of arachidonic acid was found in the plasma membrane of VSMC of SHRSP (27). Table 2 shows the GC-MS analyses of saturated and unsaturated FAMEs from SHR, WKY and NWR pooled aorta or mesenteric arteries. In addition to the mentioned differences in palmitic and arachidonic acid ratios, SHR apparently have a smaller saturated/unsaturated FAME ratio than the arteries of the normotensive WKY or NWR strains, but at this point it is difficult to ascertain whether the difference has biological significance or is statistically significant.
Spin label structural analysis
The ESR spectra of 5- and 16-PCSL spin labels (Figure 2) incorporated into membranes reconstituted from the lipids extracted from the aorta and mesenteric arteries of normotensive NWR and WKY and hypertensive SHR, at 37°C are shown in Figure 3. The nitroxide ESR signal, used to monitor viscosity and polarity in the probe microenvironment (22), can distinguish regions of different lipid packing. The spectra are typical of spin labels incorporated into lipid bilayers (23,28), presenting a much higher anisotropy at the acyl chain 5-C atom position compared to the bilayer core (16-C atom position). This increase in mobility and/or decrease in order towards the bilayer core are the fingerprint of a bilayer. Hence, the extracted lipids in the buffer system are forming vesicles consisting of one or more bilayers. There is no indication of the presence of more than one component in the ESR spectra of 5- or 16-PCSL, at any temperature, for any of the systems studied. Therefore, these membranes seem to be rather homogeneous systems. The spectra of each label incorporated into the different arteries of the different rat strains look very similar. For a better evaluation, specific ESR parameters were measured.
Spectra of 5-PCSL (Figure 3) are typical of fluid bilayers, yielded by the label in a rather ordered location, close to the bilayer-water interface, where the label has fast movement around its long axis (22). In that case, the bilayer can be well analyzed by the effective order parameter Seff (see Material and Methods). As expected, the order parameter decreases as the temperature increases (Figure 4A). 16-PCSL monitors the lipid bilayer packing at the membrane core, where the lipid chains are highly disordered and loose, and an order parameter cannot be directly calculated from the ESR spectra. So, with 16-PCSL, the best experimental parameter to be used over the whole range of temperature is the ratio between the amplitudes of the low and central field lines, corresponding to the nitrogen spin numbers mI = +1 and mI = 0 (h+1 / h0; see Figure 2). This parameter approaches unity as the movement of the label gets faster and more isotropic. As shown in Figure 4B, h+1 / ho increases with temperature as the label environment viscosity decreases.
Figure 4 shows the ESR data of lipid bilayers reconstituted from SHR, WKY, and NWR aorta and smooth muscle mesenteric arteries. The high reproducibility of the method, true for all lipid preparation samples, may be appreciated by the small deviations shown in Figure 3 (three assays were done for each preparation). Surprisingly, the reconstituted lipid bilayers from the aorta and mesenteric arteries of normotensive NWR and WKY, or hypertensive SHR, presented very similar packing, both at the 5th C- and 16th C-atom positions, at all studied temperatures (28,29). To stress their similarity, the values of Seff and h+1 / h0, obtained for 5- and 16-PCSL, respectively, at 37°C, are indicated in the corresponding spectrum in Figure 2.
It is interesting to compare the packing of the bilayers formed by the extracted lipids with those of other lipid membranes frequently used as references such as egg lecithin bilayers, a composition of phosphocholine lipids extracted from egg yolk with different hydrocarbon chains, both saturated and unsaturated (around 50% each), and single phospholipid bilayers made of the zwitterionic lipid POPC and the anionic POPS (saturated bilayers cannot be compared at the temperature interval used, as they present phase transition). Looking at the order at the 5th C-atom position (Seff in Figure 4A), it is clear that the three distinct reference bilayers, lecithin, POPC and POPS, present very similar packing parameters, which are much lower than those calculated for the extracted lipids. At the rather fluid bilayer core, where small packing differences can be well detected, 16-PCSL ESR spectra indicate a higher packing for POPS as compared to POPC and lecithin (lower h+1 / h0 ratios in Figure 4B), which can be attributed to inter-headgroup hydrogen bonding in POPS (29). The membranes containing lecithin, POPC and POPS, have the absence of cholesterol in common, which is known to increase the order and/or decrease the mobility of lipids in the fluid phase (30). When cholesterol is added to POPC and POPS, which show different ability for cholesterol incorporation, 50 and 36 mol% for POPC and POPS, respectively (20,31), the packing of the bilayers increases considerably (increase in the bilayer effective order parameter at the 5th C-atom position, Seff in Figure 4A, and increase in the order and/or decrease in the mobility at the 16th C-atom position, decrease in the h+1 / h0 in Figure 4B). Accordingly, POPC, which incorporates more cholesterol (50 mol%), displays a rather more organized/packed bilayer (higher Seff and lower h+1 / h0).
As shown in Figure 4A, the 5-PCSL Seff values obtained with the extracted lipid bilayers are similar to those obtained with cholesterol-saturated POPS bilayers, and lower than those obtained with POPC (50 mol% cholesterol). This may indicate 30 to 40 mol% as the cholesterol fraction present in the total extracted lipids. However, one should be careful in drawing such a conclusion, since the different lipid composition of the extracted lipids as compared to POPS, which could result in different cholesterol packing effects (32). The 16-PCSL label, which monitors the bilayer core, indicates that the extracted lipid bilayers are even slightly more fluid than cholesterol-saturated POPS bilayers (higher h+1 / h0 values for temperatures above 35°C in Figure 4B).
Biological and biophysical membrane properties and vascular reactivity
The present findings, in consonance with earlier reports (25,26), show that there are no important differences in bulk phospholipids or fatty acid composition of the aorta or mesenteric artery VSMC lipids from normotensive animal strains or rats with genetically determined hypertension. In parallel, membranes prepared from the lipids extracted from the different arteries of the three rat strains, hypertensive or not, were found to be structurally very similar. The fact that our data from reconstituted membranes differ from reports showing increased microviscosity of SHR membranes (2) in intact cell preparations shifts the focus to the role of cell membrane proteins or the protein-lipid rafts in the membrane microenvironment (33,34). Yet, a direct comparison of the reported ESR data (12) with our data is difficult since no ESR spectra are shown in the cited paper that refer only to the calculated parameters. However, despite the generalized increases of membrane rigidity reported for SHR systems, the link between membrane fluidity and the hypertensive state is still unclear. First, the predicted rigidity of SHR cell membranes would not translate per se into increased reactivity of resistance vessels and hypertension. Contrarily, according to most functional predictions, reduced membrane fluidity would signal reduced protein mobility and function leading to impairment of the vascular reactivity of resistance vessels. The second consideration is that the SHR aorta is hyporeactive (24) and the SHR mesenteric artery is hyperreactive (11) to vasoconstrictors notwithstanding the increased microviscosity reported for membranes of both arteries (26). Finally, membrane microviscosity may be strongly influenced by ionic concentrations in the membrane microenvironment (2), in particular those of intracellular calcium ion (Ca2+). Calcium ions have other actions directly related to membrane properties that ultimately determine smooth muscle reactivity (35-37).
Though it is well known that the characteristics of a native membrane are highly dependent on its intrinsic proteins, the result obtained with reconstituted bilayers from artery lipid extracts is a first step towards the understanding of the role of lipids alone in the structural organization of cell membranes.
Interestingly, cholesterol content was found to be lower in cells of both mesenteric and aorta arteries from the hypertensive SHR strain (~30 mol%) than in the corresponding arteries of the normotensive WKY animals and NWR (45-50 mol%; Table 1). Although the lower cholesterol concentrations in the SHR arteries bear no direct relationship to vascular reactivity, since cholesterol concentrations are the same in both aorta and mesenteric arteries, the finding may be considered to be intriguing. Since the lipids of aorta and mesenteric arteries display very similar packing as assessed by the highly sensitive technique of membrane-incorporated spin labels, it is important to keep in mind that different bilayers present different cholesterol saturating limits, as for instance POPC (50 mol%) and POPS (36 mol%) (31). Usually, cholesterol concentrations higher than the saturation limit do not influence the ESR signal (32), and thus excess cholesterol is supposed to be present in cholesterol-rich domains, either in the membrane or as cholesterol crystallites (20,31, and references therein). Thus, the cholesterol of the normotensive rat strain arteries could be localized in such cholesterol-rich domains, inside or outside the membranes, or at specific sites of the membrane, possibly surrounding certain protein structures or protein-lipid raft moieties (38). This could explain the same order/mobility for bilayers with different cholesterol compositions. Comparison with the ESR data obtained with the same labels from other natural systems such as reconstructed membranes of calf lens lipids (33) supports this argument. The latter model yielded ESR spectra comparable to those obtained with POPC + 50 mol% cholesterol, which are typical of labels in a rather organized environment (Figure 4). Furthermore, since cholesterol is known to decrease lipid mobility in the fluid phase (30), the prediction would be that high cholesterol content, and not low cholesterol in vascular smooth muscle, would decrease vascular reactivity. Nevertheless, the biological significance of cholesterol aggregates in cells, albeit attributed to a disorder of cholesterol metabolism, is still a matter of open discussion (39).
There is no indication that differences in phospholipid and fatty acid composition, cholesterol content or bulk lipid packing in hypertensive rat arteries can be related to vascular reactivity and/or hypertension. The significance of the lower cholesterol content in the SHR bears no direct relation to vascular reactivity but may be indirectly related to hypertension as a causal factor or even be caused by the hypertensive state in view of the numerous metabolic characteristics of the SHR. Alternatively, the lower cholesterol concentrations in the SHR arteries could be due to the expression of genes that co-segregate with those that determine hypertension during the process of strain selection.
Structures of lipids and spin labels. POPC = phospholipids 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine; POPS = [1-palmitoyl-2-oleoyl-sn-glycero-3-(phospho-L-serine)]; n-PCSL = 1-palmitoyl-2-(n-doxylstearoyl)-sn-glycero-3-phosphocholine.
Phospholipid composition of aorta and mesenteric artery smooth muscle cell membranes from spontaneously hypertensive rats (SHR), normotensive Wistar-Kyoto rats (WKY) and normotensive Wistar rats (NWR). PE = phosphatidyl-ethanolamine; PC = phosphatidylcholine; SM = sphigmomielin; PS = phosphatidylserine; PI = phosphatidylinositol. The height of the columns indicate the average of 3 replicate assays of a total of 5 (SHR), 3 (WKY) or 4 (NWR) pools of 10 to 30 arteries each. The bars indicate the SEM. *P < 0.05 compared to the respective phospholipid concentration in aorta arteries (ANOVA, Bonferroni's test).
ESR spectra of 5-PCSL and 16-PCSL incorporated into reconstituted lipid bilayers from the aorta and mesenteric arteries of normotensive Wistar rats (NWR), Wistar-Kyoto rats (WKY) and spontaneously hypertensive rats (SHR), at 37°C. The hyperfine splittings Amax and Amin, and the amplitudes of low (h+1) and central (h0) field lines are indicated. Total spectrum width = 100 G. The values shown are those of Seff for 5-PCSL and h+1 / h0 for 16-PCSL spectra.
Temperature dependence of Seff measured on the ESR spectra of 5-PCSL (A), and the h+1 / h0 ratio measured on the ESR spectra of 16-PCSL (B), incorporated into reconstituted lipid bilayers from the aorta (open symbols) or mesenteric arteries (filled symbols) of normotensive Wistar rats (NWR), Wistar-Kyoto rats (WKY) or spontaneously hypertensive rats (SHR). The ESR parameters are compared to those yielded by the labels incorporated into POPC (inverted triangles), POPS (star) saturated with cholesterol (filled inverted triangles and stars), and egg lecithin (S) bilayers. Each point corresponds to the mean value of three different assays, which contained lipids extracted from 10 rats, and the uncertainties are reported as standard deviations and represented by the vertical bars. When smaller than the symbols, the latter are not shown. For other abbreviations, see legend to Figure 1.
Phospholipid and cholesterol content of the aorta and mesenteric arteries of the rat strains studied.
Saturated and unsaturated fatty acid methyl ester content of aorta and mesenteric arteries from the rat strains studied.
Acknowledgments
We thank C.R. Benatti, Departamento de Física Geral, Instituto de Física, Universidade de São Paulo, São Paulo, SP, Brazil, for the POPC and POPS ESR data.
Address for correspondence: T.B. Paiva, Departamento de Biofísica, Universidade Federal de São Paulo, 04023-062 São Paulo, SP, Brasil. Fax: +55-11-5571-5780. E-mail: tbpaiva@biofis.epm.br
Research supported by FAPESP and CNPq. Received October 30, 2008. Accepted June 10, 2009.
The Brazilian Journal of Medical and Biological Research is partially financed by






- 1. Zicha J, Kunes J, Devynck MA. Abnormalities of membrane function and lipid metabolism in hypertension: a review. Am J Hypertens 1999; 12: 315-331.
- 2. Tsuda K, Nishio I. Membrane fluidity and hypertension. Am J Hypertens 2003; 16: 259-261.
- 3. Dominiczak AF, Lazar DF, Das AK, Bohr DF. Lipid bilayer in genetic hypertension. Hypertension 1991; 18: 748-757.
- 4. Le Quan Sang KH, Montenay-Garestier T, Devynck MA. Alterations of platelet membrane microviscosity in essential hypertension. Clin Sci 1991; 80: 205-211.
- 5. Tsuda K, Minatogawa Y, Iwahashi H, Nishio I, Kido R, Masuyama Y. Spin-labelling study of biomembranes in spontaneously hypertensive rats: calcium- and calmodulin-dependent regulation. Clin Exp Pharmacol Physiol Suppl 1995; 22: S234-S236.
- 6. McLaren Y, Kreutz R, Lindpaintner K, Bohr DF, Ganten D, Reid JL, et al. Membrane microviscosity does not correlate with blood pressure: a cosegregation study. J Hypertens 1993; 11: 25-30.
- 7. Orlov SN, Gulak PV, Litvinov IS, Postnov YV. Evidence of altered structure of the erythrocyte-membrane in spontaneously hypertensive rats. Clin Sci 1982; 63: 45.
- 8. Ma Z, Meddings JB, Lee SS. Cardiac plasma membrane physical properties and beta-adrenergic receptor function are unaltered in portal-hypertensive rats. Hepatology 1995; 22: 188-193.
- 9. Dominiczak AF, Bohr DF. The primacy of membrane microviscosity in genetic hypertension. Am J Hypertens 1991; 4: 963-969.
- 10. Shibata S, Kurahashi K, Kuchii M. A possible etiology of contractility impairment of vascular smooth muscle from spontaneously hypertensive rats. J Pharmacol Exp Ther 1973; 185: 406-417.
- 11. Feres T, Borges AC, Silva EG, Paiva AC, Paiva TB. Impaired function of alpha-2 adrenoceptors in smooth muscle of mesenteric arteries from spontaneously hypertensive rats. Br J Pharmacol 1998; 125: 1144-1149.
- 12. Martens JR, O'Connell K, Tamkun M. Targeting of ion channels to membrane microdomains: localization of KV channels to lipid rafts. Trends Pharmacol Sci 2004; 25: 16-21.
- 13. Blennerhassett MG, Kannan MS, Garfield RE. Density-dependent hyperpolarization in cultured aortic smooth muscle cells. Am J Physiol 1989; 256: C644-C651.
- 14. Rapp JP. Use and misuse of control strains for genetically hypertensive rats. Hypertension 1987; 10: 7-10.
- 15. Benatti CR, Ruysschaert JM, Lamy MT. Structural characterization of diC14-amidine, a pH-sensitive cationic lipid used for transfection. Chem Phys Lipids 2004; 131: 197-204.
- 16. Okamoto K, Aoki K. Development of a strain of spontaneously hypertensive rats. Jpn Circ J 1963; 27: 282-293.
- 17. Straus AH, Levery SB, Jasiulionis MG, Salyan ME, Steele SJ, Travassos LR, et al. Stage-specific glycosphingolipids from amastigote forms of Leishmania (L.) amazonensis. Immunogenicity and role in parasite binding and invasion of macrophages. J Biol Chem 1993; 268: 13723-13730.
- 18. Folch J, Lees M, Sloane Stanley GH. A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 1957; 226: 497-509.
- 19. Rouser G, Fkeischer S, Yamamoto A. Two dimensional thin layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 1970; 5: 494-496.
- 20. Epand RM, Bach D, Borochov N, Wachtel E. Cholesterol crystalline polymorphism and the solubility of cholesterol in phosphatidylserine. Biophys J 2000; 78: 866-873.
- 21. Gaffney BJ. Practical considerations for the calculation of order parameters for fatty acid or phospholipid spin labels in membranes. In: Berliner LJ (Editor), Spin labeling. Theory and applications. New York: Academic Press; 1976. p 567-571.
- 22. Hubbell WL, McConnell HM. Molecular motion in spin-labeled phospholipids and membranes. J Am Chem Soc 1971; 93: 314-326.
- 23. Schindle H, Seelig J. EPR-spectra of spin labels in lipid bilayers. J Chem Phys 1973; 59: 1841-1850.
- 24. Farias NC, Feres T, Paiva AC, Paiva TB. Lys-[Leu8,des-Arg9]-bradykinin blocks lipopolysaccharide-induced SHR aorta hyperpolarization by inhibition of Ca(++)- and ATP-dependent K+ channels. Eur J Pharmacol 2004; 498: 163-169.
- 25. Silva EG, Frediani-Neto E, Ferreira AT, Paiva AC, Paiva TB. Role of Ca(+)-dependent K-channels in the membrane potential and contractility of aorta from spontaneously hypertensive rats. Br J Pharmacol 1994; 113: 1022-1028.
- 26. Nara Y, Sato T, Mochizuki S, Mano M, Horie R, Yamori Y. Metabolic dysfunction in smooth-muscle cells of spontaneously hypertensive rats. J Hypertens 1986; 4: S105-S107.
- 27. Dominiczak AF, McLaren Y, Kusel JR, Ball DL, Goodfriend TL, Bohr DF, et al. Lateral diffusion and fatty acid composition in vascular smooth muscle membrane from stroke-prone spontaneously hypertensive rats. Am J Hypertens 1993; 6: 1003-1008.
- 28. Benatti CR, Epand RM, Lamy MT. Low cholesterol solubility in DODAB liposomes. Chem Phys Lipids 2007; 145: 27-36.
- 29. Petrache HI, Tristram-Nagle S, Gawrisch K, Harries D, Parsegian VA, Nagle JF. Structure and fluctuations of charged phosphatidylserine bilayers in the absence of salt. Biophys J 2004; 86: 1574-1586.
- 30. Hsia JC, Schneider H, Smith IC. A spin label study of the influence of cholesterol on phospholipid multibilayer structures. Can J Biochem 1971; 49: 514-522.
- 31. Bach D, Wachtel E. Phospholipid/cholesterol model membranes: formation of cholesterol crystallites. Biochim Biophys Acta 2003; 1610: 187-197.
- 32. Kusumi A, Subczynski WK, Pasenkiewicz-Gierula M, Hyde JS, Merkle H. Spin-label studies on phosphatidylcholine-cholesterol membranes: effects of alkyl chain length and unsaturation in the fluid phase. Biochim Biophys Acta 1986; 854: 307-317.
- 33. Widomska J, Raguz M, Dillon J, Gaillard ER, Subczynski WK. Physical properties of the lipid bilayer membrane made of calf lens lipids: EPR spin labeling studies. Biochim Biophys Acta 2007; 1768: 1454-1465.
- 34. Swamy MJ, Ciani L, Ge M, Smith AK, Holowka D, Baird B, et al. Coexisting domains in the plasma membranes of live cells characterized by spin-label ESR spectroscopy. Biophys J 2006; 90: 4452-4465.
- 35. Liu Y, Pleyte K, Knaus HG, Rusch NJ. Increased expression of Ca2+-sensitive K+ channels in aorta of hypertensive rats. Hypertension 1997; 30: 1403-1409.
- 36. Bravo JCR, Lindsey CJ, Farias NC, Feres T, Paiva ACM, Paiva TB. In vivo ACE-inhibitor treatment normalizes hyperpolarization of isolated SHR aorta rings. Int J Atheroscler 2006; 1: 235-241.
- 37. Toyoda Y, Shima H, Sasajima H, Nishio I. Increased calcium sequestration by sarcoplasmic reticulum in small muscular arteries in young spontaneously hypertensive rats. Clin Exp Pharmacol Physiol Suppl 1995; 22: S223-S224.
- 38. Epand RM. Cholesterol and the interaction of proteins with membrane domains. Prog Lipid Res 2006; 45: 279-294.
- 39. Geng YJ, Phillips JE, Mason RP, Casscells SW. Cholesterol crystallization and macrophage apoptosis: implication for atherosclerotic plaque instability and rupture. Biochem Pharmacol 2003; 66: 1485-1492.
Correspondence and Footnotes

Publication Dates
-
Publication in this collection
31 July 2009 -
Date of issue
Sept 2009
History
-
Accepted
10 June 2009 -
Received
30 Oct 2008