
Abstract
Peroxisome proliferator activator receptor-gamma (PPARγ) is a ligand-activated transcriptional factor involved in the carcinogenesis of various cancers. Insulin-like growth factor-binding protein-3 (IGFBP-3) is a tumor suppressor gene that has anti-apoptotic activity. The purpose of this study was to investigate the anticancer mechanism of PPARγ with respect to IGFBP-3. PPARγ was overexpressed in SNU-668 gastric cancer cells using an adenovirus gene transfer system. The cells in which PPARγ was overexpressed exhibited growth inhibition, induction of apoptosis, and a significant increase in IGFBP-3 expression. We investigated the underlying molecular mechanisms of PPARγ in SNU-668 cells using an IGFBP-3 promoter/luciferase reporter system. Luciferase activity was increased up to 15-fold in PPARγ transfected cells, suggesting that PPARγ may directly interact with IGFBP-3 promoter to induce its expression. Deletion analysis of the IGFBP-3 promoter showed that luciferase activity was markedly reduced in cells without putative p53-binding sites (-Δ1755, -Δ1795). This suggests that the critical PPARγ-response region is located within the p53-binding region of the IGFBP-3 promoter. We further demonstrated an increase in PPARγ-induced luciferase activity even in cells treated with siRNA to silence p53 expression. Taken together, these data suggest that PPARγ exhibits its anticancer effect by increasing IGFBP-3 expression, and that IGFBP-3 is a significant tumor suppressor.
PPARγ; IGFBP-3; Apoptosis; Gastric cancer
Introduction
Peroxisome proliferator-activated receptors (PPARs) are multifunctional nuclear hormone receptors that influence cell growth, cell differentiation, and cell death (11 Robinson-Rechavi M, Carpentier AS, Duffraisse M, Laudet V. How many nuclear hormone receptors are there in the human genome? Trends Genet 2001; 17: 554-556, doi: 10.1016/S0168-9525(01)02417-9.
https://doi.org/10.1016/S0168-9525(01)02...
). There are three PPAR isoforms, PPARα, PPARβ/δ, and PPARγ, and of these, PPARγ is mainly expressed in adipose tissues. It regulates lipid and glucose metabolism and adipocyte differentiation (22 Schoonjans K, Staels B, Auwerx J. The peroxisome proliferator activated receptors (PPARS) and their effects on lipid metabolism and adipocyte differentiation. Biochim Biophys Acta 1996; 1302: 93-109, doi: 10.1016/0005-2760(96)00066-5.
https://doi.org/10.1016/0005-2760(96)000...
). Because of its role in metabolic regulation, PPARγ has become a molecule of interest in many research areas, serving as a putative therapeutic target in diseases such as type 2 diabetes mellitus, dyslipidemia, inflammatory diseases, and hypertension (33 Gervois P, Fruchart JC, Staels B. Inflammation, dyslipidaemia, diabetes and PPars: pharmacological interest of dual PPARalpha and PPARgamma agonists. Int J Clin Pract Suppl 2004; 22-29, doi: 10.1111/j.1368-504X.2004.00376.x.
https://doi.org/10.1111/j.1368-504X.2004...
).
While PPARγ was initially regarded solely as a regulator of adipocyte differentiation and glucose homeostasis, it later became evident that it is also involved in cell differentiation, apoptosis and angiogenesis; biological processes that are deregulated in cancer. It is now established that PPARγ ligands, such as rosiglitazone, can inhibit the growth of tumor cells and produce antineoplastic effects in various cancers, including lung, breast, and colon cancer (44 Girnun GD, Chen L, Silvaggi J, Drapkin R, Chirieac LR, Padera RF, et al. Regression of drug-resistant lung cancer by the combination of rosiglitazone and carboplatin. Clin Cancer Res 2008; 14: 6478-6486, doi: 10.1158/1078-0432.CCR-08-1128.
https://doi.org/10.1158/1078-0432.CCR-08...
,55 Dai Y, Qiao L, Chan KW, Zou B, Ma J, Lan HY, et al. Loss of XIAP sensitizes rosiglitazone-induced growth inhibition of colon cancer in vivo. Int J Cancer 2008; 122: 2858-2863, doi: 10.1002/ijc.23443.
https://doi.org/10.1002/ijc.23443...
). One mechanism by which PPARγ exerts its effects is by increasing the expression of proteins involved in apoptosis and regulation of cell growth, such as Bax (BCL2-associated X), Bid (BH3 interacting-domain), tumor suppressor p53 (66 Gaetano C, Colussi C, Capogrossi MC. PEDF, PPAR-gamma, p53: deadly circuits arise when worlds collide. Cardiovasc Res 2007; 76: 195-196, doi: 10.1016/j.cardiores.2007.08.011.
https://doi.org/10.1016/j.cardiores.2007...
), and the cyclin-dependent kinase inhibitor (CDKI) p21 (77 Koga H, Sakisaka S, Harada M, Takagi T, Hanada S, Taniguchi E, et al. Involvement of p21(WAF1/Cip1), p27(Kip1), and p18(INK4c) in troglitazone-induced cell-cycle arrest in human hepatoma cell lines. Hepatology 2001; 33: 1087-1097, doi: 10.1053/jhep.2001.24024.
https://doi.org/10.1053/jhep.2001.24024...
). PPARγ ligands have been shown to suppress angiogenesis by inhibiting vascular endothelial growth factor (VEGF)-receptor 2 expression and blocking VEGF-receptor signaling (88 Xin X, Yang S, Kowalski J, Gerritsen ME. Peroxisome proliferator-activated receptor gamma ligands are potent inhibitors of angiogenesis in vitro and in vivo. J Biol Chem 1999; 274: 9116-9121, doi: 10.1074/jbc.274.13.9116.
https://doi.org/10.1074/jbc.274.13.9116...
). PPARγ agonists such as troglitazone and ciglitazone also inhibit metastasis by inhibiting migration of vascular endothelial cells (99 Goetze S, Bungenstock A, Czupalla C, Eilers F, Stawowy P, Kintscher U, et al. Leptin induces endothelial cell migration through Akt, which is inhibited by PPARgamma-ligands. Hypertension 2002; 40: 748-754, doi: 10.1161/01.HYP.0000035522.63647.D3.
https://doi.org/10.1161/01.HYP.000003552...
).
Since insulin-like growth factor binding protein-3 (IGFBP-3) also inhibits cancer cell growth independent of the IGF signaling system, it is now considered a tumor suppressor factor (1010 Baxter RC, Martin JL. Binding proteins for the insulin-like growth factors: structure, regulation and function. Prog Growth Factor Res 1989; 1: 49-68, doi: 10.1016/0955-2235(89)90041-0.
https://doi.org/10.1016/0955-2235(89)900...
,1111 Kelley KM, Oh Y, Gargosky SE, Gucev Z, Matsumoto T, Hwa V, et al. Insulin-like growth factor-binding proteins (IGFBPs) and their regulatory dynamics. Int J Biochem Cell Biol 1996; 28: 619-637, doi: 10.1016/1357-2725(96)00005-2.
https://doi.org/10.1016/1357-2725(96)000...
). The exact anticancer mechanism(s) of IGFBP-3 is not well understood; however, it has been reported that a number of factors, such as transforming growth factor (TGF)-β, retinoic acid, vitamin D, tumor necrosis factor (TNF), and p53, stimulate the expression and secretion of IGFBP-3, thus inhibiting cancer cell growth (1212 Colston KW, Perks CM, Xie SP, Holly JM. Growth inhibition of both MCF-7 and Hs578T human breast cancer cell lines by vitamin D analogues is associated with increased expression of insulin-like growth factor binding protein-3. J Mol Endocrinol 1998; 20: 157-162, doi: 10.1677/jme.0.0200157.
https://doi.org/10.1677/jme.0.0200157...
13 Rozen F, Zhang J, Pollak M. Antiproliferative action of tumor necrosis factor-alpha on MCF-7 breastcancer cells is associated with increased insulin-like growth factor binding protein-3 accumulation. Int J Oncol 1998; 13: 865-869.-1414 Buckbinder L, Talbott R, Velasco-Miguel S, Takenaka I, Faha B, Seizinger BR, et al. Induction of the growth inhibitor IGF-binding protein 3 by p53. Nature 1995; 377: 646-649, doi: 10.1038/377646a0.
https://doi.org/10.1038/377646a0...
). This study presents evidence that a mechanism whereby PPARγ exerts its anticancer effect is by increasing expression of the tumor suppressor gene IGFBP-3.
Material and Methods
Material
Cell culture reagents were purchased from Gibco-BRL Technologies (USA). Anti-PPARγ, IGFBP-3, p53, and secondary antibodies were purchased from Santa Cruz Biotechnology (USA). Poly(ADP-ribose) polymerase (PARP) antibody was purchased from Cell Signaling Technology (USA).
Cell line and cell culture
An established human gastric adenocarcinoma cell line (SNU-668), derived from the gastric cancer cells of a Korean patient, was purchased from the Korea Cell Line Bank (Korea). The cells were maintained in RPMI 1640 medium supplemented with 10% heat-inactivated fetal bovine serum (FBS), 300 µg/mL L-glutamine, 100 U/mL penicillin G and 100 µg/mL streptomycin sulfate. Cell cultures were incubated at 37°C in a 5% CO2 atmosphere. Cells from mid-log phase cultures were harvested, washed twice, and resuspended in culture medium prior to use. Cells that had been infected with adenovirus-expressing PPARγ (Ad/PPARγ), Ad/LacZ, or empty vector at a multiplicity of infection (MOI) of 50 or 100 were plated in 10% FBS-RPMI at a density of 2×l04 cells/well. After the indicated period of culture, trypan blue exclusion was used to assess cell viability by light microscopy. Cell viability was calculated by dividing the number of unstained viable cells count by the total number of cells.
RNA extraction and RT-PCR
Cells were treated with TRI reagent (Molecular Research Center, USA) and total RNA was extracted according to the manufacturer's instructions. RT-PCR and agarose gel electrophoresis were performed as previously described (1515 Kim MS, Kim SY, Arunachalam S, Hwang PH, Yi HK, Nam SY, et al. c-myb stimulates cell growth by regulation of insulin-like growth factor (IGF) and IGF-binding protein-3 in K562 leukemia cells. Biochem Biophys Res Commun 2009; 385: 38-43, doi: 10.1016/j.bbrc.2009.05.018.
https://doi.org/10.1016/j.bbrc.2009.05.0...
). Specific primer sets were designed from published cDNA sequences: sense PPARγ: 5′-TAA AGA GCC TGC GAA AGC-3′ and antisense PPARγ: 5′-TTG GCA GTG GCT CAG GAC-3′; sense actin: 5′-TGA CGG GGT CAC CCA CAC TGT GCC CAT CTA-3′ and antisense actin: 5′-CTA GAA GCA TTT GCG GTG GAC GAT GGA GGG-3′.
Preparation and administration of the recombinant adenoviral vector
The E1/E3-deleted replication-deficient recombinant adenovirus was made using the AdEasy system (Quantum Biotechnologies, Canada) as previously described (1515 Kim MS, Kim SY, Arunachalam S, Hwang PH, Yi HK, Nam SY, et al. c-myb stimulates cell growth by regulation of insulin-like growth factor (IGF) and IGF-binding protein-3 in K562 leukemia cells. Biochem Biophys Res Commun 2009; 385: 38-43, doi: 10.1016/j.bbrc.2009.05.018.
https://doi.org/10.1016/j.bbrc.2009.05.0...
). In brief, Kpn-XhoI restriction fragments from pcDNA3/PPARr and pcDNA3/p53 were ligated into Kpn-XhoI-digested pShuttle cytomegalovirus (CMV). Recombination into the pAdEasy viral backbone was accomplished in bacteria (recombination deficient Escherichia coli, strain BJ5183) according to the manufacturer's instructions. Recombinant adenoviruses were amplified in QBI-293A cells. Following amplification, lysates containing clonal recombinant adenovirus were purified by CsCl gradient centrifugation (100,000 g at 4°C for 20 h). Suspensions of recovered virus were aliquoted and stored at -20°C in 5 mM Tris buffer containing 50 mM NaCl, 0.05% bovine serum albumin, and 25% glycerol. Virus was titrated by serial dilution infection of QBI-293A cells, and plaques were counted under an overlay of 0.3% agarose, 10% fetal bovine serum, and 1% DMEM. For adenovirus infection, subconfluent cells were infected with adenovirus at a known MOI in culture medium supplemented with 2% FBS. After gentle and frequent shaking for 1 h at 37°C, cells were washed twice with phosphate-buffered saline (PBS) and fresh complete medium was added.
Luciferase assay
A 1.87-kb human IGFBP-3 promoter-luciferase reporter construct pGL3 (-1936/-64) and a series of deletion constructs including pGL3-Δ1100, pGL3-Δ1600, pGL3-Δ1755 and pGL3-Δ1795 were generated as previously described (1616 He TC, Zhou S, da Costa LT, Yu J, Kinzler KW, Vogelstein B. A simplified system for generating recombinant adenoviruses. Proc Natl Acad Sci U S A 1998; 95: 2509-2514, doi: 10.1073/pnas.95.5.2509.
https://doi.org/10.1073/pnas.95.5.2509...
). To generate pGL3-IGFBP-3, IGFBP-3 was digested with Bgl II, and the IGFBP-3 promoter fragment was cloned into Bgl II site of the pzGL-3 basic vector (Promega, USA). To generate pGL3-Δ1100, IGFBP-3 was double digested with Ssp I and Bgl II and the -1936/-64 region of the IGFBP-3 promoter was recombined into the pzGL-3 basic vector. SNU-668 cells were transfected for 24 h with IGFBP-3 promoter/pGL3 constructs using lipofectamine (Invitrogen, USA). The cells were washed with PBS, followed by infection with Ad/PPARγ in serum-free medium for 4 h. After 48 h, cell lysates were prepared and luciferase activity was measured using the Dual Luciferase kit (Promega) according to the manufacturer's instructions.
p53 siRNA transfection
p53 (sc-29435) siRNA was purchased from Santa Cruz Biotechnology. SNU-668 cells were transferred to 6-well plates (2×105/well), and allowed to grow overnight. p53 siRNA (200 pmol) and lipofectamine (5 µL) were diluted into OPTI-MEM (Invitrogen) to a total volume of 250 µL. After incubation at ambient temperature for 20 min and washing with serum-free RPMI medium, the siRNA/lipofectamine mix was added to the cells. Following incubation at 37°C for 6 h, the siRNA/lipofectamine mix was replaced with complete culture medium.
Crystal violet assay
Cells were transferred to 6-well plates (1×105) and infected with adenovirus vector as described above. After 3 days, cells were fixed, stained with 0.5% crystal violet, rinsed 3 times with deionized water, and allowed to air-dry.
Western blot analysis
Western blots were performed as previously described (1515 Kim MS, Kim SY, Arunachalam S, Hwang PH, Yi HK, Nam SY, et al. c-myb stimulates cell growth by regulation of insulin-like growth factor (IGF) and IGF-binding protein-3 in K562 leukemia cells. Biochem Biophys Res Commun 2009; 385: 38-43, doi: 10.1016/j.bbrc.2009.05.018.
https://doi.org/10.1016/j.bbrc.2009.05.0...
). In brief, cells were harvested 2-3 days after adenovirus infection. Proteins were extracted from transfected SNU-668 cells using lysis buffer and quantified by the Bradford dye-binding procedure (Bio-Rad, USA). Aliquots containing 20 µg protein were separated by 8% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) under denaturing conditions and transferred to Hybond-P membranes (Amersham, USA). After blocking with 5% skim milk in Tris-buffered saline and Tween 20 (TTBS), the membranes were incubated with primary antibody for 2 h. Protein signals were detected by enhanced chemiluminescence (NEN, USA).
Statistical analyses
Results are reported as means±SD. P<0.05 was considered to be statistically significant. Statistical significance between groups was assessed by the unpaired Student's t-test.
Results
Inhibition of cell growth and induction of apoptosis in SNU-668 cells by PPARγ
The anticancer effect of PPARγ was evaluated in SNU-668 cells infected with Ad/PPARγ (50 MOI). Robust expression of PPARγ was observed after 2 days (Figure 1A) and was coincident with marked inhibition of cell growth and reduction of cell numbers compared with the initial numbers (Figure 1B). The effect on cell growth was readily apparent 2 days after transfection, and even more prominent on days 3 and 4.

Effect of peroxisome proliferator activator receptor-gamma (PPARγ) overexpression on cell growth and apoptosis. A, Analysis of PPARγ expression (upper panel, RT-PCR) and PPARγ protein (lower panel, Western blot) in SNU-668 cells infected with adenovirus-expressing (Ad)/PPARγ. B, Inhibition of cell growth in SNU-668 cells infected with Ad/PPARγ. Upper panel, Cells were infected with Ad/PPARγ or Ad/LacZ at the indicated MOI for 4 h and then allowed to grow under standard culture conditions for a further 4 days. Cell number was determined by direct counting after trypan blue staining. Data are reported as means±SD of 3 independent experiments. *P<0.05 compared to control cells (Student's t-test). Lower panel, Crystal violet assay. Cell growth in mock-treated and Ad/LacZ-treated cells compared to cells treated with Ad/PPARγ. C, Effect of PPARγ overexpression on PARP, Bcl-2 and Bax in SNU-668 cells infected with Ad/PPARγ. Cell lysates were collected and processed at 48 and 72 h after infection, and total protein was analyzed by Western ligand blot with specific antibody.
We also assayed the levels of PARP protein, a caspase-3 substrate and key apoptosis-linked protease that is important in maintaining cell viability and DNA repair in response to environmental stress. Cleavage of PARP facilitates cellular disassembly and serves as a marker for cells undergoing apoptosis. We monitored enzyme activity using an antibody that binds to the 89-kDa fragment of the active enzyme. The active enzyme was present in Ad/PPARγ-transduced cells, but not in mock-transduced control or Ad/LacZ-transduced cells, indicating that PPARγ over-expression provides a stimulus for induction of apoptosis, and that the marked inhibitory effect on tumor cell growth by PPARγ is related to its ability to induce apoptosis. We also examined the effect of PPARγ on the expression of the anti-apoptotic protein Bcl-2, and the pro-apoptotic protein Bax, during induction of apoptosis in Ad/PPARγ-transduced cells. Western blot analysis showed decreased Bcl-2 protein and increased Bax protein expression in Ad/PPARγ-transduced cells compared with control cells (Figure 1C).
Relationship between IGFBP-3 expression and inhibition of cell growth by PPARγ
It has been shown that IGFBP-3 can inhibit cancer cell growth through pathways independent of the IGF signaling system (1010 Baxter RC, Martin JL. Binding proteins for the insulin-like growth factors: structure, regulation and function. Prog Growth Factor Res 1989; 1: 49-68, doi: 10.1016/0955-2235(89)90041-0.
https://doi.org/10.1016/0955-2235(89)900...
,1111 Kelley KM, Oh Y, Gargosky SE, Gucev Z, Matsumoto T, Hwa V, et al. Insulin-like growth factor-binding proteins (IGFBPs) and their regulatory dynamics. Int J Biochem Cell Biol 1996; 28: 619-637, doi: 10.1016/1357-2725(96)00005-2.
https://doi.org/10.1016/1357-2725(96)000...
). In addition to inhibiting cell growth and inducing apoptosis (Figure 2A and B), infection of SNU-668 cells with Ad/PPARγ induced a large increase in IGFBP-3 expression (Figure 2A), suggesting a relationship between IGFBP-3 expression and inhibition of cell growth by PPARγ.
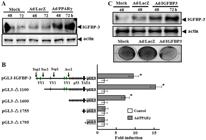
Effect of peroxisome proliferator activator receptor-gamma (PPARγ) on insulin-like growth factor-binding protein-3 (IGFBP-3) expression and IGFBP-3 promoter activity. A, Western blot analysis of IGFBP-3 protein in SNU-668 cells infected with adenovirus-expressing (Ad)/PPARγ. B, Analysis of PPARγ-induced IGFBP-3 promoter activation. SNU-668 cells were transiently transfected with a luciferase reporter vector containing the full-length 1.9-kb IGFBP-3 promoter fragment or deletion variants of the IGFBP-3 promoter. After 24 h, cells were treated with Ad/PPARγ in serum-free media for 4 h. Luciferase activity was measured in cell lysates 48 h after Ad/PPARγ infection. Data are reported as the means of fold-increase in luciferase activity of Ad/PPARγ transfected cells relative to control cells not receiving Ad/PPARγ. *P<0.05 compared to the Ad/LacZ-infected cells (Student's t-test). C, Effect of IGFBP-3 overexpression, induced by Ad/IGFBP-3 infection, on the growth of SNU-668 cells. Upper panel, Western blot showing IGFBP-3 protein levels in mock-treated cells, Ad/LacZ-treated cells and cells treated with Ad/IGFBP-3. Lower panel, Crystal violet assay. Cell growth in mock-treated and Ad/LacZ-treated cells compared to cells treated with Ad/IGFBP-3.
To investigate the molecular mechanisms underlying the effect of PPARγ on IGFBP-3 expression, we examined whether PPARγ directly affects transcription from the IGFBP-3 gene promoter. SNU-668 cells were transiently transfected with pGL3-IGFBP-3, a luciferase expression vector containing the full human IGFBP-3 pGL3 promoter construct (-1936/-64), promoter deletion constructs (pGL3-Δ1100, pGL3-Δ1600, pGL3-Δ1755 and pGL3-Δ1795), or the pGL3 vector alone before infection with Ad/PPARγ. Luciferase activity was robust in cells transfected with the full promoter sequence and even greater in cells transfected with the pGL3-Δ1100 deletion construct, which lacks 800 bp from the 5′ end of the full promoter sequence. In contrast, luciferase activity was minimal to absent in cells receiving the vector control (Figure 2B). These observations suggest that PPARγ induced the expression of IGFBP-3 through direct interaction with elements in the IGFBP-3 promoter.
Notably, cells transfected with the pGL3-Δ1600 construct exhibited robust luciferase activity compared to control cells (Figure 2B), but luciferase activity was absent in cells expressing the pGL3-Δ1755 or pGL3-Δ1795 constructs. The region deleted in pGL3-Δ1755 and pGL3-Δ1795 contains p53-binding sites. Thus, the critical PPARγ-responsive elements are located in the region containing p53-binding sites in the IGFBP-3 promoter.
We also evaluated whether IGFBP-3 itself exerts a direct effect on inhibition of SNU-668 cell growth. When the IGFBP-3 gene was overexpressed in SNU-668 cells using adenovirus (Ad/IGFBP-3), more significant cell growth retardation was observed in Ad/IGFBP-3 transduced SNU-668 cells than in control and Ad/LacZ-transduced cells (Figure 2C).
Induction of IGFBP-3 expression by PPARγ in cells in which p53 expression was inhibited
Having observed that sequences containing the p53-binding sites in the promoter region of IGFPB-3 were required for IGFBP-3 promoter activation by PPARγ, we next employed siRNA-mediated silencing of p53 to test whether the increased luciferase activity induced by PPARγ was dependent on p53 expression. SNU-668 cells were treated with p53 siRNA before induction of IGFBP-3 expression (by transient transfection) and PPARγ expression (by adenovirus infection). As anticipated, p53 siRNA reduced p53 expression in SNU-668 cells (Figure 3A). Furthermore, IGFBP-3 expression was strongly inhibited in cells lacking full p53 expression (Figure 3A). SNU-668 cells treated with p53 siRNA were transiently transfected for 24 h with the pGL3-1100 promoter-luciferase reporter plasmid, followed by infection with Ad/PPARγ in serum-free media for 4 h. Luciferase activity was compared in cells with or without p53 siRNA treatment. PPARγ-induced luciferase activity was increased over controls even in the absence of p53 siRNA treatment (Figure 3B).

Effect of p53 siRNA on insulin-like growth factor-binding protein-3 (IGFBP-3) expression and IGFBP-3 promoter activity. A, Western blot analysis of p53 and IGFBP-3 protein. The anti-actin blot indicates equal protein loading in each lane. B, Analysis of IGFBP-3 promoter activation by PPARγ. SNU-668 cells were treated with p53 siRNA for 6 h. The cells were then transiently transfected (24 h) with the pGL3-1100 promoter-luciferase reporter plasmid, followed by infection with adenovirus-expressing (Ad)/PPARγ in serum-free media. Controls were cells not treated with p53 siRNA and cells treated with Ad/LacZ. Data are reported as means±SD of 3 independent experiments. *P<0.05 compared to Ad/LacZ-infected cells without p53 siRNA (Student's t-test).
Inhibition of cell growth and IGFBP-3 expression by rosiglitazone, a PPARγ agonist
At concentrations of 1-10 μM, rosiglitazone, a known PPARγ agonist (44 Girnun GD, Chen L, Silvaggi J, Drapkin R, Chirieac LR, Padera RF, et al. Regression of drug-resistant lung cancer by the combination of rosiglitazone and carboplatin. Clin Cancer Res 2008; 14: 6478-6486, doi: 10.1158/1078-0432.CCR-08-1128.
https://doi.org/10.1158/1078-0432.CCR-08...
), induced growth inhibition of PPARγ-transfected SNU-668 cells (Figure 4A). The maximal effect was observed at a concentration of 5 μM. In PPARγ-transfected SNU-668 cells, treatment with 5 μM rosiglitazone increased luciferase activity 4-fold over untreated cells (Figure 4B), reflecting the known antineoplastic effects of the agonist. To verify the increased luciferase activity of the IGFBP-3 promoter in PPARγ-transfected SNU-668 cells following rosiglitazone treatment, we analyzed the expression of IGFBP-3 after rosiglitazone treatment. Rosiglitazone increased IGFBP-3 expression in PPARγ-transfected SNU-668 cells, which was well correlated with luciferase activity (Figure 4C).

Effect of the peroxisome proliferator activator receptor-gamma (PPARγ) agonist rosiglitazone (Rosi) on cell viability, IGFBP-3 promoter activity, IGFBP-3 expression. A, The effect of rosiglitazone on the viability of PPARγ transfected SNU-668 cells. Upper panel, PPARγ transfected cells were treated with various concentrations (1-10 μM) rosiglitazone and then allowed to grow under standard culture conditions for a further 4 days. The number of viable cells was determined by direct counting after trypan blue staining, and data are reported as the percent of the cell number before treatment. Lower panel, An equal number of cells were treated with increasing concentrations (1-10 μM) of rosiglitazone and incubated for 72 h. Cell viability was assessed using the crystal violet assay. B, Analysis of IGFBP-3 promoter activation by rosiglitazone. SNU-668 cells were transiently transfected with a luciferase reporter plasmid containing the full length IGFBP-3 promoter (pGL3-IGFBP-3), then infected with adenovirus-expressing (Ad)/PPARγ for 4 h. Twenty-four hours post transfection, luciferase activity was measured following treatment for an additional 24 h with rosiglitazone. C, Western blot analysis of GFBP-3 protein after rosiglitazone treatment in PPARγ transfected SNU-668 cells. The anti-actin blot indicates equal protein loading in each lane. Data are reported as means±SD of 3 independent experiments. *P<0.05 compared to the absence of rosiglitazone treatment (Student's t-test).
Discussion
The survival rate of patients with gastric cancer has increased dramatically since early detection and wide-range radical resection have been available. Nevertheless, gastric cancer is still a malignancy with high patient mortality. PPARγ plays an important role in controlling cellular metabolism (1717 Ahmadian M, Suh JM, Hah N, Liddle C, Atkins AR, Downes M, et al. PPARgamma signaling and metabolism: the good, the bad and the future. Nat Med 2013; 19: 557-566, doi: 10.1038/nm.3159.
https://doi.org/10.1038/nm.3159...
), and it also plays a role in inhibition of cell growth and induction of apoptosis in various types of cancer cells. Because of this, numerous studies have evaluated the therapeutic value of PPARγ in cancer. A common strategy for investigating novel approaches to treatment of gastric cancer has been to focus on interactions among the multiple factors known to be related to cancer progression (i.e., growth factors, oncogenes, tumor suppressor genes, and cell adhesion molecules) (1818 Walker GE, Wilson EM, Powell D, Oh Y. Butyrate, a histone deacetylase inhibitor, activates the human IGF binding protein-3 promoter in breast cancer cells: molecular mechanism involves an Sp1/Sp3 multiprotein complex. Endocrinology 2001; 142: 3817-3827, doi: 10.1210/endo.142.9.8380.
https://doi.org/10.1210/endo.142.9.8380...
). However, the interactions of PPARγ with other antitumor factors and their precise mechanisms, especially in gastric cancer, have not been thoroughly defined.
The expression of PPARγ in gastric cancer tissue was reported to be markedly reduced compared to its expression in normal tissue (1919 Lin J, Beerm DG. Molecular biology of upper gastrointestinal malignancies. Semin Oncol 2004; 31: 476-486, doi: 10.1053/j.seminoncol.2004.04.019.
https://doi.org/10.1053/j.seminoncol.200...
), and recent studies have indicated that apoptosis is induced following the administration of PPARγ ligands such as ciglitazone or 15-deoxy-12,14-prostaglandin J2 (15dPGJ2) in the colorectal carcinoma cell lines HT-29 and Colo-205 (2020 Yaacob NS, Darus HM, Norazmi MN. Modulation of cell growth and PPARgamma expression in human colorectal cancer cell lines by ciglitazone. Exp Toxicol Pathol 2008; 60: 505-512, doi: 10.1016/j.etp.2008.05.006.
https://doi.org/10.1016/j.etp.2008.05.00...
), and in MCG-803 human gastric cancer cells (2121 Chen YX, Zhong XY, Qin YF, Bing W, He LZ. 15d-PGJ2 inhibits cell growth and induces apoptosis of MCG-803 human gastric cancer cell line. World J Gastroenterol 2003; 9: 2149-2153.). Other recent studies have demonstrated that isorhamnetin, a 3′-O-methylated metabolite of quercetin (2222 Ramachandran L, Manu KA, Shanmugam MK, Li F, Siveen KS, Vali S, et al. Isorhamnetin inhibits proliferation and invasion and induces apoptosis through the modulation of peroxisome proliferator-activated receptor gamma activation pathway in gastric cancer. J Biol Chem 2012; 287: 38028-38040, doi: 10.1074/jbc.M112.388702.
https://doi.org/10.1074/jbc.M112.388702...
) and cannabinoids (2323 Vara D, Morell C, Rodriguez-Henche N, Diaz-Laviada I. Involvement of PPARgamma in the antitumoral action of cannabinoids on hepatocellular carcinoma. Cell Death Dis 2013; 4: e618, doi: 10.1038/cddis.2013.141.
https://doi.org/10.1038/cddis.2013.141...
), can mediate antitumor effects through modulation of the PPARγ activation pathway in gastric cancer and hepatocellular carcinoma. Therefore, we evaluated the effect of Ad/PPARγ infection in SNU-668 cells, a gastric adenocarcinoma cell line in which PPARγ is not normally expressed. PPARγ expression induced inhibition of cell growth and apoptosis in SNU-668 cells (Figure 1), strengthening the evidence that PPARγ inhibits tumor growth in gastric cancer cells via induction of apoptosis. Other studies have reported that PPARγ ligands such as 15dPGJ2 can induce apoptosis in MCG-803 gastric cancer cells (2121 Chen YX, Zhong XY, Qin YF, Bing W, He LZ. 15d-PGJ2 inhibits cell growth and induces apoptosis of MCG-803 human gastric cancer cell line. World J Gastroenterol 2003; 9: 2149-2153.), and this study shows that PPARγ expression, established in SNU-668 cells by Ad/PPARγ infection, can directly induce growth inhibition and apoptosis.
Because of its function as an antitumor agent independent of the IGF signaling pathway, IGFBP-3 is now recognized as a tumor suppressor factor (1010 Baxter RC, Martin JL. Binding proteins for the insulin-like growth factors: structure, regulation and function. Prog Growth Factor Res 1989; 1: 49-68, doi: 10.1016/0955-2235(89)90041-0.
https://doi.org/10.1016/0955-2235(89)900...
,1111 Kelley KM, Oh Y, Gargosky SE, Gucev Z, Matsumoto T, Hwa V, et al. Insulin-like growth factor-binding proteins (IGFBPs) and their regulatory dynamics. Int J Biochem Cell Biol 1996; 28: 619-637, doi: 10.1016/1357-2725(96)00005-2.
https://doi.org/10.1016/1357-2725(96)000...
). The antitumor activity of IGFBP-3 is related to its level of expression, and factors such as retinoic acid, TNF, p53 and butyric acid mediate IGFBP-3 expression through direct induction of its transcription (2424 Collard TJ, Guy M, Butt AJ, Perks CM, Holly JM, Paraskeva C, et al. Transcriptional upregulation of the insulin-like growth factor binding protein IGFBP-3 by sodium butyrate increases IGF-independent apoptosis in human colonic adenoma-derived epithelial cells. Carcinogenesis 2003; 24: 393-401, doi: 10.1093/carcin/24.3.393.
https://doi.org/10.1093/carcin/24.3.393...
). This study demonstrates that the response region of PPARγ exists within the promoter region of the IGFBP-3 gene and thus indicates that PPARγ can directly increase the expression of IGFBP-3 (Figure 2). We also observed inhibition of cell growth (Figure 4A) and increased activity from the IGFBP-3 promoter (Figure 4B) following administration of the PPARγ agonist rosiglitazone in PPARγ transfected SNU-668 cells. Although the DNA consensus sequence TGACCT (2525 Bonofiglio D, Aquila S, Catalano S, Gabriele S, Belmonte M, Middea E, et al. Peroxisome proliferator-activated receptor-gamma activates p53 gene promoter binding to the nuclear factor-kappaB sequence in human MCF7 breast cancer cells. Mol Endocrinol 2006; 20: 3083-3092, doi: 10.1210/me.2006-0192.
https://doi.org/10.1210/me.2006-0192...
), required for PPARs binding in promoter regions, does not exist within the IGFBP-3 promoter, imperfect motifs, e.g. TGACCn, exist at -190, -260, -270 bases upstream from the TATA box. Our data suggest that the imperfect TGACCn motifs are involved in the transcriptional activation of IGFBP-3 by PPARγ. The IGFBP-3 promoter contains a cluster of eleven p53 consensus binding sites (1414 Buckbinder L, Talbott R, Velasco-Miguel S, Takenaka I, Faha B, Seizinger BR, et al. Induction of the growth inhibitor IGF-binding protein 3 by p53. Nature 1995; 377: 646-649, doi: 10.1038/377646a0.
https://doi.org/10.1038/377646a0...
), and the imperfect PPARγ binding motifs (TGACCn) are coincident with part of this cluster. Our analysis of five sequential deletion mutants of the IGFBP-3 promoter indicates a significant loss of IGFBP-3 promoter activity in cells transfected with deletion constructs lacking putative p53-binding sites (i.e., pGL3-Δ1755, pGL3-Δ1795; Figure 2B).
When activated, p53 stimulates the expression of a broad range of genes, including IGFBP-3 (2424 Collard TJ, Guy M, Butt AJ, Perks CM, Holly JM, Paraskeva C, et al. Transcriptional upregulation of the insulin-like growth factor binding protein IGFBP-3 by sodium butyrate increases IGF-independent apoptosis in human colonic adenoma-derived epithelial cells. Carcinogenesis 2003; 24: 393-401, doi: 10.1093/carcin/24.3.393.
https://doi.org/10.1093/carcin/24.3.393...
). A recent study suggests that PPARγ agonists promote growth arrest and apoptosis in breast cancer cells, at least in part through cross-talk between p53 and PPARγ (2626 Lee DY, Yi HK, Hwang PH, Oh Y. Enhanced expression of insulin-like growth factor binding protein-3 sensitizes the growth inhibitory effect of anticancer drugs in gastric cancer cells. Biochem Biophys Res Commun 2002; 294: 480-486, doi: 10.1016/S0006-291X(02)00491-6.
https://doi.org/10.1016/S0006-291X(02)00...
). To determine whether p53 is required for the transcriptional activation of IGFBP-3 by PPARγ, we examined PPARγ-induced luciferase activity in pGL3-Δ1100-transfected cells pretreated with siRNA (Figure 3B). The PPARγ-induced luciferase activity was strong even when p53 expression was markedly reduced by siRNA, suggesting that the effect of PPARγ on luciferase activity is likely due to direct interaction of PPARγ on the IGFBP promoter, rather than to indirect regulation by p53. We previously reported that the exogenous addition of IGFBP-3 inhibited the growth of cells not expressing IGFBP-3, causing them to undergo apoptosis (2727 Lecka-Czernik B, Acckert-Bicknell C, Adamo ML, Marmolejos V, Churchill GA, Shockley KR, et al. Activation of peroxisome proliferator-activated receptor gamma (PPARgamma) by rosiglitazone suppresses components of the insulin-like growth factor regulatory system in vitro and in vivo. Endocrinology 2007; 148: 903-911, doi: 10.1210/en.2006-1121.
https://doi.org/10.1210/en.2006-1121...
). Our demonstration of induction of apoptosis by PPARγ in SNU-668 cells (Figure 1B) is in accordance with that observation.
In summary, our data indicated that PPARγ can act as a transcription factor, and that one mechanism whereby it exerts an antitumor effect in gastric cancer cells is by increasing expression of the tumor suppressor gene IGFBP-3. There have been few studies of the interaction of PPARγ with IGFs (2828 Higashi Y, Holder K, Delafontaine P. Thiazolidiones up-regulate insulin-like growth factor-1 receptor via a peroxisome proliferator-activated receptor gamma independent pathway. J Biol Chem 2010; 285: 36361-36368, doi: 10.1074/jbc.M110.137661.
https://doi.org/10.1074/jbc.M110.137661...
), and there are no specific data on IGFBP-3. Additional data demonstrating a role for PPARγ in increasing the anticancer potential of IGFBP-3 may provide new approaches for treatment of gastric cancers.
Acknowledgments
The authors thank Dr. Zena K. Indik (Department of Medicine, University of Pennsylvania, Philadelphia, PA, USA) and Professor Kannan (Division of Biotechnology, Advanced Institute of Biosciences, Chonbuk National University, Jeonju, Korea) for their careful review of the manuscript and comments.
References
-
1Robinson-Rechavi M, Carpentier AS, Duffraisse M, Laudet V. How many nuclear hormone receptors are there in the human genome? Trends Genet 2001; 17: 554-556, doi: 10.1016/S0168-9525(01)02417-9.
» https://doi.org/10.1016/S0168-9525(01)02417-9 -
2Schoonjans K, Staels B, Auwerx J. The peroxisome proliferator activated receptors (PPARS) and their effects on lipid metabolism and adipocyte differentiation. Biochim Biophys Acta 1996; 1302: 93-109, doi: 10.1016/0005-2760(96)00066-5.
» https://doi.org/10.1016/0005-2760(96)00066-5 -
3Gervois P, Fruchart JC, Staels B. Inflammation, dyslipidaemia, diabetes and PPars: pharmacological interest of dual PPARalpha and PPARgamma agonists. Int J Clin Pract Suppl 2004; 22-29, doi: 10.1111/j.1368-504X.2004.00376.x.
» https://doi.org/10.1111/j.1368-504X.2004.00376.x -
4Girnun GD, Chen L, Silvaggi J, Drapkin R, Chirieac LR, Padera RF, et al. Regression of drug-resistant lung cancer by the combination of rosiglitazone and carboplatin. Clin Cancer Res 2008; 14: 6478-6486, doi: 10.1158/1078-0432.CCR-08-1128.
» https://doi.org/10.1158/1078-0432.CCR-08-1128 -
5Dai Y, Qiao L, Chan KW, Zou B, Ma J, Lan HY, et al. Loss of XIAP sensitizes rosiglitazone-induced growth inhibition of colon cancer in vivo. Int J Cancer 2008; 122: 2858-2863, doi: 10.1002/ijc.23443.
» https://doi.org/10.1002/ijc.23443 -
6Gaetano C, Colussi C, Capogrossi MC. PEDF, PPAR-gamma, p53: deadly circuits arise when worlds collide. Cardiovasc Res 2007; 76: 195-196, doi: 10.1016/j.cardiores.2007.08.011.
» https://doi.org/10.1016/j.cardiores.2007.08.011 -
7Koga H, Sakisaka S, Harada M, Takagi T, Hanada S, Taniguchi E, et al. Involvement of p21(WAF1/Cip1), p27(Kip1), and p18(INK4c) in troglitazone-induced cell-cycle arrest in human hepatoma cell lines. Hepatology 2001; 33: 1087-1097, doi: 10.1053/jhep.2001.24024.
» https://doi.org/10.1053/jhep.2001.24024 -
8Xin X, Yang S, Kowalski J, Gerritsen ME. Peroxisome proliferator-activated receptor gamma ligands are potent inhibitors of angiogenesis in vitro and in vivo J Biol Chem 1999; 274: 9116-9121, doi: 10.1074/jbc.274.13.9116.
» https://doi.org/10.1074/jbc.274.13.9116 -
9Goetze S, Bungenstock A, Czupalla C, Eilers F, Stawowy P, Kintscher U, et al. Leptin induces endothelial cell migration through Akt, which is inhibited by PPARgamma-ligands. Hypertension 2002; 40: 748-754, doi: 10.1161/01.HYP.0000035522.63647.D3.
» https://doi.org/10.1161/01.HYP.0000035522.63647.D3 -
10Baxter RC, Martin JL. Binding proteins for the insulin-like growth factors: structure, regulation and function. Prog Growth Factor Res 1989; 1: 49-68, doi: 10.1016/0955-2235(89)90041-0.
» https://doi.org/10.1016/0955-2235(89)90041-0 -
11Kelley KM, Oh Y, Gargosky SE, Gucev Z, Matsumoto T, Hwa V, et al. Insulin-like growth factor-binding proteins (IGFBPs) and their regulatory dynamics. Int J Biochem Cell Biol 1996; 28: 619-637, doi: 10.1016/1357-2725(96)00005-2.
» https://doi.org/10.1016/1357-2725(96)00005-2 -
12Colston KW, Perks CM, Xie SP, Holly JM. Growth inhibition of both MCF-7 and Hs578T human breast cancer cell lines by vitamin D analogues is associated with increased expression of insulin-like growth factor binding protein-3. J Mol Endocrinol 1998; 20: 157-162, doi: 10.1677/jme.0.0200157.
» https://doi.org/10.1677/jme.0.0200157 -
13Rozen F, Zhang J, Pollak M. Antiproliferative action of tumor necrosis factor-alpha on MCF-7 breastcancer cells is associated with increased insulin-like growth factor binding protein-3 accumulation. Int J Oncol 1998; 13: 865-869.
-
14Buckbinder L, Talbott R, Velasco-Miguel S, Takenaka I, Faha B, Seizinger BR, et al. Induction of the growth inhibitor IGF-binding protein 3 by p53. Nature 1995; 377: 646-649, doi: 10.1038/377646a0.
» https://doi.org/10.1038/377646a0 -
15Kim MS, Kim SY, Arunachalam S, Hwang PH, Yi HK, Nam SY, et al. c-myb stimulates cell growth by regulation of insulin-like growth factor (IGF) and IGF-binding protein-3 in K562 leukemia cells. Biochem Biophys Res Commun 2009; 385: 38-43, doi: 10.1016/j.bbrc.2009.05.018.
» https://doi.org/10.1016/j.bbrc.2009.05.018 -
16He TC, Zhou S, da Costa LT, Yu J, Kinzler KW, Vogelstein B. A simplified system for generating recombinant adenoviruses. Proc Natl Acad Sci U S A 1998; 95: 2509-2514, doi: 10.1073/pnas.95.5.2509.
» https://doi.org/10.1073/pnas.95.5.2509 -
17Ahmadian M, Suh JM, Hah N, Liddle C, Atkins AR, Downes M, et al. PPARgamma signaling and metabolism: the good, the bad and the future. Nat Med 2013; 19: 557-566, doi: 10.1038/nm.3159.
» https://doi.org/10.1038/nm.3159 -
18Walker GE, Wilson EM, Powell D, Oh Y. Butyrate, a histone deacetylase inhibitor, activates the human IGF binding protein-3 promoter in breast cancer cells: molecular mechanism involves an Sp1/Sp3 multiprotein complex. Endocrinology 2001; 142: 3817-3827, doi: 10.1210/endo.142.9.8380.
» https://doi.org/10.1210/endo.142.9.8380 -
19Lin J, Beerm DG. Molecular biology of upper gastrointestinal malignancies. Semin Oncol 2004; 31: 476-486, doi: 10.1053/j.seminoncol.2004.04.019.
» https://doi.org/10.1053/j.seminoncol.2004.04.019 -
20Yaacob NS, Darus HM, Norazmi MN. Modulation of cell growth and PPARgamma expression in human colorectal cancer cell lines by ciglitazone. Exp Toxicol Pathol 2008; 60: 505-512, doi: 10.1016/j.etp.2008.05.006.
» https://doi.org/10.1016/j.etp.2008.05.006 -
21Chen YX, Zhong XY, Qin YF, Bing W, He LZ. 15d-PGJ2 inhibits cell growth and induces apoptosis of MCG-803 human gastric cancer cell line. World J Gastroenterol 2003; 9: 2149-2153.
-
22Ramachandran L, Manu KA, Shanmugam MK, Li F, Siveen KS, Vali S, et al. Isorhamnetin inhibits proliferation and invasion and induces apoptosis through the modulation of peroxisome proliferator-activated receptor gamma activation pathway in gastric cancer. J Biol Chem 2012; 287: 38028-38040, doi: 10.1074/jbc.M112.388702.
» https://doi.org/10.1074/jbc.M112.388702 -
23Vara D, Morell C, Rodriguez-Henche N, Diaz-Laviada I. Involvement of PPARgamma in the antitumoral action of cannabinoids on hepatocellular carcinoma. Cell Death Dis 2013; 4: e618, doi: 10.1038/cddis.2013.141.
» https://doi.org/10.1038/cddis.2013.141 -
24Collard TJ, Guy M, Butt AJ, Perks CM, Holly JM, Paraskeva C, et al. Transcriptional upregulation of the insulin-like growth factor binding protein IGFBP-3 by sodium butyrate increases IGF-independent apoptosis in human colonic adenoma-derived epithelial cells. Carcinogenesis 2003; 24: 393-401, doi: 10.1093/carcin/24.3.393.
» https://doi.org/10.1093/carcin/24.3.393 -
25Bonofiglio D, Aquila S, Catalano S, Gabriele S, Belmonte M, Middea E, et al. Peroxisome proliferator-activated receptor-gamma activates p53 gene promoter binding to the nuclear factor-kappaB sequence in human MCF7 breast cancer cells. Mol Endocrinol 2006; 20: 3083-3092, doi: 10.1210/me.2006-0192.
» https://doi.org/10.1210/me.2006-0192 -
26Lee DY, Yi HK, Hwang PH, Oh Y. Enhanced expression of insulin-like growth factor binding protein-3 sensitizes the growth inhibitory effect of anticancer drugs in gastric cancer cells. Biochem Biophys Res Commun 2002; 294: 480-486, doi: 10.1016/S0006-291X(02)00491-6.
» https://doi.org/10.1016/S0006-291X(02)00491-6 -
27Lecka-Czernik B, Acckert-Bicknell C, Adamo ML, Marmolejos V, Churchill GA, Shockley KR, et al. Activation of peroxisome proliferator-activated receptor gamma (PPARgamma) by rosiglitazone suppresses components of the insulin-like growth factor regulatory system in vitro and in vivo Endocrinology 2007; 148: 903-911, doi: 10.1210/en.2006-1121.
» https://doi.org/10.1210/en.2006-1121 -
28Higashi Y, Holder K, Delafontaine P. Thiazolidiones up-regulate insulin-like growth factor-1 receptor via a peroxisome proliferator-activated receptor gamma independent pathway. J Biol Chem 2010; 285: 36361-36368, doi: 10.1074/jbc.M110.137661.
» https://doi.org/10.1074/jbc.M110.137661
-
First published online.
Publication Dates
-
Publication in this collection
13 Jan 2015 -
Date of issue
Mar 2015
History
-
Received
28 June 2014 -
Accepted
13 Oct 2014




