Abstract
Low-level lasers are used at low power densities and doses according to clinical protocols supplied with laser devices or based on professional practice. Although use of these lasers is increasing in many countries, the molecular mechanisms involved in effects of low-level lasers, mainly on DNA, are controversial. In this study, we evaluated the effects of low-level red lasers on survival, filamentation, and morphology of Escherichia colicells that were exposed to ultraviolet C (UVC) radiation. Exponential and stationary wild-type and uvrA-deficient E. coli cells were exposed to a low-level red laser and in sequence to UVC radiation. Bacterial survival was evaluated to determine the laser protection factor (ratio between the number of viable cells after exposure to the red laser and UVC and the number of viable cells after exposure to UVC). Bacterial filaments were counted to obtain the percentage of filamentation. Area-perimeter ratios were calculated for evaluation of cellular morphology. Experiments were carried out in duplicate and the results are reported as the means of three independent assays. Pre-exposure to a red laser protected wild-type and uvrA-deficient E. coli cells against the lethal effect of UVC radiation, and increased the percentage of filamentation and the area-perimeter ratio, depending on UVC fluence and physiological conditions in the cells. Therapeutic, low-level red laser radiation can induce DNA lesions at a sub-lethal level. Consequences to cells and tissues should be considered when clinical protocols based on this laser are carried out.
DNA; Escherichia coli; Laser; Ultraviolet radiation
Introduction
Laser devices are monochromatic, collimated, and coherent radiation sources. These devices have been used with different purposes for treatment of many diseases in soft and bone tissues at varying power densities, doses, and wavelengths (11. Niemz MH. Laser-tissue interactions: Fundamentals and applications. New York: Springer-Verlag; 2007.). Low-level laser therapies are used at low power densities and doses, in the so-called therapeutic window (600-1100 nm), in pre-established clinical protocols supplied with laser devices or based on professional practice.
Although use of low-level laser therapy is increasing in many countries, there are questions regarding the molecular mechanisms involved in the effects of this therapy. Molecular targets (chromophores) appear to be some mitochondrial cytochromes and porphyrins in the cytoplasm (22. Kohli R, Gupta PK. Irradiance dependence of the He-Ne laser-induced protection against UVC radiation in E. colistrains. J Photochem Photobiol B 2003; 69: 161-167, doi: 10.1016/S1011-1344(03)00018-6.
https://doi.org/10.1016/S1011-1344(03)00...
). Laser radiation energy is absorbed by chromophores and subsequent intracellular transducers are responsible for transforming the laser radiation energy into a cellular signal (33. Eells JT, Wong-Riley MT, VerHoeve J, Henry M, Buchman EV, Kane MP, et al. Mitochondrial signal transduction in accelerated wound and retinal healing by near-infrared light therapy. Mitochondrion 2004; 4: 559-567, doi: 10.1016/j.mito.2004.07.033.
https://doi.org/10.1016/j.mito.2004.07.0...
). A cascade of molecular effects occurs as a consequence of amplification of the photosignal, including an increase in nucleic acids (44. Karu TI, Kalendo GS, Letokhov VS, Lobko VV. Biostimulation of HeLa cells by low intensity visible light. III. Stimulation of nucleic acid synthesis in plateau phase cells. Il Nuovo Cimento D 1984; 3: 319-325, doi: 10.1007/BF02457461.
https://doi.org/10.1007/BF02457461....
) and ATP (55. Karu T, Pyatibrat L, Kalendo G. Irradiation with He-Ne laser increases ATP level in cells cultivated in vitro. J Photochem Photobiol B 1995; 27: 219-223, doi: 10.1016/1011-1344(94)07078-3.
https://doi.org/10.1016/1011-1344(94)070...
), as well as gene transcription (66. Zhang Y, Song S, Fong CC, Tsang CH, Yang Z, Yang M. cDNA microarray analysis of gene expression profiles in human fibroblast cells irradiated with red light. J Invest Dermatol 2003; 120: 849-857, doi: 10.1046/j.1523-1747.2003.12133.x.
https://doi.org/10.1046/j.1523-1747.2003...
). These alterations increase metabolism, protein secretion, and cellular division after low-level laser exposure (55. Karu T, Pyatibrat L, Kalendo G. Irradiation with He-Ne laser increases ATP level in cells cultivated in vitro. J Photochem Photobiol B 1995; 27: 219-223, doi: 10.1016/1011-1344(94)07078-3.
https://doi.org/10.1016/1011-1344(94)070...
). The entire effect (biostimulation or biomodulation) is considered the basis of therapeutic applications, such as wound healing (77. Avci P, Gupta A, Sadasivam M, Vecchio D, Pam Z, Pam N, et al. Low-level laser (light) therapy (LLLT) in skin: stimulating, healing, restoring. Semin Cutan Med Surg 2013; 32: 41-52.). For other applications, such as pain relief and herpes simplex treatment, the molecular mechanisms are not understood.
Few studies have evaluated the effects of low-level laser radiation on DNA and the possible consequences to cells and tissues, and whether this radiation induces molecular damage. In fact, low-level lasers at therapeutic doses can induce free radical generation (88. Kim YG. Laser mediated production of reactive oxygen and nitrogen species; implications for therapy. Free Radic Res 2002; 36: 1243-1250, doi: 10.1080/1071576021000028389.
https://doi.org/10.1080/1071576021000028...
) and sub-lethal DNA lesions (99. Fonseca AS, Moreira TO, Paixao DL, Farias FM, Guimaraes OR, de Paoli S, et al. Effect of laser therapy on DNA damage. Lasers Surg Med 2010; 42: 481-488, doi: 10.1002/lsm.v42:6.
https://doi.org/10.1002/lsm.v42:6....
). Additionally, these lasers induce different SOS responses in cells deficient in DNA repair mechanisms (22. Kohli R, Gupta PK. Irradiance dependence of the He-Ne laser-induced protection against UVC radiation in E. colistrains. J Photochem Photobiol B 2003; 69: 161-167, doi: 10.1016/S1011-1344(03)00018-6.
https://doi.org/10.1016/S1011-1344(03)00...
,1010. Fonseca AS, Presta GA, Geller M, Paoli F. Low intensity infrared laser induces filamentation in Escherichia coli cells. Lasers Phys 2011; 21: 1-9, doi: 10.1134/S1054660X11170051.
https://doi.org/10.1134/S1054660X1117005...
).
Ultraviolet radiation absorption by DNA molecules results in pyrimidine dimers as the main direct DNA lesion, while absorption by other molecules causes free radical generation. These chemical species induce different types of lesions in DNA, mainly with nitrogen bases, by oxidizing chemical reactions (1111. Koch WH, Woodgate R. The SOS response. In: Nickoloff JA, Hoekstra MF (Editors), DNA damage and repair. Vol I: DNA repair in procaryotes and lower eukaryotes. New Jersey: Humana Press; 1998. p 107-134.). Pyrimidine dimers and oxidizing DNA lesions induced by ultraviolet radiation are repaired by the nucleotide excision repair pathway (1111. Koch WH, Woodgate R. The SOS response. In: Nickoloff JA, Hoekstra MF (Editors), DNA damage and repair. Vol I: DNA repair in procaryotes and lower eukaryotes. New Jersey: Humana Press; 1998. p 107-134.). Cells that are deficient in nucleotide excision repair fail to remove pyrimidine dimers and other bulky lesions caused by ultraviolet radiation exposure, similar to humans presenting with xeroderma pigmentosum. Escherichia coli has three proteins (uvrA, uvrB, and uvrC) involved in recognizing the lesion and incision endonuclease function (1111. Koch WH, Woodgate R. The SOS response. In: Nickoloff JA, Hoekstra MF (Editors), DNA damage and repair. Vol I: DNA repair in procaryotes and lower eukaryotes. New Jersey: Humana Press; 1998. p 107-134.). E. coli cells that are deficient in these proteins are used as experimental models to evaluate cellular responses to ultraviolet radiation (1111. Koch WH, Woodgate R. The SOS response. In: Nickoloff JA, Hoekstra MF (Editors), DNA damage and repair. Vol I: DNA repair in procaryotes and lower eukaryotes. New Jersey: Humana Press; 1998. p 107-134.). However, previous studies have shown that cells exposed to increased free radical concentrations are more resistant to ultraviolet radiation (1212. Butts BD, Kwei KA, Bowden GT, Briehl MM. Elevated basal reactive oxygen species and phospho-Akt in murine keratinocytes resistant to ultraviolet B-induced apoptosis. Mol Carcinog 2003; 37: 149-157.). Moreover, previous results in our laboratory have shown that a low-level red laser induces resistance to hydrogen peroxide (99. Fonseca AS, Moreira TO, Paixao DL, Farias FM, Guimaraes OR, de Paoli S, et al. Effect of laser therapy on DNA damage. Lasers Surg Med 2010; 42: 481-488, doi: 10.1002/lsm.v42:6.
https://doi.org/10.1002/lsm.v42:6....
) and induces filamentation in E. coli cells deficient in repair of oxidative DNA lesions (1010. Fonseca AS, Presta GA, Geller M, Paoli F. Low intensity infrared laser induces filamentation in Escherichia coli cells. Lasers Phys 2011; 21: 1-9, doi: 10.1134/S1054660X11170051.
https://doi.org/10.1134/S1054660X1117005...
).
Therefore, the effects of low-level lasers on DNA molecules by oxidative mechanisms are still controversial. This study evaluated the effects of a low-level red laser on survival, filamentation, and morphology of E. coli cells that were deficient in nucleotide excision repair and were exposed to ultraviolet C (UVC) radiation.
Material and Methods
Low-level red laser and UVC source
A therapeutic, low-level red laser (AlGaInP, 10 mW), with emission at 658 nm, was purchased from HTM Eletrônica (Brazil). UVC radiation was produced from a germicidal lamp (Philips, The Netherlands). Table 1 shows parameters of the laser.
Evaluation of low-level red laser exposure on survival of E. coli cells with UVC radiation
Cultures of E. coli AB1157 (wild-type) and AB1886 (uvrA-deficient) in the stationary and exponential growth phases were exposed to a low-level red laser (8 J/cm2) and UVC radiation (25, 50, and 100 mJ/cm2). The rate of survival was evaluated. The laser device controlled laser fluence and irradiation time. UVC fluence was measured by an ultraviolet radiometer (Instrutherm, Brazil) and irradiation times were 25, 50, and 100 s. The laser device was positioned so that the laser beam covered almost all of the surface of bacterial suspension aliquots. Aliquots of bacteria from frozen stocks were used and further incubated in nutritive medium to reach exponential growth (108cells/mL, 2-3 h, 37°C). Other experiments were carried out with cultures of the same E. coli strains in the stationary growth phase (1010 cells/mL, 18 h, 37°C). Bacterial cells were centrifuged (700 g, 15 min) and suspended twice in saline (0.9% NaCl). Aliquots (50 µL, n=5, for each fluence) of bacterial suspensions were then exposed to the low-level red laser and UVC. Bacterial suspensions that were not exposed to the laser or UVC were used as controls. Bacterial suspensions were spread onto Petri dishes. Colonies that formed after overnight incubation at 37°C were counted. The survival fraction was then calculated, and the laser protection factor was calculated by the ratio between the number of viable cells after exposure to the red laser and UVC and the number of viable cells after exposure to UVC.
Bacterial filamentation assay
To evaluate induction of filamentation, exponential and stationary E. coli AB1157 and AB1886 cultures were obtained and exposed to a low-level red laser and UVC as described above. Bacterial suspensions that were not exposed to a laser or ultraviolet radiation were used as controls. Immediately after exposure, aliquots (20 µL) were withdrawn, spread onto microscopic slides, and stained by the Gram method (1313. Cappuccino JG, Sherman N. Microbiology: a laboratory manual. California: Benjamin Cummings Science Publishing; 1999.). Bacterial cells (100 cells per field, three fields per slide, two slides per group) were visualized by a Carl Zeiss microscope (Germany) equipped with an A-plan 40× objective, a 0.90 condenser, and a 100-W halogen lamp. The images were captured with an AxioCam HRc Sony 12M color microscopy camera, using Axiovision software (Carl Zeiss). The images were then analyzed by Image Proplus software (version 6.0 for Windows XP, Microsoft Corporation, USA) to determine the percentage of bacterial filamentation. A bacterial filament was considered as 2.5 times the average area of the bacterial cells. Experiments were carried out in duplicate and the results are reported as the means of three independent assays.
Statistical analysis
Data are reported as means±SD of the protection factor, percentage of filamentation, and area-perimeter ratio. One-way analysis of variance followed by Tukey’s post hoc test were performed to determine statistical differences, with P<0.05 as the least significant level.
Results
Survival of E. coli cells exposed to low-level red laser and UVC radiation
Table 2 shows the protection factors for low-level red laser radiation on E. coli AB1157 cells, which were exposed to different UVC radiation levels, in the exponential growth phase. There was no significant (P&0.05) protection of the low-level red laser onE. coli AB1157 cultures against the lethal effect of UVC radiation.
To determine whether the growth phase interferes with laser-induced biological effects, E. coli cultures in the stationary growth phase were exposed to UVC radiation after red laser exposure. Table 2 shows the protection factors of the red laser onE. coli AB1157 cultures that were exposed to UVC in the stationary growth phase under the same conditions used to irradiate cultures of this strain in the exponential growth phase. In contrast to the exponential growth phase, low-level red laser radiation significantly protected stationaryE. coli AB1157 cultures against the lethal effect of UVC radiation at lowest fluence levels (P<0.05 vs controls). However, at the highest fluence level (100 J/cm2), exposure to the red laser decreased the protection factor significantly (P<0.05), compared with E. coli cultures exposed to UVC (controls).
Pre-exposure to low-level red laser radiation was evaluated in E. coli AB1886 cultures that were exposed to UVC radiation (Table 2). In contrast to wild-typeE. coli (AB1157), laser pre-exposure (8 J/cm2) induced significant (P<0.05) protection against the lethal effect of UVC radiation on E. coli AB1886 at the highest fluences used (50 and 100 mJ/cm2).
Laser-induced protection of the lethal effect of UVC was evaluated in stationaryE. coli AB1886 cultures (Table 2). In this condition, the red laser did not significantly protect E. coli AB1886 cells against the lethal effect of UVC radiation (P&0.05).
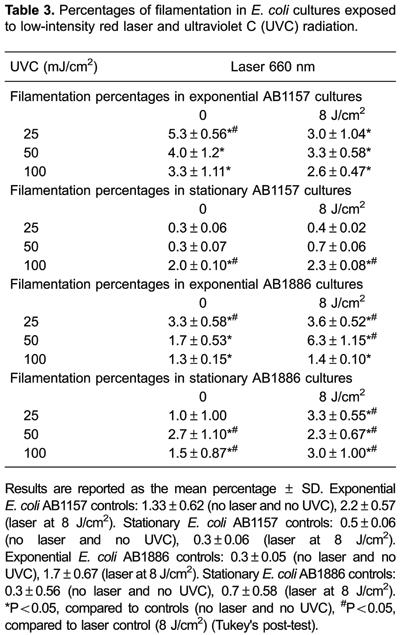
Induction of filamentation in E. coli cells exposed to low-level red laser and UVC radiation
Induction of filamentation was evaluated in exponential E. coliAB1157 cultures pre-exposed to low-level red laser radiation and exposed to UVC radiation (Table 3). Exposure to UVC radiation significantly increased the percentage of bacterial filamentation (P<0.05), compared with E. coli cultures exposed to UVC (controls). Similar percentages of bacterial filamentation were observed in cultures pre-exposed to the red laser (8 J/cm2). Percentages of bacterial filamentation were not significantly different from those inE. coli AB1157 cultures exposed to laser alone (P&0.05), except in cultures exposed to UVC radiation at the lowest fluence (25 mJ/cm2).
The percentage of filamentation in the stationary growth phase of E. coli AB1157 cultures did not significantly change after UVC exposure at the lowest fluences (25 and 50 mJ/cm2) (P&0.05, Table 3). However, the percentage of bacterial filamentation was significantly higher at the highest fluence (100 mJ/cm2) compared with controls (no laser and no UVC) and compared with the laser alone (P<0.05).
The percentage of bacterial filamentation in exponential E. coliAB1886 cultures that were exposed to UVC radiation is shown in Table 3. Exposure to UVC radiation significantly induced filamentation in non-pre-exposed and pre-exposed low-level red laser radiation (P<0.05). These percentages of bacterial filamentation were similar to those induced by the red laser alone (P&0.05).
Except for the lowest UVC fluence (25 mJ/cm2), non-laser pre-exposed and laser pre-exposed stationary E. coli AB1886 cultures had significantly higher percentages of bacterial filamentation (P<0.05), compared with E. coli cultures exposed to UVC (controls). These percentages of bacterial filamentation were significantly higher than those observed in stationary E. coli AB1886 cultures that were exposed to the red laser alone, except for UVC alone at the lowest fluence (P<0.05, Table 3).
Morphology of E. coli cells exposed to low-level red laser and UVC radiation
Figure 1 shows representative cells fromE. coli AB1157 cultures in the exponential growth phase (1A) and analysis of bacterial cells (1B). The area-perimeter ratio ofE. coli AB1157 cells in the exponential growth phase, exposed to UVC after exposure to low-level red laser radiation, was not significantly (P&0.05) altered (data not shown). Similarly, red laser and UVC radiation alone, or red laser followed by UVC radiation, did not significantly (P&0.05) alter the area-perimeter ratio of E. coli cells in the stationary growth phase.
Representative image of bacterial filamentation in an AB1157 culture in the exponential growth phase. A, The arrow indicates bacterial filamentation; B, inset shows how image analysis was performed. A bacterial filament was considered present when the area of a bacterial cell was larger than 2.5 times the mean area of bacterial cells.
Effects of low-level red laser and UVC radiation on the area-perimeter ratio of exponential E. coli AB1886 cells were also examined (Figure 2). Exposure to UVC radiation after pre-exposure to the red laser significantly increased the area-perimeter ratio, at least at the lowest UVC fluences (25 and 50 mJ/cm2, P<0.05). However, this effect was not observed in stationary E. coliAB1886 cells because the area-perimeter ratio was not significantly (P&0.05) altered (data not shown).
Area-perimeter ratio of exponential Escherichia coliAB1886 cells pre-exposed to a low intensity red laser and UVC radiation.1) Non-irradiated control, 2) red laser at 8 J/cm2, 3) UVC at 25 mJ/cm2 (white bar) and red laser at 8J/cm2+UVC at 25 mJ/cm2 (black bar),4) UVC at 50 mJ/cm2 (white bar) and red laser at 8 J/cm2+UVC at 50 mJ/cm2 (black bar), and 5) UVC at 100 mJ/cm2 (white bar) and red laser at 8 J/cm2+UVC at 100 mJ/cm2(black bar). Experiments were performed in duplicate and the results are reported as means±SD of three independent assays. *P<0.05, compared to control 1 (non-irradiated cells) (one-way analysis of variance, followed by Tukey’s post hoctest).
Discussion
Our study showed that low-level red laser radiation, at the fluence used for therapeutic applications, did not protect exponential wild-type E. coli cells (AB1157) against the lethal action of UVC radiation (Table 2). In stationary wild-type E. coli cells, the effect of the red laser was dependent on UVC fluence, presenting a protective effect at the lowest UVC fluences and a synergistic effect with UVC radiation (Table 2). In exponentialuvrA-deficient E. coli cells (AB1886), the red laser induced protection against the lethal effect of UVC radiation (Table 2). However, laser-induced protection inuvrA-deficient cells was larger than that observed in wild-typeE. coli cells. This result is in accordance with previous observations that exposure to low-level red lasers induces sub-lethal lesions in DNA molecules (99. Fonseca AS, Moreira TO, Paixao DL, Farias FM, Guimaraes OR, de Paoli S, et al. Effect of laser therapy on DNA damage. Lasers Surg Med 2010; 42: 481-488, doi: 10.1002/lsm.v42:6.
https://doi.org/10.1002/lsm.v42:6....
). The He-Ne laser (632.8 nm) protects wild-type and uvrA-deficient E. colicells against UVC radiation (22. Kohli R, Gupta PK. Irradiance dependence of the He-Ne laser-induced protection against UVC radiation in E. colistrains. J Photochem Photobiol B 2003; 69: 161-167, doi: 10.1016/S1011-1344(03)00018-6.
https://doi.org/10.1016/S1011-1344(03)00...
,1414. Kohli R, Gupta PK, Dube A. Helium-neon laser preirradiation induces protection against UVC radiation in wild-type E. colistrain K12AB1157. Radiat Res 2000; 153: 181-185, doi: 10.1667/0033-7587(2000)153[0181:HNLPIP]2.0.CO;2.
https://doi.org/10.1667/0033-7587(2000)1...
). In our study, pre-exposure to low-level red laser radiation increased the lethal effect of UVC radiation E. coli in the stationary growth phase. This finding reinforces that laser-induced effects might be different when physiological conditions in cells are modified. In addition, laser radiation parameters, such as wavelength, fluence, and irradiance, can determine the biological effects. Laser-induced protection could be dependent on UVC fluence and physiological conditions in cells, because in our study, pre-exposure to a red laser did not alter the survival of an exponential wild-type E. coli strain (AB1157) at all UVC fluences evaluated. In fact, a previous study showed that low-level laser effects depend on physiological conditions in the cells (1515. Canuto KS, Sergio LPS, Guimarães OR, Polignano GAC, Geller M, et al. DNA repair in bacterial cultures and plasmid DNA exposed to infrared laser for treatment of pain. Laser Phys Lett 2013; 10: 065606, doi: 10.1088/1612-2011/10/6/065606.
https://doi.org/10.1088/1612-2011/10/6/0...
). However, in our study, laser protection against the lethal effect of UVC radiation was not observed in stationary uvrA-deficient E. coli cells (Table 2). This result is in agreement with previous studies showing that laser-induced effects depend on physiological conditions in cells (16. Marciano RS, Sergio LPS, Polignano GAC, Presta GA, Guimarães OR, Geller M, et al. Laser for treatment of aphthous ulcers on bacteria cultures and DNA. Photochem Photobiol Sci 2012; 11: 14761483, doi: 10.1039/c2pp25027f.
https://doi.org/10.1039/c2pp25027f...
17. da Silva Sergio LP, da Silva Marciano R, Castanheira Polignano GA, Guimarães OR, Geller M, Paoli F, et al. Evaluation of DNA damage induced by therapeutic low-level red laser. J Clin Exp Dermatol Res 2012; 3: 166, doi: 10.4172/2155-9554.1000166.
https://doi.org/10.4172/2155-9554.100016...
161818. Teixeira GR, Marciano RS, Sergio LPS, Polignano GAC, Guimarães OR, Geller M, et al. Infrared laser effects at fluences used for treatment of dentin hypersensitivity on DNA repair in Escherichia coli and plasmids. Opt Laser Technol 2014; 64: 46-52, doi: 10.1016/j.optlastec.2014.04.023.
https://doi.org/10.1016/j.optlastec.2014...
).
To determine whether laser-induced protection against effects of UVC radiation involve other DNA repair mechanisms, we evaluated induction of filamentation. Exposure to UVC radiation induced similar percentages of filamentation in wild-typeE. coli cultures that were not exposed and pre-exposed to a low-level red laser, but the percentage of filamentation was larger in exponential cultures than in stationary cultures in both non- and pre-exposed to a red laser with UVC (Table 3). However, inuvrA-deficient E. coli cultures, in the exponential and stationary growth phases, pre-exposure to a low-level red laser increased the percentages of filamentation at some UVC fluences (Table 3). Filamentation is part of the SOS response, which is a set of physiological and biochemical modifications in response to DNA damage induced by chemical and physical agents (1111. Koch WH, Woodgate R. The SOS response. In: Nickoloff JA, Hoekstra MF (Editors), DNA damage and repair. Vol I: DNA repair in procaryotes and lower eukaryotes. New Jersey: Humana Press; 1998. p 107-134.). There are a few studies on induction of the SOS response in prokaryotic cells exposed to low-level lasers (1010. Fonseca AS, Presta GA, Geller M, Paoli F. Low intensity infrared laser induces filamentation in Escherichia coli cells. Lasers Phys 2011; 21: 1-9, doi: 10.1134/S1054660X11170051.
https://doi.org/10.1134/S1054660X1117005...
,1515. Canuto KS, Sergio LPS, Guimarães OR, Polignano GAC, Geller M, et al. DNA repair in bacterial cultures and plasmid DNA exposed to infrared laser for treatment of pain. Laser Phys Lett 2013; 10: 065606, doi: 10.1088/1612-2011/10/6/065606.
https://doi.org/10.1088/1612-2011/10/6/0...
,1616. Marciano RS, Sergio LPS, Polignano GAC, Presta GA, Guimarães OR, Geller M, et al. Laser for treatment of aphthous ulcers on bacteria cultures and DNA. Photochem Photobiol Sci 2012; 11: 14761483, doi: 10.1039/c2pp25027f.
https://doi.org/10.1039/c2pp25027f...
,1818. Teixeira GR, Marciano RS, Sergio LPS, Polignano GAC, Guimarães OR, Geller M, et al. Infrared laser effects at fluences used for treatment of dentin hypersensitivity on DNA repair in Escherichia coli and plasmids. Opt Laser Technol 2014; 64: 46-52, doi: 10.1016/j.optlastec.2014.04.023.
https://doi.org/10.1016/j.optlastec.2014...
). However, no studies have shown induction of an SOS response by low-level lasers followed by UVC radiation. Laser-induced SOS responses in E. coli cultures have been observed by induction of phr gene expression (22. Kohli R, Gupta PK. Irradiance dependence of the He-Ne laser-induced protection against UVC radiation in E. colistrains. J Photochem Photobiol B 2003; 69: 161-167, doi: 10.1016/S1011-1344(03)00018-6.
https://doi.org/10.1016/S1011-1344(03)00...
,1919. Kohli R, Bose B, Gupta PK. Induction of phr gene expression inE. coli strain KY706/pPL-1 by He-Ne laser (632.8 nm) irradiation. J Photochem Photobiol B 2001; 60: 136-142, doi: 10.1016/S1011-1344(01)00139-7.
https://doi.org/10.1016/S1011-1344(01)00...
). Previous studies have shown that low-level red and near-infrared lasers induce filamentation in exponential and stationary E. coli cultures (1010. Fonseca AS, Presta GA, Geller M, Paoli F. Low intensity infrared laser induces filamentation in Escherichia coli cells. Lasers Phys 2011; 21: 1-9, doi: 10.1134/S1054660X11170051.
https://doi.org/10.1134/S1054660X1117005...
,1616. Marciano RS, Sergio LPS, Polignano GAC, Presta GA, Guimarães OR, Geller M, et al. Laser for treatment of aphthous ulcers on bacteria cultures and DNA. Photochem Photobiol Sci 2012; 11: 14761483, doi: 10.1039/c2pp25027f.
https://doi.org/10.1039/c2pp25027f...
,1818. Teixeira GR, Marciano RS, Sergio LPS, Polignano GAC, Guimarães OR, Geller M, et al. Infrared laser effects at fluences used for treatment of dentin hypersensitivity on DNA repair in Escherichia coli and plasmids. Opt Laser Technol 2014; 64: 46-52, doi: 10.1016/j.optlastec.2014.04.023.
https://doi.org/10.1016/j.optlastec.2014...
). Therefore, our finding of filamentation in cells that were pre-exposed to a laser is in agreement with those previous studies. The highest percentage of filamentation observed inuvrA-deficient E. coli cells could be explained by a possible synergistic effect of the low-level red laser and UVC radiation. In addition, a larger induction of filamentation in E. coli cultures pre-exposed to a red laser could explain the highest survival of these cells exposed to UVC radiation.
The filamentation phenotype can be induced in part among stressed cells in a prokaryotic culture in response to an aggressive agent (2020. Slayden RA, Knudson DL, Belisle JT. Identification of cell cycle regulators in Mycobacterium tuberculosis by inhibition of septum formation and global transcriptional analysis. Microbiology 2006; 152: 1789-1797, doi: 10.1099/mic.0.28762-0.
https://doi.org/10.1099/mic.0.28762-0....
). However, quantification of bacterial filaments does not take into account non-filamentous cells. To evaluate this in our study, the area and perimeter of cells were measured after low-level red laser and UVC radiation exposure. We found that the laser alone or laser use prior to UVC radiation exposure did not alter the area-perimeter ratio of exponential and stationary wild-typeE. coli, suggesting no morphological alteration of cells. However, uvrA-deficient E. coli cells in the exponential growth phase had an increased area-perimeter ratio after red laser pre-exposure and UVC exposure (Figure 2). This result could be related to a protective effect of the red laser against the lethal effect of UVC radiation and the higher percentages of filamentation obtained in exponential uvrA-deficient E. coli cells (Tables 2 and 3). Additionally, results from area/perimeter ratio of non- and pre-exposed to a red laser with UVC (Figure 2) are in concordance with the hypothesis that biological low-level laser effects are dependent on DNA repair mechanisms (2121. Fonseca AS, Geller M, Bernardo Filho M, Valenca SS, de Paoli F. Low-level infrared laser effect on plasmid DNA. Lasers Med Sci 2012; 27: 121-130, doi: 10.1007/s10103-011-0905-2.
https://doi.org/10.1007/s10103-011-0905-...
). Similar to wild-type E. coli, the area-perimeter ratio in stationary uvrA-deficient E. coli cultures was not altered. This finding indicates that laser-induced effects on UVC action are dependent on the physiological condition in cells, at least among cells that are deficient in DNA repair mechanisms.
Taken together, our results suggest that low-level red laser exposure induced free radical generation in biological systems, which induced protective mechanisms against UVC radiation. This could be part, or a consequence, of a laser-induced biostimulation effect, leading to higher cell survival and regeneration in damaged tissues submitted to low-level laser therapy.
Our results showed that a low-level red laser protected cells against the lethal effect of UVC radiation, and induced filamentation and morphological alterations, depending on DNA repair mechanisms and physiological conditions in cells. Therapeutic low-level red laser radiation can induce DNA lesions at a sub-lethal level. Consequences to cells and tissues should be considered when clinical protocols based on this laser are carried out.
This work was supported by FAPERJ (#APQ1-E-26/111.794/2012), FAPEMIG (#APQ 00432/13), and CNPq (#474405/2013-3).
References
-
1Niemz MH. Laser-tissue interactions: Fundamentals and applications. New York: Springer-Verlag; 2007.
-
2Kohli R, Gupta PK. Irradiance dependence of the He-Ne laser-induced protection against UVC radiation in E. colistrains. J Photochem Photobiol B 2003; 69: 161-167, doi: 10.1016/S1011-1344(03)00018-6.
» https://doi.org/10.1016/S1011-1344(03)00018-6. -
3Eells JT, Wong-Riley MT, VerHoeve J, Henry M, Buchman EV, Kane MP, et al. Mitochondrial signal transduction in accelerated wound and retinal healing by near-infrared light therapy. Mitochondrion 2004; 4: 559-567, doi: 10.1016/j.mito.2004.07.033.
» https://doi.org/10.1016/j.mito.2004.07.033. -
4Karu TI, Kalendo GS, Letokhov VS, Lobko VV. Biostimulation of HeLa cells by low intensity visible light. III. Stimulation of nucleic acid synthesis in plateau phase cells. Il Nuovo Cimento D 1984; 3: 319-325, doi: 10.1007/BF02457461.
» https://doi.org/10.1007/BF02457461. -
5Karu T, Pyatibrat L, Kalendo G. Irradiation with He-Ne laser increases ATP level in cells cultivated in vitro J Photochem Photobiol B 1995; 27: 219-223, doi: 10.1016/1011-1344(94)07078-3.
» https://doi.org/10.1016/1011-1344(94)07078-3. -
6Zhang Y, Song S, Fong CC, Tsang CH, Yang Z, Yang M. cDNA microarray analysis of gene expression profiles in human fibroblast cells irradiated with red light. J Invest Dermatol 2003; 120: 849-857, doi: 10.1046/j.1523-1747.2003.12133.x.
» https://doi.org/10.1046/j.1523-1747.2003.12133.x. -
7Avci P, Gupta A, Sadasivam M, Vecchio D, Pam Z, Pam N, et al. Low-level laser (light) therapy (LLLT) in skin: stimulating, healing, restoring. Semin Cutan Med Surg 2013; 32: 41-52.
-
8Kim YG. Laser mediated production of reactive oxygen and nitrogen species; implications for therapy. Free Radic Res 2002; 36: 1243-1250, doi: 10.1080/1071576021000028389.
» https://doi.org/10.1080/1071576021000028389. -
9Fonseca AS, Moreira TO, Paixao DL, Farias FM, Guimaraes OR, de Paoli S, et al. Effect of laser therapy on DNA damage. Lasers Surg Med 2010; 42: 481-488, doi: 10.1002/lsm.v42:6.
» https://doi.org/10.1002/lsm.v42:6. -
10Fonseca AS, Presta GA, Geller M, Paoli F. Low intensity infrared laser induces filamentation in Escherichia coli cells. Lasers Phys 2011; 21: 1-9, doi: 10.1134/S1054660X11170051.
» https://doi.org/10.1134/S1054660X11170051. -
11Koch WH, Woodgate R. The SOS response. In: Nickoloff JA, Hoekstra MF (Editors), DNA damage and repair. Vol I: DNA repair in procaryotes and lower eukaryotes. New Jersey: Humana Press; 1998. p 107-134.
-
12Butts BD, Kwei KA, Bowden GT, Briehl MM. Elevated basal reactive oxygen species and phospho-Akt in murine keratinocytes resistant to ultraviolet B-induced apoptosis. Mol Carcinog 2003; 37: 149-157.
-
13Cappuccino JG, Sherman N. Microbiology: a laboratory manual. California: Benjamin Cummings Science Publishing; 1999.
-
14Kohli R, Gupta PK, Dube A. Helium-neon laser preirradiation induces protection against UVC radiation in wild-type E. colistrain K12AB1157. Radiat Res 2000; 153: 181-185, doi: 10.1667/0033-7587(2000)153[0181:HNLPIP]2.0.CO;2.
» https://doi.org/10.1667/0033-7587(2000)153[0181:HNLPIP]2.0.CO;2. -
15Canuto KS, Sergio LPS, Guimarães OR, Polignano GAC, Geller M, et al. DNA repair in bacterial cultures and plasmid DNA exposed to infrared laser for treatment of pain. Laser Phys Lett 2013; 10: 065606, doi: 10.1088/1612-2011/10/6/065606.
» https://doi.org/10.1088/1612-2011/10/6/065606 -
16Marciano RS, Sergio LPS, Polignano GAC, Presta GA, Guimarães OR, Geller M, et al. Laser for treatment of aphthous ulcers on bacteria cultures and DNA. Photochem Photobiol Sci 2012; 11: 14761483, doi: 10.1039/c2pp25027f.
» https://doi.org/10.1039/c2pp25027f -
17da Silva Sergio LP, da Silva Marciano R, Castanheira Polignano GA, Guimarães OR, Geller M, Paoli F, et al. Evaluation of DNA damage induced by therapeutic low-level red laser. J Clin Exp Dermatol Res 2012; 3: 166, doi: 10.4172/2155-9554.1000166.
» https://doi.org/10.4172/2155-9554.1000166 -
18Teixeira GR, Marciano RS, Sergio LPS, Polignano GAC, Guimarães OR, Geller M, et al. Infrared laser effects at fluences used for treatment of dentin hypersensitivity on DNA repair in Escherichia coli and plasmids. Opt Laser Technol 2014; 64: 46-52, doi: 10.1016/j.optlastec.2014.04.023.
» https://doi.org/10.1016/j.optlastec.2014.04.023. -
19Kohli R, Bose B, Gupta PK. Induction of phr gene expression inE. coli strain KY706/pPL-1 by He-Ne laser (632.8 nm) irradiation. J Photochem Photobiol B 2001; 60: 136-142, doi: 10.1016/S1011-1344(01)00139-7.
» https://doi.org/10.1016/S1011-1344(01)00139-7. -
20Slayden RA, Knudson DL, Belisle JT. Identification of cell cycle regulators in Mycobacterium tuberculosis by inhibition of septum formation and global transcriptional analysis. Microbiology 2006; 152: 1789-1797, doi: 10.1099/mic.0.28762-0.
» https://doi.org/10.1099/mic.0.28762-0. -
21Fonseca AS, Geller M, Bernardo Filho M, Valenca SS, de Paoli F. Low-level infrared laser effect on plasmid DNA. Lasers Med Sci 2012; 27: 121-130, doi: 10.1007/s10103-011-0905-2.
» https://doi.org/10.1007/s10103-011-0905-2.
-
First published online.
Publication Dates
-
Publication in this collection
10 July 2015 -
Date of issue
Oct 2015
History
-
Received
18 Oct 2014 -
Accepted
23 Apr 2015