Resumos
A irradiação de alimentos é um tratamento pós-colheita bastante promissor, capaz de atuar tanto na desinfestação como no aumento da vida útil de frutos. O mamão é um fruto com curta vida pós-colheita, levando de 6 a 12 dias para atingir a senescência. Além disto, a infestação por larvas de mosca diminui sensivelmente o seu potencial de exportação. O objetivo deste trabalho foi verificar a ação de uma dose de 0,5 kGy de irradiação gama, normalmente utilizada para desinfestação e aumento de vida útil dos frutos, sobre o comportamento dos carboidratos solúveis durante o amadurecimento do mamão (Carica papaya L. cv. Solo), e sobre as enzimas relacionadas ao metabolismo da sacarose: sacarose sintase (SS), sacarose-fosfato sintase (SPS) e invertases ácida e neutra. Os resultados demonstraram que a irradiação não exerce efeito sobre o teor de glicose e frutose, ou sobre a respiração e atividade da SS. Entretanto, a composição dos açúcares solúveis totais e da sacarose, e a atividade da SPS e das invertases foram afetados.
Mamão; irradiação gama; sacarose; sacarose sintase; sacarose-fosfato sintase; invertases
Food irradiation is one of the most promising treatments that can be utilized for fruits desinfestation and extension of shelf life. The authors studied the influence of 0,5 kGy of Gamma irradiation on the soluble carbohydrates composition of papaya (Carica papaya L. cv. Solo) fruit, and on sucrose metabolizing enzymes: sucrose synthase (SS), sucrose-phosphate synthase (SPS), acid and neutral invertases activities, during ripening. The results demonstred that ethylene production, total soluble sugars, sucrose content, and sucrose-phosphate synthase and invertases activities were affected by irradiation, but not respiration, glucose and fructose content, and SS activity.
papaya; gamma irradiation; sucrose; sucrose synthase; sucrose-phosphate synthase; invertases
Metabolismo de carboidratos durante o amadurecimento do mamão (Carica papaya L. cv. Solo): influência da radiação gama1 1 Recebido para publicação em 27/10/98. Aceito para publicação em 29/04/99.
M. L. P. A. GOMEZ2 1 Recebido para publicação em 27/10/98. Aceito para publicação em 29/04/99. , F. M. LAJOLO3 1 Recebido para publicação em 27/10/98. Aceito para publicação em 29/04/99. , B. R. CORDENUNSI3 1 Recebido para publicação em 27/10/98. Aceito para publicação em 29/04/99. ,* 1 Recebido para publicação em 27/10/98. Aceito para publicação em 29/04/99.
RESUMO
A irradiação de alimentos é um tratamento pós-colheita bastante promissor, capaz de atuar tanto na desinfestação como no aumento da vida útil de frutos. O mamão é um fruto com curta vida pós-colheita, levando de 6 a 12 dias para atingir a senescência. Além disto, a infestação por larvas de mosca diminui sensivelmente o seu potencial de exportação. O objetivo deste trabalho foi verificar a ação de uma dose de 0,5 kGy de irradiação gama, normalmente utilizada para desinfestação e aumento de vida útil dos frutos, sobre o comportamento dos carboidratos solúveis durante o amadurecimento do mamão (Carica papaya L. cv. Solo), e sobre as enzimas relacionadas ao metabolismo da sacarose: sacarose sintase (SS), sacarose-fosfato sintase (SPS) e invertases ácida e neutra. Os resultados demonstraram que a irradiação não exerce efeito sobre o teor de glicose e frutose, ou sobre a respiração e atividade da SS. Entretanto, a composição dos açúcares solúveis totais e da sacarose, e a atividade da SPS e das invertases foram afetados.
Palavras-chave: Mamão, irradiação gama, sacarose, sacarose sintase, sacarose-fosfato sintase, invertases.
SUMMARY
Influence of gamma radiation on carbohydrates metabolism of ripening papaya (Carica papaya L. cv. Solo)
. Food irradiation is one of the most promising treatments that can be utilized for fruits desinfestation and extension of shelf life. The authors studied the influence of 0,5 kGy of Gamma irradiation on the soluble carbohydrates composition of papaya (
Carica papaya
L. cv. Solo) fruit, and on sucrose metabolizing enzymes: sucrose synthase (SS), sucrose-phosphate synthase (SPS), acid and neutral invertases activities, during ripening. The results demonstred that ethylene production, total soluble sugars, sucrose content, and sucrose-phosphate synthase and invertases activities were affected by irradiation, but not respiration, glucose and fructose content, and SS activity.
Keywords: papaya, gamma irradiation, sucrose, sucrose synthase, sucrose-phosphate synthase, invertases.
1 INTRODUÇÃO
Frutos são componentes essenciais da alimentação humana, devido ao aporte significativo de vitaminas, minerais e carboidratos. Após a colheita, as estruturas se encontram mais suscetíveis à degradação, e uma vez desligadas da planta, inicia-se um contínuo processo de modificações metabólicas que levam ao desenvolvimento de importantes características sensoriais, como adoçamento e mudanças de coloração e textura. Para conter o amadurecimento e/ou eliminar a infestação por larvas de moscas, aumentando a vida útil dos frutos, são utilizados diversos procedimentos após a colheita que podem ser químicos, como aplicação de substâncias inseticidas, ou físicos, como tratamento térmico, armazenamento a baixas temperaturas e irradiação [23].
A aplicação de substâncias químicas foi utilizada durante muito tempo no controle da infestação de frutos por larvas de mosca, mas devido a problemas relacionados à saúde dos trabalhadores, dos consumidores e a questões ambientais, o uso de potentes inseticidas, como o 1,2-dibromoetano (EDB), foi proibido desde 1984 nos Estados Unidos. O tratamento com inseticidas também pode modificar o metabolismo de maneira indesejável, diminuindo, de certa forma, a qualidade dos frutos [23].
A aplicação de tratamento térmico, na forma de imersão em água a 40-45oC, é bastante eficiente contra a infestação por larvas de mosca; entretanto, pode acarretar uma aceleração descontrolada do metabolismo, levando a uma diminuição da vida útil do fruto. O armazenamento a baixas temperaturas (cerca de 3oC) é um dos procedimentos mais utilizados para a contenção do amadurecimento de frutos; no entanto, frutos tropicais são particularmente sensíveis a baixas temperaturas, o que pode ocasionar injúrias devido ao frio ("chilling injury"), como manchas na casca ou ressecamento.
A irradiação é um tratamento pós-colheita que pode ser utilizado tanto para a desinfestação quanto para o aumento da vida útil, oferecendo, desta forma, frutos de maior qualidade. A irradiação tem, também, a vantagem de penetrar uniformemente no tecido vegetal, atuando em qualquer etapa de desenvolvimento das larvas de mosca, como a Anastrepha sp., além de não deixar resíduos [13, 19, 25]. Entretanto, o maior empecilho para o uso da irradiação como tratamento pós-colheita de frutos é o alto custo para implantação do sistema. Apesar do uso da irradiação em alimentos ser permitido desde 1970 por entidades como a Organização das Nações Unidas para Agricultura e Alimentação (FAO) e Organização Internacional de Energia Atômica (OIEA), há ainda o alto preconceito por parte dos consumidores com relação a alimentos irradiados [24].
Para que a irradiação tenha sucesso, é necessário adequar a dose aplicada a cada fruto, de modo a evitar reações indesejáveis, como amolecimento indiscriminado ou escurecimento da casca. Deve-se levar em consideração a cultivar, tamanho, e estádio de amadurecimento dos frutos, uma vez que a eficácia do tratamento varia com estas características [13, 15]. Dentre os frutos estudados, o mamão parece ser o mais adequado à irradiação, pois apresenta poucos efeitos colaterais, relacionados diretamente ao processamento. O aumento na respiração ocasionado pela irradiação é mínimo [2], e retorna aos níveis normais em 24 horas [19]. As doses para a desinfestação e aumento da vida útil do mamão variam entre 0,5 e 1,0 kGy, tendo em 0,75 kGy a dose ótima, que deve ser aplicada quando o fruto atinge, no máximo, 30% de amarelecimento da casca [4, 18]. Nesta fase, a aplicação de 0,5 kGy de radiação é capaz de reter a firmeza de mamões por 2 a 3 dias, o que pode significar um aumento de 30 a 50% na vida útil do fruto [10, 26]. Esta contenção é devida a uma diminuição das atividades das enzimas de hidrólise da parede celular, como pectinesterases, b-galacto-sidases, celulases, entre outras. Não se tem porém, informação sobre um possível efeito no metabolismo de carboidratos.
O sabor doce, em conjunto com mudanças de coloração e textura é um dos principais parâmetros de qualidade dos frutos, sendo um dos mais exigidos pelos consumidores. Durante o amadurecimento, o adoçamento pode ser originado pelo acúmulo de sacarose originada pela fotossíntese, ou por hidrólise de carboidratos de reserva. Frutos não-climatéricos são incapazes de sintetizar grandes quantidades de açúcares após a colheita, a não ser em proporções muito pequenas, como é o caso da laranja.
Alguns frutos climatéricos, como a banana [9] e o kiwi [22], apresentam grandes quantidades de amido no fruto ainda verde, que são degradadas durante o amadurecimento, após a colheita, resultando em quantidades significativas de sacarose no fruto maduro. O mamão não possui quantidades significativas de amido que possam ser convertidas em açúcares durante o amadurecimento, existindo controvérsias sobre um possível adoçamento pós colheita [8, 19].
Neste trabalho foram acompanhados a evolução dos açúcares solúveis durante o amadurecimento do mamão e enzimas correlacionadas à sua síntese/degradação, de forma a verificar possíveis alterações, decorrentes de uma dose de 0,5kGy de irradiação gama, a exemplo do que acontece com as hidrolases da parede celular [10].
2 MATERIAL E MÉTODOS
2.1 Frutos
Foram utilizados mamões (Carica papaya L. cv. Solo) obtidos no mercado local, que apresentassem apenas 10% de coloração amarela da casca, cerca de 72h após a colheita. Dos frutos escolhidos, a metade (oito frutos) foi submetida à irradiação gama por fonte de Co65, na dose de 0,5kGy. Como grupo controle, foram utilizados oito frutos não-irradiados.
2.2 Amostragem
A amostragem foi realizada através da remoção de batoques, ao longo do amadurecimento dos frutos, que foram mantidos em câmara incubadora a 18oC e 95% de umidade relativa do ar, durante 10 dias. Foram utilizados 8 frutos irradiados e 8 frutos não-irradiados, perfazendo uma amostra significativa. Diariamente, foi retirado, da porção equatorial de cada fruto, um cilindro com 1,8cm de diâmetro e aproximadamente 5cm de profundidade, com o auxílio de um furador de rolhas metálico. Para evitar contaminações, o local da incisão foi previamente higienizado com solução de hipoclorito de sódio a 5%, e o furador de rolhas, esterilizado por flambagem. Cada amostra (irradiado e controle) foi obtida com o agrupamento dos 8 batoques respectivos ao tratamento, que foram fatiados, imediatamente congelados em nitrogênio líquido, e armazenados em freezer a -80oC até o momento da análise. Os furos resultantes nos frutos foram vedados com parafina comum, para amenizar a injúria.
2.3 Respiração e etileno
As análises de etileno foram feitas diariamente, deixando-se os frutos em frascos fechados por exatamente uma hora, quando amostras do ar do interior do frasco foram retiradas com uma seringa de 1mL, e aplicadas em cromatógrafo a gás, equipado com coluna Porapack Q e detector de ionização de chama (FID). Como padrão, foi utilizado uma mistura de ar sintético contendo 300ppm de etileno. A respiração foi determinada quantificando o CO2 produzido pelos frutos mantidos em frascos fechados conectados a um analisador de gases por infravermelho. A leitura foi realizada após dez minutos de circulação de ar, quando considerada estabilizada. O zero da escala do aparelho foi ajustado com o ar ambiente, livre de umidade e de CO2, através de passagem por colunas de sílica gel e cal sodada. Como padrão, foi utilizado ar sintético contendo 350ppm de CO2.
2.4 Determinação dos carboidratos
As análises de amido foram realizadas de acordo com CHAMP [7], após extração dos açúcares solúveis, através de incubação das amostras com a -amilase termoresistente a 100oC, gelatinização com KOH 7M em gelo e hidrólise a 37oC com amiloglicosidase, sendo a glicose liberada determinada pelo sistema glicose-oxidase/pero-xidase/ABTS [5]. Os açúcares solúveis foram extraídos com etanol 80% a 80oC e determinados enzimaticamente, pelo sistema de enzimas hexoquinase, glicose-6-fosfato desidrogenase, fosfoglicose isomerase e invertase, medindo-se redução do NADP a 340nm, segundo BERGMEYER [5]. Os carboidratos de parede celular solúveis em água (PSA) foram determinados com as amostras liofilizadas, após extração dos açúcares solúveis em etanol, por solubilização em água a 90oC e precipitação com etanol a 80%. Após hidrólise com H2SO4 72% a 121oC/1h, as amostras foram neutralizadas com NaOH grau HPLC e determinadas por HPLC acoplado a detector de pulso amperométrico (PAD).
2.5 Extração das enzimas sacarose sintase (SS), sacarose-fosfato sintase (SPS) e invertases
Para a extração da SS e da SPS, as amostras, previamente pulverizadas com nitrogênio líquido, foram homogeneizadas em Potter, na proporção de 1g para 1ml de solução extratora. O homogenato resultante foi centrifugado a 20000 x g/10min., e o sobrenadante foi dessalinizado através de coluna Sephadex G-25 HiTrap Desalting (Pharmacia). A solução extratora foi composta por Tris-HCl 500mM pH7,5 contendo EDTA 40mM, glicerol 15% (v/v), ácido ascórbico 50mM e NaCl 1M.
A extração das invertases seguiu o mesmo procedimento, sendo a solução extratora composta por tampão fosfato 50mM pH 7,5 contendo NaCl 50mM, glicerol 5%, sulfato de manganês 5mM e b-mercaptoetanol 1mM.
2.6 Determinação das atividades da SS, SPS e invertases
A atividade da SS foi determinada em meio saturante composto por 60mL de Tris-HCl 54mM pH 7,5, contendo frutose 20mM, UDP-glicose 20mM, cloreto de magnésio 30mM e 40mL de extrato enzimático. A mistura foi incubada a 37oC e a reação interrompida após 20 minutos com a adição de NaOH 1M. A sacarose formada foi determinada pelo método do ácido tiobarbitúrico (TBA), descrito por PERCHERON [20]. A atividade da SPS foi determinada da mesma maneira, sendo o meio de atividade composto por HEPES-KOH 95mM pH 7,5, contendo frutose-6-fosfato 20mM, UDP-glicose 20mM, cloreto de magnésio 30mM. A atividade da invertase ácida foi determinada por incubação de 20 mL do extrato enzimático com 20mL de sacarose 0,6M, 60mL de tampão acetato de sódio 0,2 M pH 4,6 contendo 0,01% de albumina de soro bovina e 100 mL de água purificada, a 30oC por 10 minutos, sendo a reação interrompida por fervura da mistura por 1 minuto. A glicose resultante foi determinada pelo sistema glicose-oxidase/peroxidase/ABTS, utilizando glicose como padrão. A atividade da invertase neutra foi determinada da mesma forma, substituindo-se o tampão acetato de sódio 0,2 M pH 4,6 por tampão fosfato de sódio 0,2M pH 7,0.
2.7 Determinação de proteína
A quantificação da proteína nos extratos enzimáticos foi feita pelo método de BRADFORD [6], utilizando albumina de soro bovino como padrão.
3 RESULTADOS E DISCUSSÃO
A colheita do mamão é feita baseada na coloração da casca, sem nenhum outro critério mais objetivo, resultando em frutos com grande variação nos estádios de maturação. Assim, houve a necessidade de otimizarmos um sistema de amostragem, onde fossem analisados sempre os mesmos frutos durante todo o processo de amadurecimento, através da retirada diária de um batoque da região equatorial dos frutos. O modelo se mostrou bastante adequado, sendo que os frutos amadureceram normalmente, mantendo as mesmas características dos frutos inteiros. Em experimentos preliminares (dados ainda não publicados), foi verificado não haver diferença significativa, no que diz respeito à respiração e produção de etileno, entre o amadurecimento dos frutos submetidos à amostragem por meio de batoques e dos frutos que amadureceram inteiros.
3.1 Respiração e Produção de Etileno
A classificação de frutos em climatéricos e não-climatéricos é definida pelo aumento da respiração após a colheita, associada ao aumento da produção de etileno. O mamão é classificado como fruto climatérico, com um período pré-climatério extenso, seguido de um pico climatérico bem definido [17, 24]. A irradiação, quando em doses inadequadas, pode levar a um aumento da respiração de frutos, devido à liberação repentina e descontrolada de fósforo inorgânico pelas células danificadas, contribuindo para o aumento imediato que ocorre na respiração de frutos irradiados [15]. Nossos resultados confirmam dados da literatura, de que doses de 0,5 kGy não são deletérias para o fruto, pois não houve mudança significativa na respiração dos frutos irradiados durante o amadurecimento (Figura 1A), a não ser por um pequeno aumento na produção de CO2 nos 3 primeiros dias após a colheita, comportamento característico de frutos irradiados [18].
A produção de etileno (Figura 1B) no entanto, foi bastante afetada pela irradiação. O aumento de produção de etileno, característico dos frutos climatéricos, que aconteceu do 4o ao 6o dpc nos mamões controle, não ocorreu nos mamões irradiados. Este mesmo fenômeno já foi verificado em pêras [14], o que sugere que o atraso no amadurecimento dos frutos irradiados pode ser devido à menor capacidade destes frutos produzirem etileno, já que este hormônio tem um papel estimulante do metabolismo em geral e parece estar envolvido na ativação ou regulação de algumas das enzimas envolvidas no amadurecimento. Os valores em elevação de etileno e CO2 obtidos no final do amadurecimento (10odpc) são devidos, provavelmente, ao início do processo de senescência dos frutos.
3.2 Carboidratos
Os teores de amido dos frutos irradiados foram cerca de 40% mais altos que os dos frutos controle, em todos os estádios de amadurecimento analisados, exceto em frutos completamente maduros (Tabela 1). Mas todos os valores estão abaixo de 0,2%, o que torna o amido um componente sem grande importância na composição química do mamão. Estes resultados estão de acordo com os apresentados por PAULL [19], CHAN et al. [8] e SELVARAJ et al. [24], que encontraram 0,5% de amido em frutos verdes e menos de 0,1% em frutos maduros. Não existem dados na literatura em relação a amido em mamões submetidos à irradiação.
Os açúcares solúveis totais, contabilizados como a soma dos teores de glicose, frutose e sacarose, variaram de 9% no estágio verde (3odpc) a cerca de 11% no estágio maduro (10odpc). Os perfis destes mesmos açúcares foram discretamente menores nos frutos irradiados. Estes resultados confirmam os dados encontrados por PAULL e CHAN et al. [8, 18]), de que os açúcares solúveis são, em sua maior parte, acumulados quando o fruto está ainda ligado à árvore, provavelmente em função da fotossíntese. Mas, parece haver, em pequena proporção, síntese de sacarose no período pós colheita, já que os teores de açúcares solúveis totais variaram muito no decorrer do amadurecimento dos frutos. Os teores de glicose (cerca de 3%) e frutose (cerca de 2%) foram sempre os mesmos, exceto ao final do amadurecimento, quando aumentaram, provavelmente em função da atividade das invertases (Figura 2).
Nos mamões controle, os níveis de sacarose variaram de 2 a 5% no decorrer do amadurecimento, enquanto que nos frutos irradiados esses níveis foram discretamente menores, variando entre 0,5 e 4,0%. No 6o dpc ocorreu uma queda nos teores de sacarose, coincidente com o pico de respiração dos frutos. No 1o dia após a colheita, o teor de sacarose nos frutos irradiados foi cerca de 2,5 vezes menor que o controle, e aos 10odpc foi cerca de 6 vezes menor que o controle, possivelmente devido ao tratamento. Entretanto, percebe-se que, nos frutos irradiados, a sacarose que parece ter sido hidrolisada devido ao processamento, retornou ao nível anterior, o mesmo acontecendo após o pico respiratório, tanto em frutos irradiados quanto em frutos não-irradiados. Em batatas e batatas doces irradiadas com doses de 0,2 kGy, entretanto, foi verificado que o conteúdo de sacarose aumenta significativamente durante o período de armazenamento [1, 11, 12].
Testes sensoriais, feitos de maneira informal no laboratório, sinalizaram que não existem diferenças significativas de sabor entre frutos irradiados e não-irradiados.
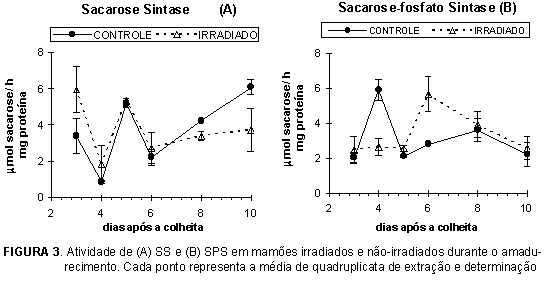
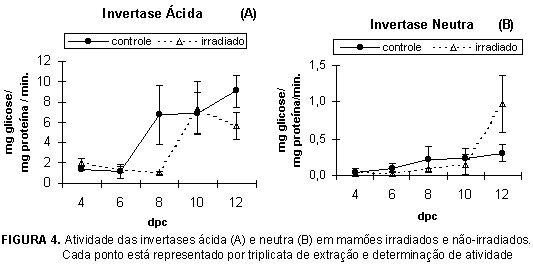
Havendo síntese de sacarose após a colheita do mamão, tornou-se importante verificar a atividade das enzimas relacionadas ao metabolismo da sacarose. Os perfis de atividade da sacarose sintase (SS) e sacarose-fosfato sintase (SPS) ao longo do amadurecimento encontram-se na Figura 3, e invertases ácida e neutra na Figura 4.
A atividade da SS teve um pico aos 5 dpc, coincidente com o período da hidrólise da sacarose, no pico respiratório (Figura 2C). Apenas com estes dados não podemos afirmar se há ou não correlação entre ambos, mas NIELSEN et al. [16] demonstraram que, em pimenta doce, ocorre uma degradação da sacarose no período final do crescimento do fruto, provavelmente originada pela ação da sacarose sintase. Também verificamos que as invertases (Figura 4) não apresentam, neste ponto, atividade tal que possa estar relacionada à hidrólise da sacarose, fortalecendo a hipótese de que a SS seja a responsável por esta hidrólise, nesta fase. A irradiação não parece ter afetado significativamente a atividade da SS, uma vez que o pico de atividade ocorreu no mesmo período e com a mesma intensidade, tanto nos frutos tratados nos não-tratados.
A atividade da SPS durante o amadurecimento do mamão foi baixa, se compararmos com a banana, por exemplo [9], tendo um pico de atividade aos 4 dpc. O atraso de dois dias deste pico de atividade no caso dos frutos irradiados sugere que, ao contrário da SS, o processo de irradiação interferiu na atividadade da SPS. HAYASHI & KAWASHIMA [12] e AJLOUNI & RAMDY [1] observaram que, em batatas, a irradiação causou o aparecimento de um pico de atividade de SS e SPS, que não ocorreu nos tubérculos não-irradiados, mas que retornaram aos níveis originais após algum tempo de armazenamento.
O aumento da sacarose acumulada em batatas irradiadas, é devido à menor hidrólise e maior síntese da sacarose, a partir de UDP-glicose e frutose-6-fosfato, associadas à maior síntese da proteína SPS [12]. Em mamões, poderíamos sugerir que a atividade da SPS esteja correlacionada apenas com a presença de sacarose acumulada, pois apesar de os teores de açúcares solúveis totais nos mamões irradiados serem menores que nos mamões não-irradiados, a atividade da SPS não é significativamente diferente, em termos numéricos. Nos mamões irradiados, o pico de atividade da SPS parece coincidir com o ponto onde está havendo hidrólise da sacarose, provavelmente associada ao aumento da respiração (6odpc), necessitando de maior síntese de sacarose.
É nítida a influência da irradiação nas atividades das invertases ácida e neutra (Figura 4). O perfil de atividade da invertase ácida nos frutos irradiados foi mantido, mas o brusco aumento, que nos frutos controle aconteceu no 4o dpc, foi retardado em dois dias, da mesma forma que a atividade da SPS. Entretanto, a irradiação pareceu acelerar a atividade da invertase neutra apenas no final do período, em cerca de 6,3 vezes. Estes resultados poderiam explicar a hidrólise mais acentuada da sacarose nos frutos irradiados no final do amadurecimento (Figura 2C), cerca de 6 vezes maior que em frutos não-irradiados. Em comparação à invertase ácida, a atividade da invertase neutra foi cerca de 5 vezes menor, em frutos irradiados, e 20 vezes menor em mamões irradiados.
4 CONCLUSÕES
O processo de irradiação de mamões com 0,5 kGy não afetou a respiração (produção de CO2) e a atividade da sacarose sintase; mas atrasou em cerca de dois dias o aumento nas atividades da SPS e das invertases ácida e neutra, verificados nos frutos controle. Não afetou também o adoçamento, já que os teores de açúcares totais nos mamões tratados foi apenas cerca de 2% menor em todas as fases do amadurecimento. Conclui-se então que, quanto ao aspecto dos carboidratos, mamões podem ser irradiados com 0,5kGy, sem que seja afetado o adoçamento e o sabor dos frutos. Pode-se concluir também que degradação da sacarose verificada no 5o dia pós-colheita, parece estar ligada à sacarose sintase e não às invertases.
5 REFERÊNCIAS BIBLIOGRÁFICAS
6 AGRADECIMENTOS
Os autores agradecem à Empresa Brasileira de Radiação, EMBRARAD, na pessoa do Professor Dirceu Vizeu, pela irradiação dos frutos, e ao Conselho de Auxílio à Pesquisa no Ensino Superior (CAPES), pelo auxílio financeiro.
2 Doutoranda em Ciência dos Alimentos, FCF, USP.
3 Professores do Departamento de Alimentos e Nutrição Experimental da FCF, USP.
* A quem a correspondência deve ser enviada.
- [1] AJLOUNI, S., HAMDY, M. K. Effect of combinated gamma-irradiation and storage on biochemical changes in sweet potato. J. Food Sci., Chicago, v. 53, n. 2, p. 477-481, 1988.
- [2] AKAMINE, E. T., GOO, W. Respiration in gamma irradiated fresh fruits. J. Food Sci., Chicago, v. 36, p. 1074-1077, 1971.
- [3] AWAD, M. Fisiologia pós-colheita de fruta. Săo Paulo: Nobel, 1993. 114p.
- [4] BALOCK, J. W., BURDITT, A. K., SEO, S. T., AKAMINE, E. K. Gamma radiation as a quarantine treatment for hawaiian fruit flies. J. Econ. Entomol., Baltimore, v. 59, p. 202, 1966.
- [5] BERGMEYER, H. U., ed. Methods of Enzimatic Analysis 2.ed. New York: Academic Press, 1974. v. 3, p. 1212-1215.
- [6] BRADFORD, M. A rapid and sensitive method for the quantitation of microgram quantities of protein-dye binding. Anal. Biochem., Baltimore, v. 72, p. 248-254, 1976.
- [7] CHAMP, M. Determination of resistant starch in foods and food products: interlaboratory study. Eur. J. Clin. Nutr., London, v. 46, suppl.2, p. 551-562, 1992.
- [8] CHAN Jr., H. T, HIBBARD, K., GOO, T. AKAMINE, E. K. Sugar composition of papayas during fruit development. HortScience, St. Joseph, v. 14, n. 2, p. 140-141, 1979.
- [9] CORDENUNSI, B. R., LAJOLO, F. M. (1995) Starch breakdown during banana ripening: Sucrose Synthase and Sucrose Phosphate Synthase. J. Agric. Food Chem., Washington, v. 43, n. 2, p. 347-351, 1995.
- [10] DINNOCENZO, M. Comportamento de enzimas da parede celular e textura da polpa relacionados ao tratamento de irradiaçăo de mamőes (Carica papaya L. cv. Solo) durante o amadurecimento. Piracicaba, 1996. 85p. (Dissertaçăo de Mestrado) Escola Superior de Agricultura "Luíz de Queiroz". Universidade de Săo Paulo (USP).
- [11] HAYASHI, T., KAWASHIMA, K. Activities of enzymes of sugar metabolism in gamma-irradiated potato tubers. J. Food Sci., Chicago, v. 48, n. 4, p. 1242-1245; 1249, 1983.
- [12] HAYASHI, T., KAWASHIMA, K. Effect of irradiation on the carbohydrate metabolism responsible for sucrose accumulation in potatoes. J. Agric. Food Chem., Washington, v. 33, p. 14-17, 1985.
- [13] IADEROZA, M., BLEINROTH, E. W., AZUMA, E. H. Efeitos da radiaçăo ionizante na atividade da pectinesterase do mamăo cultivar Solo. Colet. Inst. Tecnol. Aliment., Campinas, v. 18, n. 1, p. 76-82, 1988.
- [14] MAXIE, E. C., ABDEL-KADER, A. Food irradiation-physiology of fruits as related to feaseability of the technology. Adv. Food Res., New York, v. 15, p. 105-143, 1966.
- [15] MURRAY, D. R. Biology of food irradiation London: Research Studies Press, 1990. 255p.
- [16] NIELSEN, T. H., SKJAERBAEK, H. C., KARLSEN, P. Carbohydrate metabolism during fruit development in sweet pepper (Capsicum annuum) plants. Physiol. Plant., Copenhagen, v. 82, n. 2, p. 311-319, 1991.
- [17] PAL, D. K., SELVARAJ, Y. Biochemistry of papaya (Carica papaya L.) fruit ripening: changes in RNA, DNA, protein and enzymes of mitochondrial, carbohydrate, respiratory and phosphate metabolism. J.Hortic. Sci., London, v. 62, p. 117-124, 1987.
- [18] PAULL, R. E. Response of Tropical commodities to insect desinfestation treatments. HortScience, St. Joseph, v. 29, n. 9, p. 988-996, 1994.
- [19] PAULL, R. E. Pineapple and Papaya. In: SEYMOUR, G. B., TAYLOR, J. E., TUCKER, G. A. Biochemistry of fruit ripening London: Chapman & Hall, 1996. p. 302-315.
- [20] PERCHERON, F. Colorimetric determination of sucrose and fructofuranosidase by the tiobarbituric acid reaction. C. R. Hebd. Seances Acad. Sci., Paris, v. 255, p. 2521-2522, 1962.
- [21] PRIETO-MIRANDA, E. F.(1997) Uso de irradiación como tratamiento cuarentenário. Tratamientos Físicos de Cuarentena en Frutos Tropicales e Subtropicales (CYTED) La Habana, junio. p. 53-57.
- [22] REDGWELL, R. J., HARKER, R. Softening of kiwifruits discs: effect of inhibition of galactose loss from cell walls. Phytochesmitry, Oxford, v. 39, n. 6, p. 1319-1323, 1995.
- [23] SAUCEDO-VELÓZ, C. Tratamientos físicos de cuarentena. Tratamientos Físicos de Cuarentena en Frutos Tropicales e Subtropicales (CYTED) La Habana, junio. p. 7-14, 1997.
- [24] SELVARAJ, Y., SUBRAMANYAM, M. D., IYER, C. P. A. Changes in the chemical composition of four cultivars of papaya (Carica papaya L.) during growth and development. J. Hort. Sci., London, v. 57, p. 135-143, 1982.
- [25] THOMAS, P. Radiation preservation of foods of plant origin. III. Tropical fruits: bananas, mangoes and papayas. Crit. Rev. Food Sci. Nutr., Cleveland, v. 23, n. 2, p. 147-205, 1986.
- [26] ZHAO, M., MOY, J., PAULL, R. E. Effect of gamma-irradiation on ripening papaya pectin. Postharvest Biol. Technol., Amsterdam, v. 8, p. 209-222, 1996.
Datas de Publicação
-
Publicação nesta coleção
09 Fev 2000 -
Data do Fascículo
Maio 1999
Histórico
-
Aceito
29 Abr 1999 -
Recebido
27 Out 1998