Resumos
O ovo é o produto de eficiente transformação biológica feita pela galinha (Gallus gallus) de postura, com alta qualidade nutricional para o consumo humano. Na gema do ovo encontra-se a proteína denominada fosvitina, que atua como carreadora do ferro se ligando a ele. A deficiência de ferro ocorre normalmente devido ao consumo insuficiente de alimentos fontes de ferro ou à baixa biodisponibilidade. O objetivo da pesquisa foi verificar a concentração de proteínas, em especial a fosvitina/lipovitelina presente em gemas de ovos crus e cozidos nos diferentes ciclos de postura (inicial, intermediário e final) e avaliar a interferência desta proteína na disponibilidade do micronutriente ferro. Os tratamentos foram realizados em diferentes períodos e utilizaram ovos crus e cozidos, sendo gemas cruas de poedeiras em início, meio e fim de postura e gemas cozidas nos três ciclos de postura. Foram realizadas as análises de composição centesimal, concentração de ferro total e dialisável, concentração de proteínas em gemas e eletroforese em gel de poliacrilamida (SDS-PAGE). Houve diferença na concentração de proteínas e diferentes níveis de ferro dialisável nos três ciclos de postura, porém em gemas cruas e cozidas. Com o passar dos ciclos (inicial, intermediário e final), as poedeiras mais velhas, apresentaram ovos (gemas) com um teor mais elevado de ferro disponível, porém uma quantidade inferior de proteínas em especial a fosvitina/lipovitelina, comparando-se com os ciclos inicial e intermediário. A disponibilidade de ferro foi mais elevada em gemas que passaram pelo processo de cocção, porém em poedeiras de postura final.
ovo; gema; ferro; idade; disponibilidade; poedeiras
The egg is the product of efficient biological transformation made by a laying hen. It has high nutritional value for human consumption. Most of the iron in egg yolk is bound by phosvitin, a major protein present in the egg yolk. Iron deficiency normally occurs due to insufficient intake of dietary iron or its low the bioavailability. The objective of this research was to verify the protein concentration, specially the phosvitin/lipovitelin in egg yolks of raw and cooked eggs in different posture cycles (initial, intermediate, final), and to evaluate the interference of this protein in the availability of the iron micronutrient. The treatments were carried in different periods using raw and cooked eggs. The yolks of raw eggs of laying hens were used in the initial, intermediate and final phases of posture and yolks of cooked eggs in the three posture cycles. Analyses of proximal composition, concentration of total and available iron, protein concentration in egg yolks, and eletrophoresis in poliacrilamida (SDS-PAGE) were performed. There was difference in the protein concentration and in the levels of iron availability in the three posture cycles. After running through the cycle (initial, intermediate, and final), the eggs yolks of the older hens presented an increased amount of available iron and lower amount of proteins, specially the fosvitin/lipovitelin, if compared to the initial and intermediate cycles. The highest iron availability content was found in cooked egg yolks and in hens in the final posture cycle.
egg; yolk; iron; age; availability; hens
Concentração de proteínas em gemas de ovos de poedeiras (Gallus gallus) nos diferentes ciclos de postura e sua interferência na disponibilidade do ferro
Protein concentration (phosvitin and lipovitelin) in egg yolks of Laying Hens (Gallus gallus) in different posture cycles and its interference in the iron availability
Érika Vidal SartoriI; Solange Guidolin Canniatti-BrazacaI, * * A quem a correspondência deve ser enviada ; Sandra Helena da CruzII; Salete Aparecida GaziolaII
IDepartamento de Agroindústria, Alimentos e Nutrição, Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo USP, Av. Pádua Dias, 11, CP 9, CEP 13418-900, Piracicaba SP, Brasil, E-mail: esartori@esalq.usp.br; sgcbrazaca@esalq.usp.br; shcruz@esalq.usp.br
IIDepartamento de Genética, Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo USP, Av. Pádua Dias, 11, CP 9, CEP 13418-900, Piracicaba SP, Brasil, E-mail: sagaziol@esalq.usp.br
RESUMO
O ovo é o produto de eficiente transformação biológica feita pela galinha (Gallus gallus) de postura, com alta qualidade nutricional para o consumo humano. Na gema do ovo encontra-se a proteína denominada fosvitina, que atua como carreadora do ferro se ligando a ele. A deficiência de ferro ocorre normalmente devido ao consumo insuficiente de alimentos fontes de ferro ou à baixa biodisponibilidade. O objetivo da pesquisa foi verificar a concentração de proteínas, em especial a fosvitina/lipovitelina presente em gemas de ovos crus e cozidos nos diferentes ciclos de postura (inicial, intermediário e final) e avaliar a interferência desta proteína na disponibilidade do micronutriente ferro. Os tratamentos foram realizados em diferentes períodos e utilizaram ovos crus e cozidos, sendo gemas cruas de poedeiras em início, meio e fim de postura e gemas cozidas nos três ciclos de postura. Foram realizadas as análises de composição centesimal, concentração de ferro total e dialisável, concentração de proteínas em gemas e eletroforese em gel de poliacrilamida (SDS-PAGE). Houve diferença na concentração de proteínas e diferentes níveis de ferro dialisável nos três ciclos de postura, porém em gemas cruas e cozidas. Com o passar dos ciclos (inicial, intermediário e final), as poedeiras mais velhas, apresentaram ovos (gemas) com um teor mais elevado de ferro disponível, porém uma quantidade inferior de proteínas em especial a fosvitina/lipovitelina, comparando-se com os ciclos inicial e intermediário. A disponibilidade de ferro foi mais elevada em gemas que passaram pelo processo de cocção, porém em poedeiras de postura final.
Palavras-chave: ovo; gema; ferro; idade; disponibilidade; poedeiras.
ABSTRACT
The egg is the product of efficient biological transformation made by a laying hen. It has high nutritional value for human consumption. Most of the iron in egg yolk is bound by phosvitin, a major protein present in the egg yolk. Iron deficiency normally occurs due to insufficient intake of dietary iron or its low the bioavailability. The objective of this research was to verify the protein concentration, specially the phosvitin/lipovitelin in egg yolks of raw and cooked eggs in different posture cycles (initial, intermediate, final), and to evaluate the interference of this protein in the availability of the iron micronutrient. The treatments were carried in different periods using raw and cooked eggs. The yolks of raw eggs of laying hens were used in the initial, intermediate and final phases of posture and yolks of cooked eggs in the three posture cycles. Analyses of proximal composition, concentration of total and available iron, protein concentration in egg yolks, and eletrophoresis in poliacrilamida (SDS-PAGE) were performed. There was difference in the protein concentration and in the levels of iron availability in the three posture cycles. After running through the cycle (initial, intermediate, and final), the eggs yolks of the older hens presented an increased amount of available iron and lower amount of proteins, specially the fosvitin/lipovitelin, if compared to the initial and intermediate cycles. The highest iron availability content was found in cooked egg yolks and in hens in the final posture cycle.
Keywords: egg; yolk; iron; age; availability; hens.
1 Introdução
O ovo é o produto de eficiente transformação biológica feita pela galinha (Gallus gallus) de postura. Esta ave transforma recursos alimentares de menor valor biológico em produto com alta qualidade nutricional para o consumo humano (BERTECHINI, 2005).
Dentre os vários alimentos disponíveis, o ovo é o que mais se aproxima de um perfeito balanço de todos os nutrientes: é fonte absoluta de nutrição para o embrião, sendo valioso pela sua qualidade nutricional, flavor e outras propriedades funcionais, quando comparado a outros alimentos (ENSMINGER, 1992; YASHODA et al., 2004).
Segundo Bertechini (2005), o ovo pode ser considerado o maior aliado para reabilitar a nutrição humana, além de outras contribuições nutricionais importantes, sendo de baixo custo.
A produção de ovos comerciais para o consumo cresceu consideravelmente nos últimos anos em todo o mundo (54%) (BERTECHINI, 2005). No primeiro trimestre de 2006, a produção nacional de ovos de galinha (510,885 milhões de dúzias) cresceu 5,47% em relação ao mesmo período de 2005 (484,401 milhões de dúzias). A produção encontra-se distribuída por todo o território nacional, sendo São Paulo (34%), Minas Gerais (13%) e Paraná (8%) os principais Estados produtores (IBGE, 2006).
Segundo a Asociacion de Productores de Huevos do Chile (2005), o ovo apresenta alta qualidade proteica, com elevada concentração de aminoácidos essenciais, e seu conteúdo de vitaminas A, B2 e B12, D e minerais, como ferro, manganês, zinco, fósforo e magnésio. Apenas um ovo supre aproximadamente 10% da ingestão recomendada para adultos de vitamina A e de ácido fólico, 17% da recomendação de vitamina B2 sendo considerável sua contribuição de B12 e vitamina D.
O ferro é essencial para a vida e atua principalmente na síntese das células vermelhas do sangue e no transporte do oxigênio para todas as células do corpo (MINISTÉRIO DA SAÚDE, 2004).
A deficiência de ferro ocorre normalmente devido ao consumo insuficiente de alimentos fontes de ferro ou a baixa biodisponibilidade (MINISTÉRIO DA SAÚDE, 2005). A carência deste micronutriente tem sido a causa mais frequente de anemia em todo o mundo, constituindo-se em grave problema de saúde pública em nosso meio (MELO et al., 2002).
Na gema do ovo encontra-se a proteína denominada fosvitina, que atua como carreadora do ferro se ligando a ele, fazendo com que este não esteja disponível para absorção. Segundo Jiang e Mine (2000), 95% do ferro presente na gema está ligado à fosvitina, numa conformação muito estável.
A fosvitina é sintetizada no fígado de galinhas em postura (SCHIRM; GRUBER, 1973). Ocorrendo a produção de enzimas como a vitelogenina, que é produzida em resposta a estrógenos ovarianos, a vitelogenina é transportada por via sanguínea ao ovário onde é endocitada e fragmentada em duas proteínas vitelínicas: lipovitelina e fosvitina (WALLACE; SELMAN, 1981; MATSUBARA; SAWANO, 1995).
Takata et al. (2001) mencionam que as aves atingem maturidade sexual entre 17 e 18 semanas de idade, e que a postura atinge seu pico entre 28 e 35 semanas de idade, decrescendo gradativamente após este período. Os hormônios vão se alterando de acordo com o ciclo de vida da poedeira. A atividade de muitas espécies de aves é controlada pelo estímulo ambiental. Portanto, a atividade sexual diminui com os dias mais curtos e aumenta naqueles mais longos (SOUZA-SOARES; SIEWERDT, 2005). Moraes (2006) menciona que a função hormonal não está bem esclarecida, sabe-se que os hormônios ovarianos são essenciais para o desenvolvimento e funcionamento do sistema reprodutivo das aves.
O objetivo da pesquisa foi verificar a concentração de proteínas, em especial a fosvitina/lipovitelina presente em gemas de ovos crus e cozidos nos diferentes ciclos de postura (inicial, intermediário e final) e avaliar a interferência desta proteína na disponibilidade do micronutriente ferro.
2 Material e métodos
2.1 Matéria-prima
Foram utilizados 270 ovos de galinhas da raça Bovans, provenientes de uma granja situada na cidade de Cordeirópolis, SP. Os ovos foram coletados de um único lote de 200 m com aproximadamente nove mil poedeiras. A coleta foi realizada em diferentes períodos de idade, no início, meio e fim de postura (18, 40 e 75 semanas respectivamente), sendo coletados 90 ovos por época de postura. A primeira coleta se deu em janeiro de 2006, a segunda em junho de 2006 e a terceira em dezembro de 2006. A alimentação oferecida às galinhas foi ração comercial (Tabelas 1 e 2), em diferentes proporções de nutrientes, conforme a idade e produção de cada uma delas.
Os tratamentos em gemas cruas e cozidas foram realizados em diferentes períodos.
Tratamento 1: gemas cruas de poedeiras em início de postura;
Tratamento 2: gemas cruas de poedeiras em idade média de postura;
Tratamento 3: gemas cruas de poedeiras em final de postura;
Tratamento 4: gemas cozidas de poedeiras em início de postura;
Tratamento 5: gemas cozidas de poedeiras em idade média de postura; e
Tratamento 6: gemas cozidas de poedeiras em final de postura.
Para a cocção nos tratamentos 4, 5 e 6, os ovos utilizados foram colocados em água à temperatura ambiente em panela de alumínio e permaneceram por 15 minutos em água fervente, considerados a partir do início da fervura da água. Imediatamente após a cocção foram descascados e preparados para a realização das análises.
2.2 Composição centesimal e teor de ferro
As análises químicas da matéria seca, cinza, extrato etéreo e proteínas foram realizadas de acordo com a metodologia descrita pela Associação Oficial de Química Analítica, AOAC (2006). O teor de ferro foi quantificado segundo Sarruge e Haag (1974).
2.3 Diálise de ferro in vitro
A análise foi realizada pelo método descrito por Whittaker, Fox e Forbes (1989), utilizando pepsina e pancreatina bile para a digestão das amostras. A diálise foi realizada colocando o material digerido em sacos de diálise (25 Angstrons). Foi realizada a leitura a 533 nm em espectrofotômetro Beckman modelo DU 640. A quantidade de ferro dialisado foi obtida por meio de curva padrão previamente preparada. Os resultados foram expressos em porcentagem.
2.4 Extração da fosvitina
A fosvitina foi isolada de acordo com o método de McBee e Cotterill (1979). As gemas, após quebra manual dos ovos frescos, foram cuidadosamente separadas da clara e calaza com o auxílio de papel filtro. A membrana vitelina foi perfurada com uma agulha e o conteúdo coletado em um béquer à temperatura de 4 °C. As gemas foram homogeneizadas com NaCl 0,17 M e então centrifugadas (J2-MC Centrifuge) por 45 minutos a 10.000 g. Coletaram-se os grânulos que misturados em volume de 2:1 mL de solução de NaCl 0,17 M, e essa mistura foi então centrifugada novamente. Os grânulos lavados foram misturados em solução de 0,8 M de NaCl (1:10 mL). Em seguida, foi realizada diálise durante toda a noite. Após a diálise, a suspensão foi então centrifugada a 20.000 g por 30 minutos. Ao sobrenadante obtido contendo fosvitina foi adicionada acetona para a retirada de lipídios, obtendo-se amostras com massa de 6, 4,5 e 3,5 g (gemas de poedeiras em início, meio e fim de postura). Em seguida as amostras foram liofilizadas, obtendo-se então uma massa de 4,5, 3 e 2 g, respectivamente.
Todo o procedimento acima foi realizado em gemas cruas e em gemas cozidas.
2.5 Eletroforese em gel de poliacrilamida
Foi coletado 1 g de amostra liofilizada e homogeneizado em 300 µL de água mili-Q. Depois de quantificadas, as amostras foram submetidas à eletroforese em gel de poliacrilamida (SDS-PAGE procedimento de preparação e corrida dos géis segundo Laemmli (1970)). O sistema foi constituído de 2 géis, um de resolução (gel principal) e um de empacotamento (gel de empacotamento), ambos contendo acrilamida com uma concentração de 12 e 4%, respectivamente. Para a preparação das amostras, adicionou-se tampão de amostra, o qual continha 3,0 mL de água destilada, 1,0 mL de solução Tris-HCl (pH 6,7), 1,6 mL de glicerol, 1,6 mL de SDS 10%, 0,4 mL de β-mercaptoetanol e 0,4 mL da solução de 0,5% de azul de bromofenol. A proporção utilizada de amostra e tampão foi de 1:1 (v/v).
Depois de preparadas, as amostras foram fervidas durante 5 minutos, antes de serem colocadas no gel. Como marcador de massa molecular foi utilizado o BenchMarkTM Protein Ladder (Invitrogen). A eletroforese foi conduzida em sistema vertical à corrente constante de 15 mA e à temperatura ambiente, por cerca de 3,5 a 4 horas.
A coloração dos géis foi realizada de acordo com a quantidade de proteína aplicada no gel, com reagente de Coomassie (para 20 µg de proteína) ou Coloração com prata (para 5 µg de proteína).
Para a coloração do gel com Coomassie, foi utilizada solução de 0,5 g de Coomassie azul brilhante R-250 em 500 mL de uma solução 20% de metanol, 8% de ácido acético e 76% de água destilada overnight. Para descorar o gel, utilizou-se solução 20% de metanol, 8% de ácido acético e 76% de água destilada, durante toda a noite.
2.6 Delineamento estatístico
O delineamento estatístico foi inteiramente ao acaso com comparação das médias obtidas nos diferentes tratamentos. Foram analisados 6 tratamentos em ovos obtidos em 3 tempos e 2 formas. Os tratamentos foram analisados pelo teste F e posteriormente pelo método de Tukey (5%) utilizando o software Statistical Analysis System (1999).
3 Resultados e discussão
Os valores de umidade, extrato etéreo, proteínas, cinza e carboidratos presentes em gemas de poedeiras em postura inicial, intermediária e final para gemas cruas e cozidas estão apresentados na Tabela 3.
Tratando-se de gemas cruas e gemas cozidas foi possível perceber diferenças significativas em alguns tratamentos, porém de modo geral observou-se que não houve diferença (p < 0,05) significativa entre os tratamentos.
Em relação aos valores de umidade, estes apresentaram-se inferiores em relação ao valor apresentado pelo IBGE (1999). Quando se considera o ovo inteiro, Franco (1999) apresentou valor superior.
Quando comparadas gemas cruas e cozidas de ovos de poedeiras em início, meio e fim de postura, foi possível observar que as diferenças foram pequenas, sendo significativas apenas no tempo de postura intermediária, com teor de umidade mais elevado em gemas que passaram pelo processo de cocção. Já entre os tempos de postura inicial e final não houve diferença significativa, indicando que a cocção não alterou o teor de umidade da gema.
Segundo Ordónez (2005); Souza-Soares e Siewerdt (2005), a gema adquire água do albúmen durante o período de armazenamento de ovos, portanto o seu conteúdo em umidade pode variar de 46 a 59%, dependendo do tempo e condições de armazenamento. Durante o experimento não houve a influência do armazenamento, já que os ovos foram coletados e analisados em seguida.
Em relação à idade das poedeiras, notou-se que poedeiras em postura inicial apresentaram gemas com teor de umidade mais elevado, porém em gemas cruas. Já em gemas cozidas os resultados foram diferenciados nos três ciclos de postura.
Quanto aos resultados de extrato etéreo, constatou-se que foram semelhantes aos encontrados na literatura (FRANCO, 1999; IBGE, 1999; FARIA, 2002). Para o ovo integral os valores foram mais baixos segundo Fennema (1993). Houve diferença entre gemas cruas e cozidas, mostrando que o teor de extrato etéreo foi mais elevado em gemas cozidas de poedeiras em postura inicial e intermediária; já em gemas de poedeiras no tempo final de postura, os valores de extrato etéreo foram semelhantes nos dois tratamentos (gemas cruas e cozidas). Na análise dos resultados relacionados à idade das poedeiras, foi observado que: em gemas cruas o teor de extrato etéreo aumentou conforme o envelhecimento da poedeira; em gemas cozidas o teor de lipídios foi menor em gemas de poedeiras que estavam em início de postura; e nos ciclos intermediário e final os resultados foram semelhantes, porém mais elevados que no ciclo inicial.
Os valores de proteínas foram semelhantes aos registrados na literatura (IBGE, 1999; FRANCO, 1999; FARIA, 2002; FENNEMA, 1993). Não ocorreu diferença significativa entre gemas cruas e cozidas, mas sim em gemas de diferentes fases de produção, como pode ser constatado nos dados apresentados na Tabela 1.
O teor de cinza nas amostras foi concordante aos apresentados pela literatura (IBGE, 1999; FARIA, 2002). Já Fennema (1993) apresentou valor inferior aos mostrados na tabela, considerando o ovo inteiro. Ocorreu diferença dos valores de cinza quando comparadas gemas cruas e cozidas, isto em poedeiras de postura intermediária, mostrando um teor mais elevado em gemas cozidas, já em postura inicial e final a diferença entre os resultados não foi significativa.
Quando comparados os resultados de lipídios e proteínas da Tabela 3, foi observada diferença entre os três ciclos de postura, visto que quanto mais elevada a quantidade de nutrientes na ração alimentar de poedeiras mais jovens, menor o teor de lipídios e proteínas em gemas. Isso está associado aos resultados encontrados por Ângelo (2007), que menciona que poedeiras mais jovens apresentam dificuldades em digerir e absorver certos nutrientes, tendo a anatomia e fisiologia do aparelho digestivo diferenciada das aves em estágios mais adiantados de vida, o que influencia na deposição desses elementos na gema.
A quantidade de ferro total apresentada na Tabela 4 foi concordante com achados da USDA (2001), Franco (1999) e IBGE (1999), visto que a gema ao passar pelo processo de cocção sofreu diminuição da quantidade de ferro total, considerando que a literatura aponta valores em ovos cozidos e íntegros e não somente na gema.
Observou-se que no tempo inicial a quantidade de ferro diminuiu com a cocção, o mesmo ocorreu com as gemas na postura intermediária, já na postura final os resultados obtidos mantiveram-se estáveis em gemas cruas e cozidas.
Tratando-se dos três períodos de produção das poedeiras, constatou-se que do ciclo inicial ao ciclo final a quantidade de ferro total foi diminuindo sucessivamente.
Na análise dos resultados de ferro dialisável, observou-se que os três períodos (inicial, intermediário e final) (Tabela 5) apresentaram-se inferiores aos mencionados por Machado e Canniatti-Brazaca (2006) e Harder et al. (2007), que utilizaram urucum na composição da ração oferecida às poedeiras, contribuindo assim para o aumento da disponibilidade do ferro, sendo o urucum rico em carotenoides.
Ao se analisarem gemas cruas e cozidas, constatou-se que a quantidade de ferro dialisável em gemas cozidas do período inicial foi maior em relação a gemas cruas, no período intermediário os resultados foram semelhantes aos do período inicial e final.
Em relação aos três períodos foi possível notar que quanto mais velha a poedeira maior a quantidade de ferro disponível, visto que nos três períodos a quantidade de ferro foi aumentando sucessivamente, porém o último período se mostrou com um resultado mais satisfatório. O ferro disponível está correlacionado com a quantidade proteica presente em gemas, pois se liga à proteína fosvitina, ficando indisponível para ser absorvido (JIANG; MINE, 2000). Segundo Anton, Le Denmat, Gander (2000), a fosvitina resiste a altas temperaturas, porém os resultados obtidos não condizem com a literatura, acredita-se que este fato possa ser resultante do tempo de cocção empregado.
Wallace e Selman (1981); Matsubara e Sawano (1995) determinam que a síntese de fosvitina ocorre em resposta a hormônios ovarianos, ou seja, Rankouhi et al. (2002) citam que é sintetizada em consequência de estrógenos-dependentes, gene expresso em hepatócitos de poedeiras fêmeas e de machos. Estes hormônios estão diretamente ligados à idade das poedeiras. Elas atingem maturidade sexual entre 17 e 18 semanas, atingindo o pico de postura entre 28 e 35 semanas, que se reduz gradativamente após este período. Isto significa que os hormônios vão se alterando de acordo com o ciclo de vida da poedeira (TAKATA et al., 2001). Estes hormônios (estrogênio, progesterona e androgênio) são essenciais para o desenvolvimento e funcionamento do sistema reprodutivo das aves (MORAES, 2006), e podem afetar a produção de diferentes proteínas.
Os resultados obtidos (Tabela 6) mostraram que gemas cozidas de poedeiras em ciclos intermediário e final apresentaram baixo teor de proteínas, após a extração, quando comparados com gemas que não passaram pelo processo de cocção. Nas gemas obtidas de ovos de postura inicial e cozidas não foram extraídas proteínas. Ao comparar as gemas em relação aos três períodos de produção, foi observado que quanto mais velhas as poedeiras menor o teor proteico extraído, tanto em gemas cruas como em gemas cozidas.
A proteína fosvitina apresenta poucos aminoácidos básicos e aromáticos que podem ser detectados por absorbância UV ou por eletroforese (WALLACE, 1963).
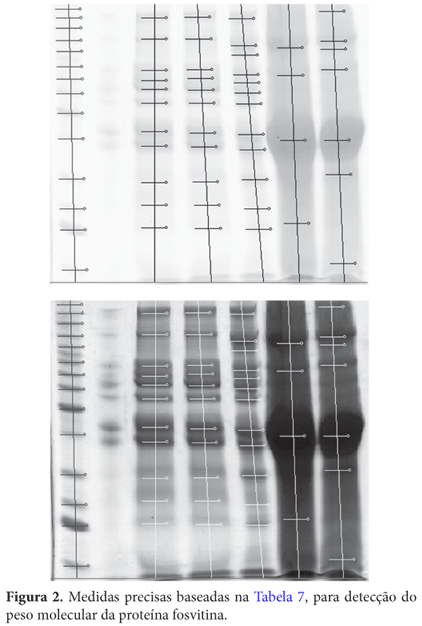
Segundo Jiang e Mine (2000), a fosvitina também denominada de fosfoproteína apresenta massa molecular de 36 kDa (Figura 1), contendo aproximadamente 10% de fósforo e 6,5% de carboidratos.
Comparando-se ao padrão, pode-se observar que esta proteína extraída de gemas cruas esteve presente nos três períodos de postura (inicial, intermediário e final). Já em gemas que passaram pelo processo de cocção, a percepção não foi satisfatória, devido à dificuldade de precipitação proteica.
Através do Programa Kodak Digital Science 1D versão 3.01 (Tabela 7), foi constatado que a banda apontada (Figuras 1 e 2) apresentou 38 kDa, porém bastante próximo de 36 kDa como determina Jiang e Mine (2000).
4 Conclusões
Houve diferença na concentração de proteínas e diferentes níveis de ferro dialisável nos três ciclos de postura, para gemas cruas e cozidas.
Através deste estudo, concluiu-se que é interessante consumir ovos de poedeiras com idade mais avançada, visto que com o passar dos ciclos (inicial, intermediário e final), as poedeiras mais velhas, apresentaram ovos (gemas) com teor mais elevado de ferro disponível, porém com quantidade inferior de proteínas em especial a fosvitina/lipovitelina, comparando-se com os ciclos inicial e intermediário.
Em relação às análises de gemas cruas e cozidas, pode-se concluir que o teor de ferro dialisável foi maior em gemas cozidas em poedeiras de postura final.
Agradecimentos
Ao Prof. Dr. Ricardo Antunes de Azevedo do Departamento de Genética da ESALQ/USP pelo uso do laboratório, às Granjas Küller (Cordeirópolis SP) pela doação dos ovos e informações essenciais para o desenvolvimento da pesquisa e à FAPESP pelo auxílio financeiro.
Recebido para publicação em 23/7/2007
Aceito para publicação em 3/1/2009 (002699)
- ÂNGELO, J. C. Benefícios da nutrição de aves na primeira semana. Disponível em:<http://www.guabi.com.br/rc/aves_postura>. Acesso em: 25 janeiro 2007.
- ANTON, M.; LE DENMAT, M.; GANDEMER, G. Thermostability of hen egg yolk granules: contribution of native structure of granules. Journal of food Science, v. 65, n. 4, p. 581-584, 2000.
- ASOCIACION de Productores de Huevos de Chile. Disponível em:<http://www.asohuevo.cl>. Acesso em: 20 abril 2005.
» link - ASSOCIATION of Official Analytical Chemists AOAC. Official methods of analysis. 16 ed. Washington, 2006.
- BERTECHINI, A. G. Mitos e verdades sobre o ovo de consumo. Disponível em:<http://www.avisite.com.br>. Acesso em: 10 março 2005.
- ENSMINGER, M. E. Poultry Science, Champaign. 3 ed. Illinois: Interstate Publishers, 1992. 469 p.
- FARIA, D. E. Suplementação de vitaminas D e C para poedeiras durante o primeiro ciclo de produção. Revista Brasileira de Ciência Avícola, v. 1, n. 2, p.135-144, 2002.
- FENNEMA, O. R. Química de los Alimentos. Zaragoza: Acribia, 1993. 53 p.
- FRANCO, G. Tabela de composição química dos alimentos. 9 ed. São Paulo: Editora Atheneu, 1999.
- HARDER, M. N. C. et al. Cholesterol and iron availability in yolk of laying hens feed with annatto (Bixa orellana). Animal, v. 1, n. 3, p. 477-482, 2007.
- Instituto Brasileiro de Geografia e Estatística – IBGE. Disponível em:<http://www.ibge.gov.br>. Acesso em: 26 novembro 2006.
» link - JIANG, B.; MINE, Y. Preparation of novel functional oligophosphopeptides from hen egg yolk phosvitin. Journal Agriculture Food Chemical, v. 48, n. 4, p. 990-994, 2000.
- LAEMMLI, U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, v. 227, Aug, p. 680-685, 1970.
- MACHADO, F. M. V. F.; CANNIATTI-BRAZACA, S. G. Avaliação da disponibilidade de ferro em ovo, cenoura e couve e em suas misturas. Ciência e Tecnologia de Alimentos, v. 26, n. 3, p. 610-618, jul/set. 2006.
- MATSUBARA, T.; SAWANO, K. Proteolytic Cleavage of Vitellogenin and Yolk Proteins During Vitellogenin Uptake and Oocyte Maturation in Barfin Flounder (Verasper moseri). Journal Experimental Zoology, v. 272, n. 1, p. 34-35, 1995.
- MCBEE, L. E.; COTTERILL, O. J. Ion exchange chromatography and electrophoresis of egg yolk proteins. Journal Food Science, v. 44, n. 3, p. 656-660, 1979.
- MELO, M. R. et al. Uso de índices hematimétricos no diagnóstico diferencial de anemias microcíticas: uma abordagem a ser adotada?. Revista Associação de Medicina Brasileira, v. 48, n. 3, p. 222-224, 2002.
- MINISTÉRIO DA SAÚDE. Departamento de atenção básica. Disponível em:<http://dtr2004.saude.gov.br>. Acesso em: 23 janeiro 2007a.
» link - MINISTÉRIO DA SAÚDE. Departamento de atenção básica. Disponível em:<http://dtr2005.saude.gov.br>. Acesso em: 23 janeiro 2007b.
» link - MORAES, I. A. Fisiologia da reprodução das aves domésticas. Disponível em:<http://www.uff.br/fisiovet/fisio_rep_aves.htm>. Acesso em: 05 março 2006.
- ORDÓNEZ, J. A. Ovos e produtos derivados. In: Tecnologia de alimentos: Alimentos de origem animal. Porto Alegre: Artmed, 2005. p. 269-279.
- RANKOUHI, T. R. et al. Effects of primary exposure to environmental and natural estrogens on vitellogenin production in carp (Cyprinus carpio) hepatocytes. Toxicological Sciences, v. 67, n. 1, p. 75-80, 2002.
- SARRUGE, J. R.; HAAG, H. P. Análises química em plantas. Piracicaba: ESALQ, Depto Química, 1974. 50 p.
- SAS Institute. SAS user's guide: statistics. Version 8.0. Cary: SAS, 1999.
- SCHIRM, J.; GRUBER, M.; AB, G. Post-translational phosphorylation of phosvitin. Febs Letters, v. 30, 8 p, março 1973.
- SOUZA-SOARES, L. A.; SIEWERDT, F. Aves e ovos. Pelotas: Editora da Universidade UFPEL, 2005. 137 p.
- Institudo Brasileiro de Geografia e Estatistica – IBGE. Tabelas de Composição de Alimentos. Estudo Nacional da Despesa Familiar. 5 ed. Rio de Janeiro, 1999. 137 p.
- TAKATA, F. N. et al. Aspectos morfológicos do oviduto de galinha doméstica (Gallus gallus) antes e após a puberdade. Revista Brasileira de Reprodução Animal, v. 25, n. 2, p. 174-176, 2001.
- National Nutriente Database for Standard Reference USDA. Release 17. United States, 2001. Disponível em:<http://www.nal.usda.gov/fnic/foodcomp/cgi-bin/list_nut_edit.p1>. Acesso em: 10 junho 2006.
- WALLACE, R. A. Studies on amphibian yolk 4. An analysis of the main body component of yolk platelets. Biochemical Biophysical Acta, v. 74, p. 505-518, 1963.
- WALLACE, R. A.; SELMAN, K. Cellular and dynamic aspects of oocyte growth in Teleosts. American Zoology, v. 21, n. 2, p. 325-343, 1981.
- WHITTAKER, P.; FOX, M. R. S.; FORBES, A. L. In vitro prediction of iron bioavailability for food fortification. Nutrition Reports International, v. 39, n. 6, p. 1205-1215, 1989.
- YASHODA, K. P. et al. Egg loaf and changes in its quality during storage. Food Control, v. 15, p. 523-526, 2004.
Datas de Publicação
-
Publicação nesta coleção
04 Nov 2009 -
Data do Fascículo
Set 2009
Histórico
-
Aceito
03 Jan 2009 -
Recebido
23 Jul 2007