Resumos
O trabalho teve como objetivo avaliar a influência dos compostos nitrogenados na concentração de proteína da cianobactéria Aphanothece microscopica Nägeli quando cultivada em meio de cultivo padrão BG11 e no efluente do processamento de pescado. As condições experimentais foram inóculo de 200 mg.L-1, 30 °C, pH 7,6 e razão C/N 20. As amostras foram retiradas no início, meio e fim da fase logarítmica e da fase estacionária e analisadas quanto à concentração de nitrogênio total, nitrogênio não proteico, amônio, nitrato, nitrito e ácidos nucleicos. Ficou demonstrado em todas as culturas que o íon amônio, juntamente com os ácidos nucleicos, representa uma importante fração do nitrogênio não proteico presente na biomassa. Os resultados mostraram que o meio de cultivo influencia a concentração de nitrogênio e que a determinação de proteína pelo método de Kjeldahl superestima a concentração proteica em cianobactérias.
meio de cultivo; nitrogênio; cianobactéria
This study aimed to evaluate the influence of nitrogen compounds on the protein concentration of the cyanobacterium Aphanothece microscopica Nägeli cultivated in BG11 medium and fish processing effluent. The experimental conditions were 200 mg L-1 inoculum, 30 °C, pH 7.6, and C/N ratio 20. Each culture was harvested during the early, mid-log, and late logarithmic growth phase and during the stationary phase and analyzed in terms of total nitrogen, non-protein nitrogen, ammonium nitrogen, nitrate, nitrite, and nucleic acid. It can be said that in all cultures the ammonium ion and the nucleic acids account for great portion of the non-protein nitrogen in the biomass. The results show that the culture medium influences the nitrogen content, and that the Kjeldahl method overestimates the real protein concentration in cyanobacteria.
culture medium; nitrogen; cyanobacteria
ORIGINAL
Influência dos compostos nitrogenados na concentração de proteína da cianobactéria Aphanothece microscopica Nägeli
Influence of nitrogen compounds on the protein concentration of the cyanobacterium Aphanothece microscopica Nägeli
Marcio Hornes* * A quem a correspondência deve ser enviada ; Adriana Gonçalves Silva; Marina Leite Mitterer; Maria Isabel Queiroz
Laboratório de Biotecnologia, Departamento de Química, Universidade Federal do Rio Grande - FURG, Rua Eng. Alfredo Huch, 475, CEP 96201-900, Rio Grande - RS, Brasil, E-mail: biotecnofurg@yahoo.com.br
RESUMO
O trabalho teve como objetivo avaliar a influência dos compostos nitrogenados na concentração de proteína da cianobactéria Aphanothece microscopica Nägeli quando cultivada em meio de cultivo padrão BG11 e no efluente do processamento de pescado. As condições experimentais foram inóculo de 200 mg.L-1, 30 °C, pH 7,6 e razão C/N 20. As amostras foram retiradas no início, meio e fim da fase logarítmica e da fase estacionária e analisadas quanto à concentração de nitrogênio total, nitrogênio não proteico, amônio, nitrato, nitrito e ácidos nucleicos. Ficou demonstrado em todas as culturas que o íon amônio, juntamente com os ácidos nucleicos, representa uma importante fração do nitrogênio não proteico presente na biomassa. Os resultados mostraram que o meio de cultivo influencia a concentração de nitrogênio e que a determinação de proteína pelo método de Kjeldahl superestima a concentração proteica em cianobactérias.
Palavras-chave: meio de cultivo; nitrogênio; cianobactéria.
ABSTRACT
This study aimed to evaluate the influence of nitrogen compounds on the protein concentration of the cyanobacterium Aphanothece microscopica Nägeli cultivated in BG11 medium and fish processing effluent. The experimental conditions were 200 mg L-1 inoculum, 30 °C, pH 7.6, and C/N ratio 20. Each culture was harvested during the early, mid-log, and late logarithmic growth phase and during the stationary phase and analyzed in terms of total nitrogen, non-protein nitrogen, ammonium nitrogen, nitrate, nitrite, and nucleic acid. It can be said that in all cultures the ammonium ion and the nucleic acids account for great portion of the non-protein nitrogen in the biomass. The results show that the culture medium influences the nitrogen content, and that the Kjeldahl method overestimates the real protein concentration in cyanobacteria.
Keywords: culture medium; nitrogen; cyanobacteria.
1 Introdução
Cianobactérias ou algas verdes-azuis são microrganismos procariontes largamente distribuídos na natureza que possuem a capacidade de realizar fotossíntese como as plantas superiores (SÁNCHEZ-LUNA et al., 2004). Entretanto, a despeito de sua natureza fotossintética aeróbica, algumas espécies de cianobactérias podem crescer no escuro, utilizando substratos orgânicos (STAL; MOEZELAAR, 1997; QUEIROZ et al., 2007). As cianobactérias, além de várias espécies de microalgas, têm sido consideradas organismos promissores para várias aplicações biotecnológicas e são alvo de estudos devido ao seu potencial para diversas formas renováveis de compostos químicos úteis, tanto para alimentação humana como animal, além de produtos farmacêuticos, biofertilizantes, entre outras aplicações (DE PHILIPPIS; VINCENZINI, 1998; SHI et al., 2000; BURJA et al, 2001; MOLINA-GRIMA et al., 2002; PATEL et al.; 2005; ZEPKA et al., 2008).
O efeito de mudanças nas condições ambientais em culturas de muitas espécies de cianobactérias e microalgas tem sido estudado para um melhor entendimento da sua fisiologia e para responder questões específicas no que se refere ao crescimento das culturas. A ação de alguns nutrientes, especialmente o nitrogênio, permite obter resultados sobre algumas tendências gerais de culturas em batelada (WITJITRA; SHAH; CHERYAN, 1996; LOURENÇO et al., 1997; SHI et al., 2000).
O nitrogênio é um nutriente que pode limitar o crescimento microbiológico em ambientes naturais ou artificiais. Íons nitrato, nitrito e amônio e muitos compostos nitrogenados orgânicos dissolvidos (ureia, aminoácidos e peptídeos) são as principais fontes de nitrogênio, e são utilizadas principalmente na síntese de aminoácidos e proteínas (REYNOLDS, 1984; LUQUE; FLORES; HERRERO, 1994; MURO-PASTOR; FLORÊNCIO, 2003). De acordo com Rajoka et al. (2006), a utilização de diferentes fontes de nitrogênio no meio causa uma mudança na síntese de proteínas e pigmentos. A disponibilidade do íon amônio interfere diretamente na assimilação de outras fontes de nitrogênio. Por isso, em um meio em que há várias fontes de nitrogênio, inicialmente são selecionadas as formas mais reduzidas, como aminoácidos, ureia e amônio (HERRERO;MURO-PASTOR; FLORE, 2001; MURO-PASTOR; FLORÊNCIO, 2003). Von Rückert e Giani (2004) realizaram experimentos utilizando um meio com fonte de nitrogênio amônio e nitrato nas concentrações de 3450 e 3500 μg.L-1, respectivamente. Os resultados demonstraram que o íon amônio foi removido completamente no 10º dia, enquanto que o íon nitrato permanecia ainda disponível.
A composição química das cianobactérias pode variar com a idade da cultura e com mudanças nas condições ambientais. A manipulação destes fatores pode alterar as características de crescimento e a composição química das células cultivadas nas condições aplicadas, e qualquer variabilidade associada com a fase de crescimento pode incluir também mudanças associadas à concentração de nutrientes (LOURENÇO et al., 1997; FIDALGO et al., 1998).
A cianobactéria Aphanothece microscopica Nägeli apresenta ocorrência natural nos corpos hídricos adjacentes à cidade do Rio Grande, e sua utilização em tratamento de efluentes de indústria de alimentos vem sendo estudada pela Fundação Universidade Federal do Rio Grande, obtendo ótimos resultados na remoção de matéria orgânica e nutrientes (QUEIROZ et al., 2002; HORNES; QUEIROZ, 2004; QUEIROZ et al., 2004; QUEIROZ et al., 2007). No entanto, no que tange a avaliação química em diferentes meios de cultivo, pouca informação até o momento está disponível. Em face disso, o trabalho teve como objetivo avaliar a influência dos compostos nitrogenados na concentração de proteína da cianobactéria Aphanothece microscopica Nägeli quando cultivada em meio de cultivo padrão BG11 e no efluente do processamento de pescado.
2 Material e métodos
2.1 Microrganismo
Suspensões de Aphanothece microscopica Nägeli (RSMan92, Unidade de Cianobactérias da Universidade Federal do Rio Grande), isoladas do estuário da cidade de Rio Grande, RS, Brasil, foram cultivadas fotoautotroficamente a 2 Klux e fotoperíodo de 12 horas. A luminosidade foi provida por uma série de três lâmpadas fluorescentes (40 W). As demais condições foram 30 °C, pH 7,8 - 8,0, em frascos de 4000 mL contendo meio padrão BG11: K2HPO4.3H2O (0,04 g.L-1), MgSO4.7H2O (0,075 g.L-1), Na2CO3 (0,15 g.L-1), NaNO3 (0,15 g.L-1), EDTA (0.001 g.L-1), H3BO3 (2,86 g.L-1), MnCl2.4H2O (1.81 g.L-1), ZnSO4.7H2O (0,222 g.L-1), Na2MoO4.2H2O (0,39 g.L-1), CuSO4.5H2O (0,079 g.L-1), CoCl2.6H2O (0,040 g.L-1), ácido cítrico (0,006 g.L-1) e citrato férrico de amônio (0,006 g.L-1) (RIPKA et al., 1979).
As células na fase exponencial de crescimento foram separadas do meio padrão BG11 por centrifugação a 4000 g/20 minutos e, em seguida, lavadas com água destilada. Para todos os experimentos, a concentração inicial de inóculo foi de 200 mg.L-1.
2.2 Meio de cultivo
Como meios de cultivo, foram utilizados o efluente do processamento de pescado e meio padrão BG11 previamente esterilizados. O efluente proveniente do tanque de equalização do sistema de tratamento de efluentes de uma indústria de pescado local foi coletado em recipientes de polietileno e estocado a 4 °C. A Tabela 1 mostra a caracterização do efluente coletado em uma indústria processadora de pescado local, cujos dados refletem o resultado de dez amostragens por um período de seis meses. O meio BG11 foi preparado de acordo com o item 2.1. Os meios de cultivo foram esterilizados a 1,1 kgf.cm-2 (120 °C) por 15 minutos. A escolha de diferentes condições de cultivo, no que se refere à iluminação do meio, foi atribuída ao fato de que, no efluente, devido à alta concentração de matéria orgânica, a ausência de luz conduz ao metabolismo respiratório, comprovadamente eficiente no crescimento e produção de componentes celulares de cianobactéria (QUEIROZ et al., 2004; QUEIROZ et al., 2007; ZEPKA et al., 2008).
2.3 Sistema experimental
Os experimentos foram realizados em sistema de batelada a 30 °C e aeração de 1 vvm (L.L-1 minuto-1). Os valores iniciais de pH nas culturas foram ajustados a 7,6 antes da inoculação. Os tratamentos foram feitos em duplicada e os resultados, apresentados como valores médios.
A cianobactéria Aphanothece microscopica Nägeli foi cultivada no efluente do processamento de pescado na ausência de luz por 60 horas. Foi utilizado um biorreator com diâmetro interno de 0,10 m e altura de 0,8 m, tendo um volume operacional de 4,0 L. A razão C/N no meio de cultura foi 20, sendo que, como fontes de carbono e nitrogênio para eventuais ajustes, foram usados glicose e cloreto de amônio, respectivamente. As células no meio BG11 foram cultivadas em frascos contendo 4,0 L de meio por 280 horas e iluminação de 2 Klux em ciclos claro-escuro de 12 horas. As amostras foram retiradas em intervalos regulares para medir as concentrações de biomassa celular seca. A biomassa foi separada dos meios de cultivo por filtração em filtro Millipore com porosidade de 0,45 μm e seca em estufa a 60 °C até peso constante.
Os parâmetros cinéticos determinados foram velocidade específica de crescimento máxima (μmax) e tempo de geração. A velocidade específica de crescimento máxima foi calculada pela relação linear das curvas de crescimento na fase exponencial, onde μmax corresponde ao coeficiente "x" na equação linear e "y" é o logaritmo neperiano da concentração celular (von RÜCKERT; GIANI, 2004).
2.4 Análise dos compostos nitrogenados
As amostras das culturas no início, meio e fim da fase logarítmica e da fase estacionária foram separadas do meio de cultivo por centrifugação a 4000 × g por 15 minutos e filtradas em filtro Millipore (porosidade de 0,45 μm). As células foram secas a 60 °C até peso constante para estimar a biomassa seca.
Para as frações de nitrito, nitrato e amônio, o filtro Milipore (porosidade de 0,45 μm) contendo a biomassa foi imerso em água destilada sob refrigeração a 5 °C por 15 horas. Após esse período, o material foi filtrado por sistema a vácuo e lavado com água a 80 °C até atingir um volume aproximado de 50 mL. Os íons amônio foram determinados após adição de solução de fenol e nitroprussiato de sódio na presença de excesso de hipoclorito de sódio em meio alcalino, formando azul de indofenol absorvido em 640 nm (APHA, 2000). A determinação dos íons nitrito foi realizada pela reação com a sulfanilamida em meio ácido, formando um íon diazoico, o qual reage com N-naftil etilenodiamina, resultando em coloração rósea, cuja densidade ótica foi medida em 543 nm (APHA, 2000). Os íons nitrato foram determinados pela reação com o ácido fenoldisulfônico, resultando em um complexo colorido que pode ser avaliado por espectrofotometria (λ = 410 nm) (APHA, 2000).
O nitrogênio não proteico total foi determinado através do sobrenadante obtido após a precipitação das proteínas com ácido tricloroacético (TCA) 5% (AOAC, 2000).
Os ácidos nucleicos presentes na biomassa da cianobactéria Aphanothece foram extraídos através do método da Extração DNA/RNA com Fenol/Clorofórmio, conforme descrito por Sambroc (1989), e quantificados espectrofotometricamente (λ = 260 nm).
Para a determinação do nitrogênio total Kjeldahl (N-NTK), a biomassa retida em filtro Millipore (porosidade de 0,45 μm) foi seca em estufa a 60 °C, digerida em ácido sulfúrico e, então, destilada em meio alcalino. O destilado foi recolhido em solução de ácido bórico e titulado com ácido clorídrico de título conhecido. O teor de proteína foi calculado a partir da concentração de nitrogênio total mediante a utilização do fator 6,25 (AOAC, 2000).
3 Resultados e discussão
3.1 Crescimento
As Figuras 1 e 2 apresentam as curvas de crescimento da cianobactéria Aphanothece microscopica Nägeli no efluente do processamento de pescado e no meio padrão BG11, respectivamente. O comportamento das curvas sugerem duplicação celular em 23 horas com uma velocidade específica de crescimento máxima de 0,032 horas-1 para o efluente do processamento de pescado, enquanto que, para o meio BG11, a duplicação ocorreu em 41 horas com uma velocidade específica de crescimento máxima de 0,017 horas-1, conforme pode ser verificado na Tabela 2. O crescimento acelerado destes microrganismos em condições naturais e artificiais é constatado em lagos e reservatórios, causando problemas quando em altas densidades, como fortes odores e depleção do oxigênio dissolvido na água (HAVENS et al., 2003), e em estudos realizados em reatores em escala laboratorial e piloto em meios sintéticos e efluentes de indústrias de alimentos (DE LA NOÜE; LALIBERTÉ; PROULX, 1992; LEE; KIM, 2001; QUEIROZ et al., 2004; JACOB-LOPES et al., 2007; QUEIROZ et al., 2007; ZEPKA et al., 2008).
O crescimento de uma população microbiológica é afetado pela disponibilidade de nutrientes essenciais. A concentração inicial de um dado nutriente pode comprometer tanto a taxa do crescimento como o tamanho final da população. Nesse sentido, a quantidade de biomassa que pode ser sintetizada a partir de uma fonte de energia está correlacionada à quantidade de energia capturada pelo catabolismo celular. Considerando a influência das diferentes condições utilizadas, como luminosidade e composição do meio de cultivo, um fator importante que pode ter contribuído para o rápido crescimento da cianobactéria Aphanothece no efluente, em comparação ao meio padrão BG11, é a presença de diferentes fontes de nitrogênio nos meios em estudo. Sabe-se que o nitrogênio compõe cerca de 10% do peso seco de cianobactérias e é o elemento essencial mais requerido pelas células para a síntese de aminoácidos, proteínas, ácidos nucleicos, clorofila e outros pigmentos acessórios, como as ficobilinas (SAHA; UMA; SUBRAMANIAN, 2003). Enquanto que no meio padrão BG11 a fonte de nitrogênio disponível para as células provém do NaNO3, no efluente de pescado este componente aparece principalmente como compostos orgânicos e derivados inorgânicos como o íon amônio, que é a forma nitrogenada inorgânica mais facilmente assimilada (von RÜCKERT; GIANI, 2004). Hornes e Queiroz (2004) caracterizaram o efluente da indústria da pesca obtendo valores médios de nitrogênio amoniacal de 38,4 mg.L-1. Observa-se que a cianobactéria Aphanothece microscopica Nägeli pode alcançar uma densidade de células similar para o crescimento tanto no meio BG11 como no efluente (Figuras 1 e 2). Por outro lado, quando cresce no efluente, a concentração máxima de biomassa é alcançada mais rapidamente, registrando-se uma velocidade específica de crescimento máxima quase duas vezes maior quando comparado ao crescimento em meio BG11 (Tabela 2). Esse rápido crescimento é típico para a cianobactéria Aphanothece cultivada no efluente de indústria de alimentos, o que demonstra a capacidade destes microrganismos em removerem e assimilarem heterotroficamente os compostos orgânicos e inorgânicos presentes nos efluentes, conforme pode ser verificado nos trabalhos realizados por Hornes e Queiroz (2004), Zepka et al. (2008) e Queiroz et al. (2007), nos quais foi constatada a ausência da fase lag quando a cianobactéria foi inoculada no efluente do processamento de pescado e da parboilização do arroz, respectivamente.
3.2 Concentração das frações nitrogenadas na biomassa
Os diferentes meios de cultura, com composição de nutrientes e material carbonado provenientes de fontes orgânicas e inorgânicas, além do metabolismo respiratório e fotossintético utilizados nos experimentos, acarretaram perfis distintos quanto aos compostos nitrogenados gerados das culturas de Aphanothece.
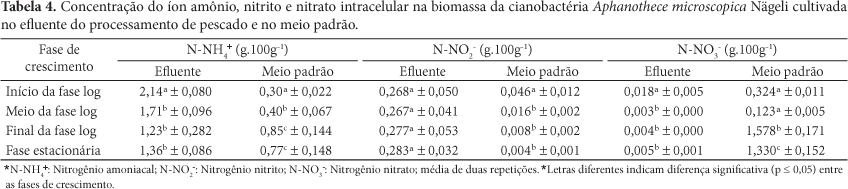
Analisando a Tabela 3, verifica-se que a concentração de N-NTK presente na biomassa cultivada no efluente do processamento de pescado apresentou no final da fase log e início da fase estacionária as maiores concentrações (10,67 e 11,61 mg N.g-1), valores estes que não diferem significativamente (p < 0,05). Na biomassa cultivada no meio padrão BG11, foram registrados maiores valores no início da fase logarítmica de crescimento (11,39 mg N.g-1), diminuindo no decorrer do cultivo. Comparando estes resultados com o nitrogênio amoniacal (Tabela 4), observa-se que também esta forma de composto nitrogenado se encontra em concentrações superiores na biomassa quando o crescimento é realizado no efluente em comparação ao meio BG11, demonstrando a influência do N-NH4+ na concentração de N-NTK e, consequentemente, de proteínas, quando esta é determinada por conversão de nitrogênio total.
A maior concentração de N-NTK e N-NH4+ intracelular na biomassa cultivada no efluente pode ser explicada pelo fato do meio BG11 conter basicamente NaNO3 como fonte de nitrogênio, enquanto que o efluente contém formas mais reduzidas e facilmente assimiladas, como N-NH4+, aminoácidos e ureia, provenientes da degradação das proteínas carreadas no afluente (ASPÉ; MARTÍ; ROECKEL, 1997; LIM; KHIM, HWANG, 2003). Muro-Pastor e Florêncio (2003) explicam que as cianobactérias são capazes de utilizar diferentes formas de nitrogênio, desde as moléculas mais oxidadas até as mais reduzidas. Contudo o uso de nitrato ou outras formas mais oxidadas requerem maior gasto energético no sentido de reduzir esses compostos e posteriormente assimilá-los nas moléculas biológicas. Isso quer dizer que íons amônio e formas reduzidas são mais facilmente assimiladas pelas cianobactérias com um esforço metabólico mínimo. De acordo com Herrero et al. (2001) e Luque et al. (2004), a maioria das cianobactérias desenvolveu um sistema para detecção do nitrogênio disponível na forma reduzida. Quando o íon amônio está disponível, todas as enzimas e transportadores envolvidos na assimilação de nitrogênio inorgânico são inibidos pelo NtcA, uma proteína transcricional repressora. Então, quando tanto o íon nitrato como o amônio estão presentes, este é geralmente preferido, embora a completa supressão da assimilação do nitrato seja rara (SYRETT; CRESWELL, 1979; GARCÍA-FERNÁNDEZ e DIEZ, 2004). A extensão dessa inibição varia de acordo com a concentração de amônio, o estado metabólico do microrganismo e variáveis ambientais como temperatura, pH e iluminação (von RÜCKERT; GIANI, 2004).
De acordo com Vonshak e Zeng (1998), a adição de nitrato em culturas de Spirulina representa aproximadamente 50% do custo total na produção em larga escala. Nesse sentido, Faintuch et al. (1992) relatam que um dos principais problemas na produção de proteína unicelular é o custo de compostos nitrogenados. Assim, um fator que merece destaque é a utilização de um meio de cultivo rico em nitrogenados no custo total em um sistema de cultivo de cianobactérias visando à produção de biomassa, que poderia ser utilizada como matéria-prima na fabricação de ração animal ou biofertilizante. A substituição por fontes baratas, como por exemplo, efluentes industriais, representa uma economia considerável no custo total da produção. De fato, em trabalhos realizados por Queiroz et al. (2007) e Zepka et al. (2007), foi utilizada a cianobactéria Aphanothece microscopica Nägeli na produção de proteína unicelular, tendo como meio de cultivo o efluente da parboilização de arroz. Estimou-se uma produção de 355,2 mg N.Kg-1 de arroz processado em um tempo de detenção hidráulica de 15 horas, concentração inicial de biomassa de 100 mg.L-1 e temperatura controlada de 30 °C.
No que se refere ao parâmetro NNP, as concentrações no meio BG11 foram significativamente maiores (p < 0,05) do que quando cultivada no efluente em todas as fases de crescimento. Da mesma forma, com exceção do meio da fase log, todas as concentrações de NNP, em função do N-NTK, foram superiores para o meio BG11. Esses dados mostram a importante contribuição dos compostos nitrogenados não proteicos para o nitrogênio total e, consequentemente, para a fração proteica. O aumento significativo do NNP na biomassa gerada no meio BG11 entre o meio e fim da fase logarítmica e da fase estacionária significa que houve máxima conversão do nitrogênio absorvido em ácidos nucleicos, que é o componente mais expressivo do NNP, além do possível acúmulo das frações de nitrato e amônio.
Observa-se na Tabela 4 que a maior concentração em nitrogênio amoniacal, quando a cianobactéria é cultivada no efluente, é registrada no início da fase log, o que não ocorreu no meio BG11, em que a concentração de N-NH4+ totalizou 29,3% do nitrogênio não proteico presente na biomassa no final da fase logarítmica. Lourenço (1997) reporta que, quando o íon N-NH4+ encontra-se em altas concentrações no meio intracelular de microalgas, gera alterações no metabolismo celular que culminam, em muitos casos, na redução ou interrupção do processo de crescimento. Isto justifica o maior tempo de geração (41 horas) para o crescimento do microrganismo em meio padrão quando comparado ao inoculado no efluente do processamento de pescado (23 horas), conforme podemos avaliar na Tabela 2, já que nitrato e nitrito devem ser enzimaticamente reduzidos a amônio antes da assimilação em moléculas biológicas, como aminoácidos e ácidos nucleicos. De acordo com Herrero, Muro-Pastor e Flores (2001) e García-Fernández e Diez (2004), isto pode ser claramente evidenciado, uma vez que as concentrações de nitrogênio como íon amônio são maiores no início e meio da fase exponencial e, conforme ocorre a depleção deste componente, verifica-se o aumento das concentrações intracelulares de nitrato e nitrito (Tabela 4), provavelmente devido à inibição da proteína NtcA e consequente absorção de outras formas nitrogenadas.
Nas cianobactérias, o íon amônio reprime o sistema de assimilação de nitrato, o que inclui o transporte deste e a posterior redução a amônio. Além disso, na presença de íons amônio há também a redução da síntese de glutamina sintetase e, em algumas espécies, pode até mesmo inativar a síntese desta enzima, a qual é responsável pela metabolização do amônio (HERRERO; FLORES; GUERRERO, 1981). Um efeito similar foi observado quando a microalga Chlorella protothecoides foi utilizada na presença de diferentes fontes de nitrogênio. A concentração máxima de biomassa nas culturas com nitrato e amônio foram 18,4 e 18,9 g.L-1 de células por dia, respectivamente (SHI; ZHANG; CHEN, 2000).
A Tabela 5 mostra as concentrações de ácidos nucleicos presentes na biomassa da cianobactéria Aphanothece. De acordo com Sgarbieri (1996), o rápido crescimento dos microrganismos em geral acarreta um maior acúmulo de ácidos nucleicos. Porém o cultivo da cianobactéria Aphanothece no efluente do pescado resultou em uma concentração menor em relação ao meio BG11. Um possível motivo que poderia contribuir para isso é a concentração de fosfatos presente nos meios de cultivo. Enquanto no efluente a concentração de P-PO4-3 apresenta um valor médio de 9,9 mg.L-1 (Tabela 1), no meio padrão BG11 a concentração é de 40 mg.L-1. Essa maior concentração de fosfatos no meio contribui para a síntese de DNA e RNA, além do aumento das reservas internas (GEIDER; LAROCHE, 2002). Quando comparados com outras cianobactérias e microalgas em diferentes condições de crescimento, os valores máximos e mínimos na biomassa cultivada no efluente do pescado foram inferiores (ORTEGA et al., 1993; FUENTES et al., 2000). Ortega et al. (1993) avaliaram o perfil bioquímico da cianobactéria Spirulina, que apresentou uma concentração de 4 a 6% de ácidos nucleicos. Da mesma forma, Fuentes et al. (2000) analisaram o perfil de nutrientes na biomassa da microalga Porphyridium cruentum submetida a diferentes tempos de residência e luminosidade, na temperatura de 20 °C e pH 7,6. Os autores encontraram uma concentração máxima de 2,36 g.100 g-1 de ácidos nucleicos.
Na Tabela 6 estão apresentadas as concentrações de proteína em relação ao nitrogênio total Kjeldahl (N-NTK) e nitrogênio não proteico (NNP).
Avaliando os dados tabelados, observa-se que as menores concentrações de nitrogênio total presentes no meio se refletiram nas menores concentrações de proteína unicelular, ou seja, no início e meio da fase logarítmica no efluente e a partir do meio da fase log no meio BG11. Vários autores relatam que a composição, tanto em proteína como em carboidratos e lipídios, depende naturalmente de uma série de fatores que incluem o estágio da divisão celular, o ciclo de crescimento e a composição de micro e macronutrientes (LOURENÇO et al., 1997; FIDALGO et al., 1998; BENNING et al., 2004; SILVA; SILVA, 2007). Lourenço et al. (1997) encontraram concentrações de proteínas significativamente maiores (p < 0,01) na microalga Tetraselmis gracilis, quando utilizado o meio em que predominava como fonte de nitrogênio amônio ao invés de nitrato. Essas concentrações foram verificadas no final da fase logarítmica e na fase estacionária, da mesma forma que foi constatado no presente trabalho. Nestas fases, as concentrações de proteína foram 66,6 e 72,4% no efluente e 60,3 e 52,2% para o cultivo no meio BG11. Outro ponto em comum dos dois estudos é: quando a fonte de nitrogênio proveio do íon nitrato, as maiores concentrações de proteína ocorreram no início da fase exponencial. Isso poderia ser atribuído à adaptação prévia dos microrganismos em um meio contendo nitrato, somado ao fato de que os experimentos iniciam com as células na fase exponencial de crescimento. Isto pode explicar o porquê da maior concentração de proteína na biomassa cultivada no efluente do pescado no início da fase exponencial e o subsequente decréscimo no meio da fase log, logo após a adaptação ao meio, o que corresponde a aproximadamente 12 horas de cultivo (Figura 1).
Por outro lado, von Giani e Rückert (2004) estimaram a composição proteica em tratamentos contendo diferentes composições de nitrogênio (NH4++N-NO3- , N-NH4+ e NO3-) pela cianobactéria Microcystis viridis Lemmerman a 20 °C e ciclo de 12 horas claro/escuro. Para o experimento, que utilizou somente nitrato, a concentração final de proteína foi sempre maior do que nos experimentos que utilizaram apenas amônio e amônio mais nitrato. Na biomassa, em ambos os regimes, a quantidade de proteína representou mais do que 40% da amostra seca. Esse resultado está em concordância com estudos prévios de cianobactérias, como na Cyanothece sp. (SCHNEEGURT et al., 1995), Spirulina platensis (TOKUSOGLU; ÜNAL, 2003) e Aphanothece microscopica Nägeli (ZEPKA et al., 2008). Isso mostra que diferentes variáveis poderão atuar no aumento de produção celular e na composição química do microrganismo e, mesmo que existam condições aparentemente mais favoráveis, este pode obter a energia para a sua manutenção e crescimento de outras formas do que seria esperado, utilizando caminhos metabólicos alternativos.
No que se refere ao meio BG11, foi verificado que, mesmo utilizando uma concentração de NaNO3 menor (0,15 g.L-1), a concentração de proteína no final da fase logarítmica foi maior que a encontrada por Schneegurt et al. (1995), utilizando a cianobactéria Cyanothece sp. em cultivo com ciclo claro e escuro e meio contendo 1,5 g.L-1 de NaNO3, que encontrou 51,7% de proteína.
Zepka et al. (2008) estudaram o efeito de diferentes condições de secagem na biomassa da cianobactéria Aphanothece microscopica Nägeli cultivada no efluente da parboilização do arroz. Os autores verificaram que o teor proteico apresentou mínimo e máximo de 41,33 e 49,29%, respectivamente, e que o valor máximo foi obtido utilizando a temperatura de 60 °C. Verifica-se que, mesmo na melhor condição operacional, o teor proteico na biomassa da cianobactéria cultivada no efluente do arroz é inferior quando o mesmo microrganismo cresce no efluente do pescado, considerando as mesmas condições experimentais. Este fato demonstra o efeito da composição do meio de cultivo no perfil bioquímico, uma vez que o efluente oriundo do processamento de pescado se caracteriza por conter uma alta concentração em compostos nitrogenados orgânicos.
Quando são consideradas as frações dos ácidos nucleicos e amônio, verifica-se importante contribuição destes em todas as fases de crescimento, tanto quando a cianobactéria foi cultivada no efluente quanto no meio padrão BG11. Esses altos valores de compostos nitrogenados não proteicos refletiram na fração proteica calculada em função do N-NTK e (N-NTK)-NNP. As porcentagens de proteínas foram significativamente menores em todos os casos, atingindo mais de 50% de redução da quantidade de proteína, quando é considerada a fração não proteica. Os resultados demonstram que o fator tradicional de conversão de N-NTK em proteína (6,25) é inadequado, pois estima exagerada e erroneamente as concentrações de proteína presentes na biomassa de cianobactérias.
4 Conclusões
Os resultados obtidos para as frações nitrogenadas presentes na biomassa da cianobactéria Aphanothece microscopica Nägeli cultivada no efluente do processamento de pescado e no meio padrão BG11 mostraram que o nitrogênio total presente na biomassa reduz da fase logarítmica para a fase estacionária, independente do meio de cultura. O nitrogênio amoniacal juntamente com os ácidos nucleicos representa importante percentual em relação às frações nitrogenadas intracelulares. As concentrações do íon amônio foram maiores para a biomassa cultivada no efluente e o íon nitrato no meio BG11. O nitrogênio não proteico totaliza de 24,1 a 44,4% e de 35,8 a 65,6% do N-NTK presente na biomassa cultivada no efluente de pescado e meio BG11, respectivamente. Portanto, a utilização do fator de conversão de N-NTK em proteína (6,25) é inadequada, pois superestima a concentração real de proteína presente na biomassa da cianobactéria.
Recebido para publicação em 7/4/2008
Aceito para publicação em 5/1/2009 (003384)
- AMERICAN PUBLIC HEALTH ASSOCIATION - APHA. Standard Methods for the Examination of Water and Wastewater. 20 ed. Washington, D. C., 2000.
- ASPÉ, E.; MARTÍ, C.; ROECKEL, M. Anaerobic treatment of fishery wastewater using a marine sediment inoculum. Water Research, v. 31, n. 9, p. 2147-2160, 1997.
- ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS - AOAC. Official Methods of Analysis 15 ed. Washington, D.C., 1990.
- BENNING, L. G. et al. Molecular characterization of cyanobacterial silicification using synchrotron infrared micro-spectroscopy. Geochimica et Cosmochimica Acta, v. 68, n. 4, p. 729-741, 2004.
- BURJA, A. et al. Marine cyanobacteria: a profile source of natural products. Tetrahedron, v. 57, n. 46, p. 9347-9377, 2001.
- DE LA NOÜE, J.; LALIBERTÉ, G.; PROULX, D. Algae and wastewater. Journal of Applied Phycology, v. 4, n. 3, p. 247-254, 1992.
- DE PHILIPPIS, R.; VINCENZINI, M. Exocellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiology Reviews, v. 22, n. 3, p. 151-175, 1998.
- FAINTUCH, L.; SATOS, B.; AQUARONI, E. Emprego de algumas fontes de nitrogenados na obtenção de biomassa da Oscilatória limnética Revista de Microbiologia, v. 23, p. 32-33, 1992.
- FIDALGO, J. P. et al. Effects of nitrogen source and growth phase on proximate biochemical compostion, lipid classes and fatty acid profile of the marine microalga Isochrysis galbana Aquacult, v. 166, n. 1-2, p. 105-116, 1998.
- FUENTES, M. M. R. et al. Biomass nutrient profiles of the microalga Porphyridium cruentum Food Chemistry, v. 70, n. 3, p. 345-353, 2000.
- GARCÍA-FERNANDÉZ, J. M.; DIEZ, J. Adaptive mechanisms of nitrogen and carbon assimilatory pathways in the marine cyanobacteria Prochlorococcus Research in Microbiology, v. 155, n. 10, p. 795-802, 2004.
- GEIDER, R. J.; LaROCHE, J. Redfield revisited: variability of C:N:P in marine microalgae and its biochemical basis. European Journal of Phycology, v. 37, n. 1, p. 1-17, 2002.
- HAVENS, K. E. et al. N:P ratios, light limitation, and cyanobacterial dominance in a subtropical lake impacted by non-point source nutrient pollution. Environmental Pollution, v. 122, n. 3, p. 379-390, 2003.
- HERRERO, A.; FLORES, E.; GUERRERO, M. G. Regulation of nitrate reductase levels in the cyanobacteria Anacystis nidulans, Anabaena sp. Strain 7119, and Nostoc sp. Strain 6719. The Journal of Bacteriology, v. 145, n. 1, p. 175-180, 1981.
- HERRERO, A.; MURO-PASTOR, A. M.; FLORES, E. Nitrogen control in cyanobacteria. The Journal of Bacteriology, v. 183, n. 2, p. 411-425, 2001.
- HORNES, M. O.; QUEIROZ, M. I. Evaluation of the growth of cyanobacterium Aphanothece microscopica Nägeli in effluent of fishing industry. In: INTERNATIONAL CONGRESS OF CHEMICAL AND PROCESS ENGINEERING, 16, 2004, Prague.
- JACOB-LOPES, E. et al. Characteristics of thin-layer drying of the cyanobacterium Aphanothece microscopica Nägeli. Chemical Engineering and Processing, v. 46, p. 63-69, 2007.
- LEE, B. K.; KIM, J. K. Production of Cândida utilis biomass on molasses in different culture types. Aquacultural Engineering, v. 25, n. 2, p. 111-124, 2001.
- LIM, J.; KIM, T.; HWANG, S. Treatment of fish-processing wastewater by co-culture of candida rugopelliculosa and Brachionus plicatilis Water Research, v. 37, n. 9, p. 2228-2232, 2003.
- LOURENÇO, S. O. et al. Changes in Biochemical profile of Tetraselmis gracilis I. Compostion of two culture media. Aquacult, v. 148, n. 2-3, p. 153-168, 1997.
- LUQUE, I. et al. In vivo activity of the nitrogen control transcription factor NtcA is subjected to metabolic regulation in Synechococcus sp. strain PCC 7942. FEMS Microbiology Letters, v. 236, n. 1, p. 47-52, 2004.
- LUQUE, I.; FLORES, E.; HERRERO, A. Molecular mechanism for the operation of nitrogen control in cyanobacteria. The EMBO Journal, v. 13, n. 12, p. 2862-2869, 1994.
- MOLINA GRIMA, E. et al. Recovery of microalgal biomass and metabolites: process option and economics. Biotechnology Advances, v. 20, p. 491-515, 2002.
- MORIST, A. et al. Recovery and treatment of Spirulina platensis cells cultured in a continuous photobioreactor to be used as food. Process Biochemistry, v. 37, n. 5, p. 535-547, 2001.
- MURO-PASTOR, M. I.; FLORENCIO, F. J. Regulation of ammonium assimilation in cyanobacteria. Plant Physiology and Biochemistry, v. 41, n. 6-7, p. 595-603, 2003.
- ORTEGA, J. J. et al. Chemical composition of Spirulina and eukaryotic algae food products marketed in Spain. Journal of Applied Phycology, v. 5, n. 4, p. 425-435, 1993.
- PATEL, A. et al. Purification and characterization of C-Phycocyanin from cyanobacterial species of marine and freshwater habitat. Protein Expression and Purification, v. 40, n. 2, p. 248-255, 2005.
- QUEIROZ, M. I. et al. Evaluación del crescimiento de la Aphanothece microscopica Nägeli en las aguas residuales de la parbolización del arroz. Revista Información Tecnologica, v. 13, p. 61-66, 2002.
- QUEIROZ, M. I. et al. Evaluation of single cell protein production by Aphanothece microscopica Nägeli in batch reactor. Food Science and Biotechnology, v. 13, n. 3, p. 130-141, 2004.
- QUEIROZ, M. I. et al. The kinetics of the removal of nitrogen and organic matter from parboiled rice effluent by cyanobacteria in a stirred batch reactor. Bioresource Technology, v. 98, n. 11, p. 2163-2169, 2007.
- RAJOKA, M. I. et al. Kinetics of batch single cell protein production from rice polishings with Candida utilis in continuously aerated tank reactors. Bioresource Technology, v. 97, n. 15, p. 1934-1941, 2006.
- REYNOLDS, C. S. The ecology of freshwater phytoplankton: Cambridge studies in ecology. Cambridge: Cambridge University Press, 1984.
- RIPPKA, R. et al. Generic assignments strain histories and properties of pure cultures of cyanobacteria. Journal of General Microbiology, v. 111, p. 1-61, 1979.
- SAHA, S. K.; UMA, L.; SUBRAMANIAN, G. Nitrogen stress induced changes in the marine cyanobacterium Oscillatoria willei BDU 130511. FEMS Microbiology Ecology, v. 45, n. 3, p. 263-272, 2003.
- SÁNCHEZ-LUNA, L. D. et al. Continuous and pulse feedings of urea as a nitrogen source in fed-batch cultivation of Spirulina platensis Aquacultural Engineering, v. 31, n. 3-4, p. 237-245, 2004.
- SCHNEEGURT, M. A. et al. Compositional and toxicological evaluation of the diazotrophic cyanobacterium, Cyanothece sp. Strain ATCC 51142. Aquacult, v. 134, n. 3-4, p. 339-349, 1995.
- SGARBIERI, V. C. Proteínas em alimentos protéicos: Propriedades - Degradações - Modificações. São Paulo: Editora Unicamp, 1996.
- SHI, XIAN-MING; ZHANG, XUE-WU; CHEN, F. Heterotrophic production of biomass and lutein by Chlorella protothecoides on various nitrogen sources. Enzyme and Microbial Technology, v. 27, n. 3-5, p. 312-318, 2000.
- SILVA, P. G.; SILVA, H. J. Effect of mineral nutrients on cell growth and self-flocculation of Tolypothrix tenuis for the production of a biofertilizer. Bioresource Technology, v. 98, n. 3, p. 607-611, 2007.
- SPOLAORE, P. et al. Commercial applications of microalgae. Journal of Bioscience and Bioengineering , v. 101, n. 2, p. 87-96, 2006.
- STAL, L. J.; MOEZELAAR, R. Fermentation in cyanobacteria. FEMS Microbiology Reviews, v. 21, n. 2, p. 179-211, 1997.
- SYRETT, P. J.; CRESSWELL, R. C. Ammonium inhibition of nitrate uptake by the diatoms Phaeodactylum tricormutum Plant science letters, v. 14, n. 4, p. 321-325, 1979.
- TOKUSOGLU, Ö.; ÜNAL, M. K. Biomass nutrient profiles of three microalgae: Spirulina platensis, Chlorella vulgaris, and Isochrisis galbana Journal of Food Science, v. 68, n. 4, p. 1144-1148, 2003.
- von RÜCKERT, G.; GIANI, A. Effect of nitrate and ammonium on the growth and protein concentration of Microcystis viridis Lemmermann (Cyanobacteria). Revista Brasileira de Botânica, v. 27, n. 2, p. 325-331, 2004.
- VONSHAK, A.; ZENG, MING-TAO. Adaptation of Spirulina platensis to salinity-stress. Comparative Biochemistry and Physiology, v. 120, n. 1, p. 113-118, 1998.
- WITJITRA, K.; SHAH, M. M.; CHERYAN, M. Effect of nutrient sources on growth and acetate production by Clostridium thermoaceticum. Enzyme and Microbial Technology, v. 19, n. 5, p. 322-327, 1996.
- ZEPKA, L. Q. et al. Production and biochemical profile of the microalgae Aphanothece microscopica Nägeli submitted to different drying conditions. Chemical Engineering and Processing, v. 47, n. 8, p. 1311-1316, 2008.
Datas de Publicação
-
Publicação nesta coleção
05 Ago 2010 -
Data do Fascículo
Jun 2010
Histórico
-
Recebido
04 Jul 2008 -
Aceito
05 Jan 2009