Resumos
O presente trabalho teve o objetivo de avaliar as alterações fisiológicas e bioquímicas em sementes de feijão submetidas ao envelhecimento artificial. Utilizaram-se dois lotes de sementes de feijão, cultivar Iapar 44, com diferentes qualidades fisiológicas, submetidos ao envelhecimento artificial (41ºC e 100% UR do ar), por períodos de zero, 24, 48, 72 e 96 horas. As análises fisiológicas constaram de teste de germinação, primeira contagem de germinação, condutividade elétrica, comprimento de hipocótilo e de raízes das plântulas; as análises bioquímicas foram realizadas pela técnica de eletroforese de sistemas enzimáticos, avaliando-se a fosfatase ácida, malato e glutamato desidrogenase e esterase. Os resultados obtidos permitem concluir que: a) lotes com alta germinação apresentam acentuada diferença na resposta ao processo de envelhecimento, verificados pela redução na capacidade seletiva das membranas das sementes e pelo crescimento e desenvolvimento das plântulas de feijoeiro; b) a partir de 72 horas de envelhecimento artificial, há redução na atividade das enzimas fosfatase ácida e esterase; c) a atividade das enzimas fosfatase ácida, malato desidrogenase, glutamato desidrogenase e esterase é influenciada pelo período de envelhecimento e pela qualidade inicial dos lotes
Phaseolus vulgaris; deterioração; fosfatase ácida; malato desidrogenase; glutamato desidrogenase; esterase
The present work had the objective of evaluating the physiologic and biochemical alterations in bean seeds submitted to the artificial ageing. Two lots of bean seeds were used, cultivar Iapar 44, with different physiologic qualities, submitted to the artificial ageing (41ºC - 100% UR of the air), for zero, 24, 48, 72 and 96 hours. The physiologic analyses consisted of germination test, first germination count, electric conductivity, seedling and root length; the biochemical analyses were accomplished by the electrophoretical technique of enzymatic systems, being evaluated the acid phosphatase, malate and glutamate dehydrogenase and esterase. The obtained results concluded, a) lots with high germination presents accentuated difference in the answer to the ageing process, verified by the reduction in the selective capacity of the membranes of the seeds and growth and development of the bean seedlings; b) starting from 72 hours of artificial aging, there is reduction in the activity of the enzymes acid phosphatase and esterase; c) the activity of the enzymes acid phosphatase, malate and glutamate dehydrogenase and esterase are influenced by the ageing period and for the quality of the lots of seeds.
Phaseolus vulgaris; deterioration; acid phosphatase; malate dehydrogenase; glutamate dehydrogenase; esterase
Alterações fisiológicas e bioquímicas em sementes de feijão envelhecidas artificialmente1 1 Submetido em 31/07/2003. Aprovado em 06/01/2004
Physiologic and biochemical alterations in artificially aged bean seeds
Candice Mello Romero SantosI; Nilson Lemos de MenezesII; Francisco Amaral VillelaIII
IEngº. Agrª. Drª. SQN 204, bloco G, apt 301, Asa Norte- Brasília-DF. 70842-070. E-mail: candice_romero@hotmail.com
IIEngº. Agrº. Dr., Professor Adjunto do Depto. Fitotecnia da Universidade Federal de Santa Maria. 97105-900.E-mail: nlmenezes@smail.ufsm.br
IIIEng. Agrícola, Dr., Professor Adjunto do Depto. de Fitotecnia da Universidade Federal de Pelotas. Caixa Postal 354 96010-900. Email: bloisvillela@uol.com.br
RESUMO
O presente trabalho teve o objetivo de avaliar as alterações fisiológicas e bioquímicas em sementes de feijão submetidas ao envelhecimento artificial. Utilizaram-se dois lotes de sementes de feijão, cultivar Iapar 44, com diferentes qualidades fisiológicas, submetidos ao envelhecimento artificial (41ºC e 100% UR do ar), por períodos de zero, 24, 48, 72 e 96 horas. As análises fisiológicas constaram de teste de germinação, primeira contagem de germinação, condutividade elétrica, comprimento de hipocótilo e de raízes das plântulas; as análises bioquímicas foram realizadas pela técnica de eletroforese de sistemas enzimáticos, avaliando-se a fosfatase ácida, malato e glutamato desidrogenase e esterase. Os resultados obtidos permitem concluir que: a) lotes com alta germinação apresentam acentuada diferença na resposta ao processo de envelhecimento, verificados pela redução na capacidade seletiva das membranas das sementes e pelo crescimento e desenvolvimento das plântulas de feijoeiro; b) a partir de 72 horas de envelhecimento artificial, há redução na atividade das enzimas fosfatase ácida e esterase; c) a atividade das enzimas fosfatase ácida, malato desidrogenase, glutamato desidrogenase e esterase é influenciada pelo período de envelhecimento e pela qualidade inicial dos lotes
Termos para indexação:Phaseolus vulgaris, deterioração, fosfatase ácida, malato desidrogenase, glutamato desidrogenase, esterase.
ABSTRACT
The present work had the objective of evaluating the physiologic and biochemical alterations in bean seeds submitted to the artificial ageing. Two lots of bean seeds were used, cultivar Iapar 44, with different physiologic qualities, submitted to the artificial ageing (41ºC - 100% UR of the air), for zero, 24, 48, 72 and 96 hours. The physiologic analyses consisted of germination test, first germination count, electric conductivity, seedling and root length; the biochemical analyses were accomplished by the electrophoretical technique of enzymatic systems, being evaluated the acid phosphatase, malate and glutamate dehydrogenase and esterase. The obtained results concluded, a) lots with high germination presents accentuated difference in the answer to the ageing process, verified by the reduction in the selective capacity of the membranes of the seeds and growth and development of the bean seedlings; b) starting from 72 hours of artificial aging, there is reduction in the activity of the enzymes acid phosphatase and esterase; c) the activity of the enzymes acid phosphatase, malate and glutamate dehydrogenase and esterase are influenced by the ageing period and for the quality of the lots of seeds.
Index terms:Phaseolus vulgaris, deterioration, acid phosphatase, malate dehydrogenase, glutamate dehydrogenase, esterase.
INTRODUÇÃO
A qualidade fisiológica das sementes é máxima por ocasião da maturidade fisiológica. A partir deste momento processos degenerativos, de natureza física, fisiológica ou bioquímica começam a ocorrer, caracterizando a deterioração.
A deterioração é considerada toda e qualquer mudança degenerativa, após a semente ter atingido sua máxima qualidade, os mecanismos responsáveis por essas modificações ainda não foram totalmente elucidados. A sensibilidade das sementes ao processo de deterioração, em determinado ambiente, tem sido atribuída à constituição genética. Além disso, as condições de temperatura e umidade relativa do ar, durante o armazenamento, são de grande importância na evolução da deterioração, a qual não pode ser evitada, mas pode ser minimizada no armazenamento sob condições adequadas.
A deterioração é evidenciada por danos genéticos, perda da integridade do sistema de membranas, redução da capacidade seletiva, peroxidação de lipídios, lixiviação de solutos, mudanças na atividade respiratória das sementes, modificações na atividade enzimática e síntese de proteínas, a incapacidade de manutenção do gradiente eletroquímico e a perda da compartimentalização celular e acúmulo de substâncias tóxicas. Também foram observadas alterações fisiológicas, tais como: atraso na germinação, decréscimo na tolerância às condições ambientais sub-ótimas durante a germinação, redução no crescimento e/ou vigor das plântulas, aumento do número de plântulas anormais, maior suscetibilidade a ataques de microrganismos patogênicos, emergência desuniforme, redução na produtividade, modificações na coloração das sementes, diminuição do potencial de armazenamento, completa perda da capacidade germinativa e a morte das sementes (Wilson & McDonald, 1986; Basavarajappa et al., 1991).
Reduções na atividade enzimática durante a germinação de sementes envelhecidas pode ser resultante da diminuição da síntese protéica sendo que o início da deterioração das sementes geralmente está associado à redução na síntese de proteínas (Abdul-Baki & Anderson, 1972). O envelhecimento de sementes de trigo, segundo Dell'aquila (1994), foi acompanhado pelo atraso progressivo do início da germinação e pelo decréscimo de incorporação de metionina em proteínas solúveis de embriões hidratados.
O principal desafio das pesquisas sobre testes de vigor está na identificação dos eventos relacionados à deterioração das sementes, que precedam a perda da capacidade germinativa.
A técnica de eletroforese de proteínas é capaz de auxiliar a detecção dos estádios iniciais de deterioração através da avaliação da atividade de enzimas associadas à degradação e oxidação de substâncias de reserva, bem como com a biossíntese de novas substâncias (Spinola et al., 2000). A capacidade de avaliação e a correta interpretação da variação eletroforética nos perfis de proteínas e enzimas pode ser uma eficiente ferramenta na determinação de mudanças bioquímicas resultantes do processo deteriorativo (Carraro,1990). Possíveis indicadores do estádio de deterioração em sementes de algodoeiro foram encontrados por Vieira (1996), através das variações eletroforéticas de proteínas e das enzimas glutamato desidrogenase, malato desidrogenase, fosfatase ácida, enzima málica, peroxidase e 6 - fosfogluconato.
Em várias espécies, mudanças na atividade de enzimas em sementes secas e germinadas ocorrem concomitantemente com um decréscimo na qualidade da semente no armazenamento, e aumento na atividade de proteinases, b -amilases e nucleases foram relatadas por Basavarajappa et al.(1991); Bewley & Black (1994); Salinas et al. (1998) e Padilha et al. (2001).
A susceptibilidade das sementes ao estresse durante o envelhecimento artificial pode ser correlacionada com a qualidade da semente. Foram observadas alterações nos lipídios e nas proteínas insolúveis em ácido em decorrência do envelhecimento artificial (Ferguson et al., 1990). Diferentes mudanças na atividade enzimática, em conseqüência do envelhecimento artificial, foram observados por Bock (1999), em sementes de soja, indicando que o envelhecimento promove desorganização nos sistemas enzimáticos da semente.
Em sementes de cevada e soja com teor de água de 11 a 13%, à medida que o processo de envelhecimento progrediu, houve aumento no número de bandas de proteínas e da enzima esterase, indicando elevação da atividade metabólica (Chauhan, 1985).
Contudo, o método mais utilizado para avaliar o nível de deterioração da semente é o envelhecimento artificial, cujo princípio está fundamentado no aumento da deterioração das sementes pela exposição a níveis elevados de temperatura e umidade relativa do ar.
Embora muitos trabalhos tenham avaliado a influência do envelhecimento artificial em sementes, poucos evidenciam as conseqüências em termos de alterações enzimáticas. Portanto, para avaliar a ocorrência de alterações no vigor, germinação, e na atividade das enzimas malato e glutamato desidrogenase, esterase e fosfatase ácida em sementes de feijão submetidas ao envelhecimento artificial, foi desenvolvido o presente trabalho.
MATERIAL E MÉTODOS
O presente trabalho foi conduzido no Laboratório de Bio-sementes, da Faculdade de Agronomia Eliseu Maciel da Universidade Federal de Pelotas RS e no Laboratório de Análise de Sementes de Produção (LASP), do Departamento de Fitotecnia na Universidade Federal de Santa Maria - RS. Utilizaram-se dois lotes de sementes de feijão (Phaseolus vulgaris L.), cultivar IAPAR 44, que apresentaram qualidades fisiológicas distintas (Tabela 1).
A qualidade dos lotes foi avaliada pelos testes de germinação, primeira contagem da germinação, comprimento de raízes e de hipocótilo das plântulas e condutividade elétrica.
Envelhecimento artificial - utilizaram-se 200 sementes, distribuídas em subamostras de 50 sementes, para cada lote de sementes de feijão. Os teores iniciais de água foram ajustados até atingirem ± 13%, conforme recomenda Marcos Filho (1994). A seguir, as sementes foram colocadas sobre uma bandeja de tela de alumínio, fixada no interior de caixas plásticas, as quais funcionaram como compartimento individual (mini-câmara). Foram colocados 40mL de água destilada no interior destas caixas plásticas, que foram mantidas em incubadora a temperatura de 41º C. Os tratamentos constaram do teste de envelhecimento acelerado, por períodos de zero, 24, 48, 72 e 96 horas.
Teor de água das sementes - realizou-se a determinação do teor de água das sementes, antes e após os tratamentos de envelhecimento artificial, pelo método da estufa a temperatura de 105 ± 3º C, durante 24 horas, com duas amostras por lote de sementes (Brasil, 1992). Os resultados foram expressos em porcentagem média por lote. O teor de água das sementes expostas aos tratamentos, de envelhecimento acelerado, não foram ajustados para a realização dos testes.
Teste de germinação - utilizaram-se quatro subamostras de 50 sementes, para cada tratamento de período de envelhecimento artificial, em rolo de papel umedecido com água destilada na quantidade de 2,5 vezes o peso do papel seco, os quais foram mantidos em germinador regulado à temperatura constante de 25º C, por um período de sete dias. A interpretação foi realizada no sétimo dia após instalação, segundo critérios estabelecidos pelas Regras para Análise de Sementes (Brasil, 1992). Computaram-se as plântulas normais para cada repetição e obtendo-se a seguir uma média das subamostras. Os resultados foram expressos em porcentagem.
Primeira contagem do teste de germinação - foi determinada conjuntamente com o teste de germinação. Computaram-se as plântulas normais na primeira contagem do teste de germinação, realizada no quarto dia após a instalação do teste. Os resultados foram obtidos pela média aritmética das quarto subamostras e foram expressos em porcentagem.
Comprimento de hipocótilo e de raízes - utilizaram-se quatro subamostras de 25 sementes semeadas em rolos de papel, sobre uma linha horizontal no terço superior do papel umedecido com água destilada na quantidade de 2,5 vezes o peso do papel seco. Os rolos contendo as sementes permaneceram em germinador por quatro dias a temperatura de 25º C e após procedeu-se à medição (Nakagawa, 1999). Os comprimentos médios do hipocótilo e das raízes das plântulas normais foram obtidos dividindo-se a soma das medidas tomadas das subamostras pelo número de plântulas normais mensuradas. Os resultados foram expressos em centímetros.
Condutividade elétrica - foi conduzido conforme Vieira (1994), pesaram-se quatro subamostras de 50 sementes, por lote para cada tratamento. Após a pesagem de cada amostra, as sementes foram colocadas em copos plásticos contendo 75mL de água destilada e mantidas em germinador a temperatura de 25ºC, durante 24 horas. Decorrido esse período, a condutividade elétrica da solução foi determinada com o uso de um condutivimetro de marca DIGIMED CD-21 e os valores obtidos no aparelho foram divididos pelo peso da amostra (g) e os resultados foram expressos em m S.cm-1.g-1 de sementes.
Avaliação das alterações bioquímicas - determinou-se a atividade enzimática das sementes de feijão dos dois lotes, antes e após serem submetidos aos tratamentos de envelhecimento artificial. Foram retiradas subamostras de sementes de cada lote e mantidas em temperatura de ± 5ºC, para posterior determinação da atividade das enzimas malato desidrogenase, glutamato desidrogenase, esterase e fosfatase ácida. Utilizou-se a técnica de eletroforese em gel vertical de poliacrilamida na concentração de 6%, para verificar a atividade das enzimas. Para a obtenção do eletroforegrama foi utilizado o sistema tampão contínuo, conforme descrito por Scandálios (1969). O preparo das amostras foi realizado utilizando-se uma semente triturada por repetição. As amostras foram maceradas em recipiente de porcelana sobre gelo para evitar a desnaturação de proteínas pelo aquecimento; a seguir, procedeu-se a pesagem de 50-100mg da amostra e a colocação da solução extratora, conforme descrito Scandálios (1969), na proporção de 5:1 (volume/peso). As amostras permaneceram no extrator durante 18 horas à baixa temperatura (± 5ºC). Posteriormente, as amostras foram centrifugadas por cinco minutos e foram adicionados 25m l do sobrenadante do material na parte superior do gel. Realizou-se a corrida eletroforética dentro de balcão refrigerado 4º C, com polaridade normal e constante de 25mA. Foram usados os sistemas de revelação citados por Alfenas (1998), para as enzimas malato e glutamato desidrogenase e o sistema recomendado por Scandálios (1969) para as enzimas esterase e fosfatase ácida. Em cada gel, foram obtidos todos os tratamentos, sendo quatro repetições para cada tratamento. Os géis foram fotografados e digitalizados utilizando o "Kodak Electrophorese Documentation and Analysis System" (EDAS 120) e a identificação das bandas e determinação das densidades relativas pelo programa "Gel-Pro Analyser" (Media Cybernectis, 1997).
Análise estatística - Utilizou-se o delineamento inteiramente casualizado em esquema bifatorial 2x5, dois lotes e cinco períodos de envelhecimento artificial, com quatro repetições. Foram realizadas análises da variância e o estudo da regressão polinomial. Os dados referentes às porcentagens de germinação, primeira contagem de germinação e densidade relativa foram transformados em arcsen (%/100)1/2.
RESULTADOS E DISCUSSÃO
Os lotes utilizados neste trabalho possuíam qualidade fisiológica distintas (Tabela 1), os lotes 1 e 2 apresentaram 90 e 94% de germinação, respectivamente. Quando se avaliou o vigor dos lotes através da velocidade de germinação e permeabilidade de membranas, o lote 1 apresentou-se menos vigoroso.
Os teores de água das sementes antes de cada período de exposição ao envelhecimento artificial foram inicialmente de 13,5 e 13,2%, para os lotes 1 e 2, respectivamente. Marcos Filho (1994) ressaltou a importância da uniformização do teor de água das sementes antes do envelhecimento artificial, uma vez que, as sementes mais úmidas sofrerão maiores efeitos deletérios do estresse, devido à intensificação da sua atividade metabólica.
Os teores de água das sementes dos lotes 1 e 2 cresceram com o aumento no período de exposição das sementes ao envelhecimento artificial, embora o lote 1 tenha apresentado maior incremento (Tabela 2). Essa ocorrência poderia ser atribuída ao fato do lote 1, provavelmente, apresentar menor integridade do seu sistema de membranas e/ou menor seletividade, permitindo a entrada de água mais rapidamente nas células. Estes resultados concordam com os dados encontrados por Souza et al. (1996), ao verificarem em sementes de calopogônio, que no final do período de embebição, os lotes que exibiram maiores proporções de sementes intumescidas eram os de menor qualidade fisiológica.
O estresse provocado às sementes pela umidade relativa do ar e temperatura elevada, afetou o teor de água das sementes, intensificando a atividade metabólica e, de acordo com Delouche (2002), a duração do processo de deterioração é determinada principalmente pela interação entre herança genética, o grau de hidratação da semente e a temperatura.
Pode-se observar, na Figura 1A, que os tratamentos praticamente não afetaram a germinação do lote 2, entretanto o lote 1, diminuiu sua porcentagem de germinação com o aumento do período de envelhecimento, mostrando-se como o lote menos vigoroso. Após 96 horas de envelhecimento, o lote 1 apresentou menos de 15% de plântulas normais no teste de germinação, o que mostra o quanto sua qualidade era baixa uma vez que, segundo Delouche (2002) a perda da capacidade de germinação é a conseqüência ou efeito final da deterioração, sendo o último atributo da qualidade fisiológica da semente que é perdido.

Na primeira contagem do teste de germinação (Figura 1B), ambos os lotes apresentaram redução declínio da porcentagem de plântulas normais com os diferentes períodos de envelhecimento artificial, entretanto o lote 1 foi mais afetado, sofrendo maior atraso para o início da germinação. O envelhecimento artificial está baseado no fato da taxa de deterioração das sementes acelerar conforme a exposição aos níveis elevados de temperatura e umidade relativa do ar, justificando desta forma o maior decréscimo da porcentagem de plântulas normais apresentadas no lote 1, comparativamente ao lote 2, de maior qualidade fisiológica.
Através do teste de condutividade elétrica (Figura 1C), que avalia a permeabilidade seletiva das membranas celulares, foi possível evidenciar que o aumento nos períodos de envelhecimento aumentou a lixiviação de exsudatos em ambos os lotes de feijoeiro. No lote 1, parece ter ocorrido maiores mudanças deteriorativas, desintegração do sistema de membranas, descontrole de trocas hídricas e de solutos entre as células e o exterior, menor capacidade de reparação aos danos causados à dupla camada fosfolipídica e proteínas na membrana plasmática, tornando-a menos seletiva. Segundo Copeland & McDonald (1995), o grau de deterioração das sementes está associado com a concentração de exsudatos das sementes na solução, e estes são o reflexo da degradação das membranas. É válido ressaltar que, conforme Powell & Matthews (1977) e Delouche (2002), os danos nas membranas são os eventos iniciais das alterações degenerativas nas sementes. O lote 2, que exibiu menor lixiviação, possivelmente apresenta maior capacidade para reorganizar e reparar danos nas membranas, sendo, portanto, de qualidade superior.
Na avaliação do comprimento de hipocótilo e de raízes das plântulas de feijoeiro (Figuras 2A e 2B), notou-se que os tratamentos de envelhecimento artificial reduziram o comprimento do hipocótilo e de raízes das plântulas com o aumento do período de exposição, em ambos os lotes. Entretanto, o lote 1 apresentou maior sensibilidade aos tratamentos, exibindo acentuada diminuição nos comprimentos de hipocótilo e raízes quando comparado ao lote 2. Este fato pode ser justificado pelo menor vigor do lote 1. As sementes do lote 2 originaram plântulas com maior taxa de crescimento, provavelmente em função de apresentarem maior capacidade de suprimento de reservas e transformação dos tecidos de armazenamento e maior incorporação destes pelo eixo embrionário, de acordo com Dan et al. (1987).

Os resultados observados para os lote 1 e 2 concordam com aqueles observados por Matthews (1985) que relatou que, sob o aspecto do comportamento fisiológico da semente, a manifestação inicial do processo de envelhecimento é o declínio da velocidade de germinação seguido pela redução do tamanho das plântulas. A redução na velocidade de germinação pode ser determinada pela morte parcial ou total de tecidos importantes, em diferentes regiões da semente.
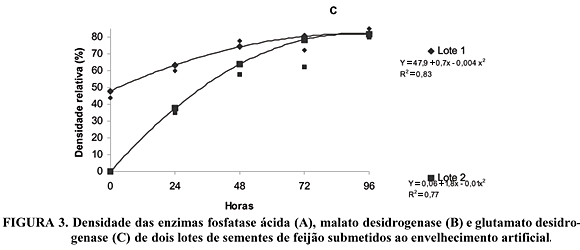
A atividade da enzima fosfatase ácida nos dois lotes de sementes de feijão (Figura 3A), foi afetada pelos tratamentos de envelhecimento. A fosfatase ácida é uma hidrolase que participa em reações de hidrólise de ésteres, podendo atuar sobre os fosfolipídios de membrana, provocando a peroxidação destes, principalmente da fosfatil colina, fosfatidil etanolamina e fosfatidil inositol. Esta enzima está envolvida também na manutenção do fosfato celular, podendo sua atividade afetar o metabolismo do fosfato em sementes, como os níveis de ATP e nucleotídeos. No lote 1, a atividade desta enzima foi maior no período de 24 a 48 horas, sendo máxima com 44,5 horas, ocorrendo, a partir daí redução na atividade. Para o lote 2, verificou-se que a atividade enzimática foi mais elevada no período de 48 a 72 horas, sendo máxima com 56 horas, e só a partir deste período houve redução da atividade. Notou-se também que a atividade desta enzima, no período zero, era maior em lotes com menor qualidade fisiológica, possivelmente, esta hidrolase já deveria estar atuando nos fosfolipídios de membrana, provocando peroxidação.

. Os valores da condutividade elétrica do lote 2 apresentaram aumento mais acentuado no período de 24 e 72 horas, relacionando-se com a maior atividade da fosfatase ácida durante este período, a qual pode ter provocado a peroxidação dos fosfolipídios de membrana e, conseqüentemente, aumento de sua permeabilidade. Estes dados concordam com Spinola et al. (2000), os quais observaram que esta enzima teve menor atividade em lotes de sementes de milho com qualidade fisiológica superior. Entretanto, Chauhan et al. (1985), Basavarajappa et al. (1991), Sung & Jeng (1994) e Bock (1999) relataram que em sementes de soja, amendoim, milho, repolho e soja, respectivamente, a atividade da fosfatase ácida decresceu com envelhecimento artificial. No presente trabalho, tal ocorrência só foi verificada após 44,5 e 56 horas, respectivamente, para os lotes 1 e 2.
A atividade da malato desidrogenase (Figura 3B) para o lote 1, foi máxima no período de 72,7 horas, com decréscimos a partir daí, enquanto que, no lote 2 a atividade desta enzima foi crescente conforme o aumento do período de estresse durante o envelhecimento artificial. A partir de 72,7 horas de envelhecimento ocorreu redução na capacidade respiratória das sementes do lote 1, de menor qualidade, possivelmente devido aos danos nas membranas mitocondriais, sendo esta organela a mais suscetível à peroxidação. Pois, conforme Wilson & McDonald (1986) e Ferguson et al. (1990), a membrana mitocondrial é rica em lipídios insaturados, o que aumenta a superfície exposta a peroxidação, elevando a permeabilidade das membranas e afetando a atividade respiratória aeróbica, por quebrar o gradiente protônico necessário para a manutenção do acoplamento respiratório, diminuindo a produção de ATP e absorção de oxigênio.
A enzima malato desidrogenase atua com a fosfoenolpiruvato carboxilase, para a redução de oxaloacetado a malato, utilizando NAD como doador ou receptor de elétrons nessas reações, tendo uma importante função no Ciclo de Krebs (mitocôndria) para a produção de NADH. E, as mitocôndrias constituem-se no centro da respiração, por isso fica evidente a importância dos efeitos da deterioração sobre o desempenho germinativo da semente quando são consideradas as modificações ocorridas nessa organela. Conforme Salinas et al. (1998), as mitocôndrias dos eixos embrionários são responsáveis pelo fornecimento de energia usada no alongamento dos eixos raiz/caule e, se a taxa respiratória diminui nos eixos, a emergência e crescimento das plântulas também diminuirão. Estudos realizados por Basavarajappa et al. (1991), com o teste de tetrazólio demonstraram gradual diminuição na atividade das desidrogenases em sementes envelhecidas de milho, indicando que esta perda poderia estar associada ao baixo nível de produção de ATP.
A atividade da glutamato desidrogenase no período zero apresentou-se mais elevada no lote 1 (Figura 3C), demonstrando que a atividade respiratória das sementes de lote estava mais elevada, caracterizando um processo deteriorativo mais avançado, quando comparado ao lote 2. A atividade desta enzima foi superior no lote 1 até 72 horas, no entanto, a partir desse período foi semelhante para os dois lotes. Esta enzima é responsável pela oxidação de aminoácidos, fornecendo energia para o Ciclo de Krebs, produção de NADPH e/ou na redução do a -cetoglutarato para síntese de aminoácidos. O estresse provocando pelo período de 96 horas de envelhecimento artificial não foi suficiente para diminuir a atividade desta enzima, em ambos os lotes, sendo esta a menos afetada pelos períodos de estresse, embora tendendo para a estabilização, sendo máxima após 87,5 horas, para o lote 1 e 90 horas para o lote 2. Em plântulas de eucalipto envelhecidas artificialmente, Camargo et al. (2000) observaram aumento na atividade da enzima glutamato desidrogenase, concordando com os dados encontrados neste trabalho e discordando dos dados encontrados por Shatters et al. (1994), os quais verificaram diminuição na atividade desta enzima em sementes de soja, a partir de 48 horas de envelhecimento.
No presente trabalho não foram testados períodos superiores a 96 horas de estresse, entretanto Brandão-Júnior et al. (1999) descreveram que a atividade desta enzima em sementes de milho diminui a partir de 96 horas de envelhecimento.
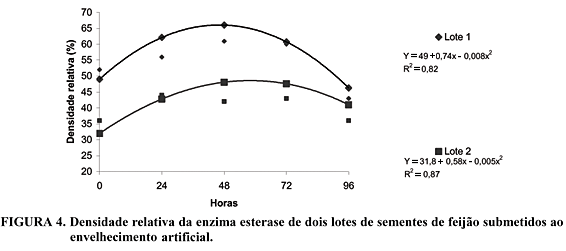
A atividade da enzima esterase no período zero foi mais elevada no lote 1, demonstrando que as sementes deste lote apresentavam maior peroxidação de seus lipídios de membrana, causando um aumento na permeabilidade das membranas e em processo deteriorativo maior do que o lote 2. A atividade desta enzima para os lotes 1 e 2 foi máxima, em 46 e 58 horas de envelhecimento artificial, respectivamente, a partir dos quais ocorreu diminuição na atividade desta enzima (Figura 4). O lote 1 apresentou maiores valores de atividade quando comparado ao lote 2. Estes resultados concordam com os encontrados por Padilha et al. (2001) que verificaram diminuição da intensidade das bandas para os estresses mais drásticos em sementes de milho e Chauhan (1985) e Bock (1999), que verificaram aumento no número de bandas desta enzima, em sementes envelhecidas de cevada e soja, respectivamente. Alterações nos padrões da enzima esterase evidenciam a ocorrência de eventos deteriorativos, que podem contribuir para a redução na germinação das sementes à medida que são aumentados os níveis de fatores adversos de temperatura e teor de água das sementes no processo de envelhecimento artificial, pois a esterase é uma enzima envolvida em reações de hidrólise de ésteres, estando diretamente ligada ao metabolismo dos lipídios.
CONCLUSÕES
Lotes com alta germinação podem apresentar acentuadas diferenças em resposta ao processo de envelhecimento, verificados pela redução na capacidade seletiva das membranas das sementes e pelo crescimento e desenvolvimento das plântulas de feijoeiro,
A partir de 72 horas de envelhecimento artificial, há redução na atividade das enzimas fosfatase ácida e esterase;
A atividade das enzimas fosfatase ácida, malato desidrogenase, glutamato desidrogenase e esterase é influenciada pelo período de envelhecimento e pela qualidade inicial dos lotes.
- ALFENAS, A.C. Eletroforese de isoenzimas e proteínas afins. Viçosa: UFV, 1998. 574 p.
- ABDUL-BAKI, A.A.; ANDERSON, J.D. Physiological and biochemical deterioration of seeds. In: KOZLOWSKI, T.T. Seed Biology New York: Academic Press, 1972. v.2, p. 283-315.
- BASAVARAJAPPA, B.S.; SHETTY, H.S.; PRAKASH, H.S. Membrane deterioration and other biochemical changes, associated with accelerated ageing of maize seeds. Seed Science and Technology, Zürich, v. 19, n. 2, p. 279-286, 1991.
- BEWLEY, J.D.; BLACK, M. Seeds: physiology of development and germination. New York: Plenum Press, 1994. 445 p.
- BOCK, F.L. Resposta a nível molecular do envelhecimento artificial, natural e pré-condicionamento de sementes de soja. Pelotas. 1999. 27f. Dissertação (Mestrado em Ciência e Tecnologia de Sementes), Universidade Federal de Pelotas, 1999.
- BRASIL. Ministério da Agricultura e Reforma Agrária. Regras para análise de sementes Brasília: SNAD/DNPV/CLAV,1992. 365 p.
- BRANDÃO-JUNIOR, D.S.; CARVALHO, M.L.M.; VIEIRA, M.G.G.C. Variações eletroforéticas de proteínas e isoenzimas relativas à deterioração de sementes de milho envelhecidas artificialmente. Revista Brasileira de Sementes, Brasília, v. 21, n.1, p. 114-121, 1999.
- CAMARGO, M.L.P.; MORI, E.S.; DE MELLO, E.J.; ODA, S.; LIMA, G.P. Atividade enzimática de sementes envelhecidas artificial e naturalmente. Ciência Florestal, Santa Maria, v. 10, n. 2, p.113-122, 2000.
- CARRARO, D.M. Variação e herança dos padrões eletroforéticos em órgãos e estágios de desenvolvimento em milho (Zea mays L.) Piracicaba, 1990. 121f. Dissertação (Mestrado em Agronomia) - Escola Superior de Agricultura Luiz de Queiroz, 1990.
- CHAUHAN, K.P.S. Electrophoretic variations of proteins and enzymes in relation to seed quality. Seed Science & Technology, Zurich , v.13, p. 629-641, 1985.
- DAN, E.L.; MELLO, V.D.C.; WETZEL, C.T.; POPINIGIS, F.; SOUZA, E.P. Transferência de matéria seca como método de avaliação do vigor de sementes de soja. Revista Brasileira de Sementes, Brasília, v. 9, n. 3, p. 45-55, 1987.
- DELL'AQUILA, A. Wheat seed ageing and embryo protein degradation. Seed Science Research, Wallingford, v. 4, n. 2, p.293-298, 1994.
- DELOUCHE, J. Germinação, deterioração e vigor da semente. Seed News, Pelotas, n. 6, p.24-31, 2002.
- FERGUSON, J.M.; TEKRONY, D.M.; EGLI, D.B. Changes during early soybean seed and axes deterioration: II. Lipids. Crop Science, Madison, v. 30, p. 179-182, 1990.
- MARCOS FILHO, J. Teste de envelhecimento acelerado. In: VIEIRA, R.D.; CARVALHO, N.M. (Ed.). Testes de vigor em sementes. Jaboticabal: FUNEP, 1994. p.133 -149.
- MEDIA CYBERNETICS. Gel - Pro analyser Version 3.0 for windows user's guide. Media Cybernetics, L.P. Silver Spring, 1997.
- NAKAGAWA, J. Testes de vigor baseados no desempenho das plântulas. In: KRZYZANOWSKI, F.C.; VIEIRA, R.D.; FRANÇA NETO, J.B. (Ed.). Vigor de sementes: conceitos e testes. Londrina: ABRATES, 1999. p. 1-23.
- PADILHA, L.; VIEIRA, M.G.G.C.; VON PINHO, E.V.R. Relação entre o teste de deterioração controlada e o desempenho de sementes de milho em diferentes condições de estresse. Revista Brasileira de Sementes, Brasília, v. 23, n.1, p. 198-204, 2001.
- POWELL, A.A.; MATTHEWS, S. Deteriorative changes in pea seeds (Pisum sativum L.) stored in humid or dry conditions. Journal of Experimental Botany, Oxford, v. 28, p.227-236, 1977.
- SALINAS, A.R.; SANTOS, O.S.B.; VILLELA, F.A.; SANTOS FILHO, B.G.; SOUZA SOARES, L.A.; OLIVEIRA, M.F. Fisiologia da deterioração em sementes de soja (Glycine max (L.) Merril) durante o armazenamento. Revista Científica Rural, Bagé, v.3, n.2 p. 106-118, 1998.
- SHATTERS, R.G.JR.; ABDELGHANY, A.; ELBAGOURY, O.; WEST, S.H. Soybean seed deterioration and response to priming: changes in specific enzyme activities in extracts from dry and germinating seeds. Seed Science and Technology, Zürich, v.4, p.33-41, 1994.
- SCANDÁLIOS, J.G. Genetic control of multiple molecular forms of enzymes in plants: a review. Biochemical Genetics, New York, v. 3, p.37-79, 1969.
- SPINOLA, M.C.M.; CICERO, S.M.; MELO, M. Alterações bioquímicas e fisiológicas em sementes de milho causadas pelo envelhecimento acelerado. Scientia Agrícola, Piracicaba, v. 57, n. 2, p. 263-270, 2000.
- SOUZA, F.H.D.; MARCOS FILHO, J..; NOGUEIRA, M.C. Características físicas das sementes de Calopogonium mucunoides Desv. associadas à qualidade fisiológica e ao padrão de absorção de água I. Tamanho. Revista Brasileira de Sementes, Brasília, v. 18, n.1, p. 33-40, 1996.
- SUNG, J.M.; JENG, T.L. Lipid peroxidation and peroxide scavenging enzymes associated with accelerated ageing of peanut seed. Physiologia Plantarum, Copenhagen, v. 91, n.1, p. 51-55, 1994.
- VIEIRA, R.D. Teste de condutividade elétrica. In: VIEIRA, R.D.; CARVALHO, N.M.(Ed.). Testes de vigor em sementes. Jaboticabal: FUNEP, 1994. p.103-132.
- VIEIRA, M.G.G.C. Utilização de marcadores moleculares no monitoramento da qualidade sanitária e nível de deterioração de sementes de algodoeiro (Gossypium hirsutum L.). Lavras. 1996. 114f. Tese (Doutorado em Fitotecnia). Universidade Federal de Lavras, 1996.
- WILSON, D.O.; MCDONALD, M.B. The lipid peroxidation model of seed ageing. Seed Science and Technology, Zürich, v. 14, p. 269-300,1986.
Datas de Publicação
-
Publicação nesta coleção
14 Fev 2005 -
Data do Fascículo
2004
Histórico
-
Recebido
31 Jul 2003 -
Aceito
01 Jun 2004