Resumos
O momento da colheita e os métodos de secagem podem influenciar a qualidade das sementes de cafeeiro durante o armazenamento. Este trabalho foi realizado com o objetivo de avaliar os efeitos do estádio de maturação e do método de secagem sobre a qualidade fisiológica e a armazenabilidade de sementes de cafeeiro. Os ensaios foram realizados nos Laboratórios de Análise de Sementes e de Técnicas Moleculares do Departamento de Agricultura da UFLA. Os frutos do cultivar Rubi foram colhidos, despolpados, e as sementes lavadas e deixadas sobre papel para retirada da água superficial. As sementes nos estádios verde cana e cereja foram submetidas à secagem convencional (à sombra) e à secagem em secador estacionário sob temperatura de 35ºC. Como testemunha foram analisadas sementes sem secagem. As avaliações foram feitas imediatamente após os tratamentos de secagem e após 4 e 8 meses de armazenamento. As sementes foram armazenadas a 10ºC em sacos plásticos impermeáveis. Foram realizados os testes de germinação, de protrusão radicular, de matéria seca de plântulas, de índice de velocidade de emergência, de condutividade elétrica, além de análises eletroforéticas de proteínas resistentes ao calor e da quantificação da atividade da enzima endo-ß-mananase. O delineamento foi inteiramente ao acaso em esquema fatorial dois (estádios de maturação) x três (métodos de secagem) x três (tempos de armazenamento), com quatro repetições. Sementes de cafeeiro colhidas no estádio cereja têm maior potencial de armazenamento que no estádio verde cana. Ocorre redução de germinação e vigor nas sementes de cafeeiro colhidas no estádio verde cana, quando submetidas à secagem rápida. A presença ou intensidade de bandas de proteínas resistentes ao calor está associada à secagem das sementes. Ocorre maior atividade da enzima endo-ß-mananase em sementes colhidas no estádio cereja que no estádio verde-cana. Ocorre aumento da atividade da enzima endo-ß-mananase durante o armazenamento.
Coffea arabica; tolerância à dessecação; armazenamento
The harvesting time and drying methods may influence the quality of coffee seeds during storage. This work was conducted with the objective of understand the effects of maturation stages and drying methods on the physiological quality and storability of coffee seeds. The experiments were performed at the Seed Analysis and Electrophoresis Laboratory of the Department of Agricultural of UFLA. The fruits of the cultivar Rubi were harvested, depulped and the mucilages removed by fermentation and the seeds washed and let on paper for removal of the water. The seeds were submitted to conventional drying (under shade) and drying in stationary drier at temperature of 35ºC. As a control, seeds with no drying were analyzed. The evaluations were done soon after the drying treatments and after four and eight months of storage. The seeds were stored at 10ºC in air-tight plastic packages. The germination test, radicle protrusion, seedling dry matter, emergence index speed, electric conductivity, electrophoresis analyses of heat-resistant proteins and quantification of endo-â-mannanase activity were the parameter evaluated. The design was completely randomized with 2 (maturation stages) x 3 (dryings) x 3 (storage time) factorial scheme with four replications. Seeds harvested in the cherry stage have greater storage potential than in the green-yellow stage. Reduced germination and vigor occur in the coffee seeds harvested in the green yellow stage, when submitted to fast drying. The presence or intensity heat-resistant protein bands is associated with seed drying. Greater activity of endo-ß-mananase enzyme occurs in seeds harvested in the cherry stage than in the green yellow stage. Increase of the activity of endo-ß-mananase enzyme occurs during storage.
Coffea arabica; desiccation tolerance; storage
Armazenabilidade de sementes de cafeeiro colhidas em diferentes estádios de maturação e submetidas a diferentes métodos de secagem1 1 parte da Dissertação de Mestrado apresentada pelo primeiro autor à UFLA, em 2005.
Storability of coffee seeds harvested at different maturation stages and submitted to different drying methods
André Delly VeigaI; Renato Mendes GuimarãesII; Sttela Dellyzete Veiga Franco da RosaIII; Édila Vilela de Resende Von PinhoII; Luiz Hildebrando de Castro e SilvaIV; Adriano Delly VeigaIV
IDoutorando Agronomia UFLA, Setor de Sementes/DAG, C.P. 37, Lavras, MG, CEP 37.200-000,adelly2@yahoo.com.br
IIProf. Dr. Ufla, renatomg@ufla.br, edila@ufla.br
IIIPesquisador Dr. Embrapa, sttelaveiga@ufla.br
IVGraduandos Agronomia UFLA
RESUMO
O momento da colheita e os métodos de secagem podem influenciar a qualidade das sementes de cafeeiro durante o armazenamento. Este trabalho foi realizado com o objetivo de avaliar os efeitos do estádio de maturação e do método de secagem sobre a qualidade fisiológica e a armazenabilidade de sementes de cafeeiro. Os ensaios foram realizados nos Laboratórios de Análise de Sementes e de Técnicas Moleculares do Departamento de Agricultura da UFLA. Os frutos do cultivar Rubi foram colhidos, despolpados, e as sementes lavadas e deixadas sobre papel para retirada da água superficial. As sementes nos estádios verde cana e cereja foram submetidas à secagem convencional (à sombra) e à secagem em secador estacionário sob temperatura de 35ºC. Como testemunha foram analisadas sementes sem secagem. As avaliações foram feitas imediatamente após os tratamentos de secagem e após 4 e 8 meses de armazenamento. As sementes foram armazenadas a 10ºC em sacos plásticos impermeáveis. Foram realizados os testes de germinação, de protrusão radicular, de matéria seca de plântulas, de índice de velocidade de emergência, de condutividade elétrica, além de análises eletroforéticas de proteínas resistentes ao calor e da quantificação da atividade da enzima endo-ß-mananase. O delineamento foi inteiramente ao acaso em esquema fatorial dois (estádios de maturação) x três (métodos de secagem) x três (tempos de armazenamento), com quatro repetições. Sementes de cafeeiro colhidas no estádio cereja têm maior potencial de armazenamento que no estádio verde cana. Ocorre redução de germinação e vigor nas sementes de cafeeiro colhidas no estádio verde cana, quando submetidas à secagem rápida. A presença ou intensidade de bandas de proteínas resistentes ao calor está associada à secagem das sementes. Ocorre maior atividade da enzima endo-ß-mananase em sementes colhidas no estádio cereja que no estádio verde-cana. Ocorre aumento da atividade da enzima endo-ß-mananase durante o armazenamento.
Termos para indexação:Coffea arabica, tolerância à dessecação, armazenamento.
ABSTRACT
The harvesting time and drying methods may influence the quality of coffee seeds during storage. This work was conducted with the objective of understand the effects of maturation stages and drying methods on the physiological quality and storability of coffee seeds. The experiments were performed at the Seed Analysis and Electrophoresis Laboratory of the Department of Agricultural of UFLA. The fruits of the cultivar Rubi were harvested, depulped and the mucilages removed by fermentation and the seeds washed and let on paper for removal of the water. The seeds were submitted to conventional drying (under shade) and drying in stationary drier at temperature of 35ºC. As a control, seeds with no drying were analyzed. The evaluations were done soon after the drying treatments and after four and eight months of storage. The seeds were stored at 10ºC in air-tight plastic packages. The germination test, radicle protrusion, seedling dry matter, emergence index speed, electric conductivity, electrophoresis analyses of heat-resistant proteins and quantification of endo-â-mannanase activity were the parameter evaluated. The design was completely randomized with 2 (maturation stages) x 3 (dryings) x 3 (storage time) factorial scheme with four replications. Seeds harvested in the cherry stage have greater storage potential than in the green-yellow stage. Reduced germination and vigor occur in the coffee seeds harvested in the green yellow stage, when submitted to fast drying. The presence or intensity heat-resistant protein bands is associated with seed drying. Greater activity of endo-ß-mananase enzyme occurs in seeds harvested in the cherry stage than in the green yellow stage. Increase of the activity of endo-ß-mananase enzyme occurs during storage.
Index terms:Coffea arabica, desiccation tolerance, storage.
INTRODUÇÃO
A causa da lenta e desuniforme germinação das sementes de cafeeiro pode estar relacionada ao estádio de maturação dos frutos, aos mecanismos que regem o processo de maturação e aos processos de pós-colheita, como secagem e armazenamento, fato já observado para outras espécies (Bewley e Black, 1994).
Sementes de cafeeiro possuem um alto conteúdo dos polissacarídeos celulose e hemicelulose (Wolfrom e Patin, 1964, citados por Silva, 2002), sendo que as principais hemiceluloses são mananas e galacto-mananas insolúveis, as quais possuem 2% de galactose (Bewley e Black, 1994). Segundo Silva (2002), estes polissacarídeos, os quais geralmente são depositados como fonte de reserva na semente, são degradados no momento da germinação pela ação de enzimas, incluindo endo-b-mananase, b-manosidase, galactosidase e celulase, resultando no enfraquecimento das paredes celulares do endosperma. Este pesquisador estudou os mecanismos e a regulação do processo de germinação nas sementes de cafeeiro e constatou que a atividade da enzima endo-b-mananase ocorreu primeiramente na região de protrusão da radícula e posteriormente nas demais regiões do endosperma. Esta atividade aumenta antes da protrusão da radícula; já a atividade da celulase ocorre em toda a região do endosperma.
Em sementes cuja germinação é limitada pela presença do endosperma, como as sementes de Coffea arabica L., há necessidade de amolecimento desse tecido para que haja a protrusão da radícula. Esse papel é desempenhado por várias enzimas, como, por exemplo, a endo-b-mananase, que está presente no endosperma em diferentes isoformas, sendo duas dessas inibidas pelo ácido abscísico na fase de enfraquecimento do endosperma na região próxima à radícula, inibindo a força de pressão da radícula (Silva et al., 2004).
Ao contrário do comportamento de sementes ortodoxas, que necessitam de redução do teor de água para que ocorra a germinação, sementes sensíveis à dessecação não passam pela secagem ao final da fase de maturação e, aparentemente, não adquirem completa tolerância à dessecação, provavelmente porque tais sementes iniciam a germinação logo após a maturação e nesta fase poderiam ser vistas já como plântulas em desenvolvimento, apresentando os eventos metabólicos associados à germinação (Farrant et al., 1988; Pammenter e Berjak, 1999).
Em várias pesquisas tem sido abordado que, para a obtenção de sementes de cafeeiro de melhor qualidade, é recomendada a colheita dos frutos no estádio cereja. No entanto, em outras tem-se observado que sementes de comportamento intermediário, completamente maduras, podem não apresentar o melhor potencial para o armazenamento (Finch-Savage, 1996), devido à possível iniciação do processo de germinação nos últimos estádios de desenvolvimento (Ellis et al., 1991; Hong e Ellis, 1992).
Sementes e organismos que toleram a desidratação apresentam algumas proteínas, cujas funções ainda não estão bem esclarecidas, mas sua estabilidade, sua alta afinidade com moléculas de água e abundância em organismos que toleram a desidratação, sugerem seu importante papel em aquisição de tolerância à dessecação (Blackman et al., 1991).
Estas proteínas, primeiramente descobertas em embriões de algodão, nomeadas proteínas lea, moduladas por ácido abscísico, acumulam em embriões de sementes de milho e cevada, durante os estádios mais tardios do desenvolvimento, em fase correspondente à aquisição de tolerância à dessecação (Bostock e Quatrano, citados por Leprince et al., 1993) e a sua expressão cessa rapidamente após a germinação (Blackman et al., 1991; Blackman et al., 1992). Segundo Thomann et al. (1992) e Rosa et al. (2004), proteínas lea também se acumulam em embriões de sementes de milho durante a lenta secagem, em fase correspondente à aquisição de tolerância à dessecação. Workers et al. (1998) observaram que embriões imaturos de milho, os quais adquirem tolerância à dessecação quando submetidos à lenta secagem, apresentam um padrão de proteínas semelhante às proteínas lea, presentes em sementes maduras e tolerantes à dessecação.
Diante do exposto, o presente trabalho foi realizado com o objetivo de avaliar a tolerância à dessecação e o potencial de armazenamento de sementes de cafeeiro (Coffea arabica L.) colhidas nos estádios verde cana e cereja.
MATERIAL E MÉTODOS
O presente trabalho foi desenvolvido nos Laboratórios de Análises de Sementes e de Técnicas Moleculares do Departamento de Agricultura da Universidade Federal de Lavras.
Foram colhidos frutos de Coffea arabica L., cultivar Rubi, no estádio cereja e verde cana os quais foram despolpados em despolpador manual e desmucilados por fermentação natural em água à 30ºC por 24 horas. As sementes foram lavadas e deixadas sobre papel para retirada do excesso da água superficial, antes de serem submetidas aos diferentes métodos de secagem. Neste momento, as sementes apresentavam teor de água médio de 53%.
As sementes foram submetidas a dois métodos de secagem: secagem convencional, à sombra, em ambiente de laboratório (secagem lenta) e secagem em secador estacionário (secagem rápida). Uma parte das sementes não foi submetida à secagem, sendo avaliada após a retirada da água superficial.
Na secagem à sombra, as sementes foram acondicionadas por 17 dias em bandejas metálicas forradas com papel e diariamente revolvidas, até atingirem teor de água de 12%.
Na secagem rápida foi empregado um secador estacionário experimental, de pequena escala, conforme modelo descrito por Navratil e Burris (1982). O secador foi regulado para funcionar na temperatura de 35ºC e fluxo de ar de aproximadamente 20m3.min-1.t-1. As sementes também foram secadas até teor de água de 12%.
As avaliações foram realizadas imediatamente após os tratamentos de secagem, e após quatro e oito meses de armazenamento sob temperatura de 10ºC e UR de 50%, em sacos plásticos impermeáveis. Para as avaliações da qualidade das sementes, foram realizados os testes de germinação, de protrusão radicular, de matéria seca de plântulas, de índice de velocidade de emergência e de condutividade elétrica. Foram também realizadas análises eletroforéticas de lea proteínas e quantificação da atividade da enzima endo-ß-mananase. Para estas análises foram reservadas sementes de cada tratamento, as quais foram primeiramente congeladas em submersão em nitrogênio líquido por 15 segundos e conservadas em deep-freezer a -86ºC, até o momento de utilização.
Teste de germinação - Realizado com quatro subamostras de 50 sementes sem pergaminho distribuídas em papel germitest umedecido com quantidade de água equivalente a duas vezes e meia o peso do substrato seco e colocadas para germinar à temperatura de 30ºC, na presença de luz. As avaliações foram realizadas aos trinta dias após a semeadura, segundo as Regras para Análise de Sementes (RAS) (Brasil, 1992) e os resultados expressos em porcentagem.
Protrusão radicular - Realizado aos quinze dias do início do teste de germinação, em que foram computadas as sementes que apresentavam emissão de radícula com, pelo menos, um milímetro de comprimento e os resultados foram expressos em porcentagem.
Matéria seca de plântulas - Realizado aos 30 dias de germinação, quando as plântulas destacadas dos endospermas remanescentes foram pesadas após secagem em estufa de circulação de ar sob temperatura de 65ºC até pesos constantes. Os resultados foram expressos em g.planta -1.
Índice de velocidade de emergência (IVE) - Realizado com quatro subamostras de 50 sementes distribuídas em caixas plásticas, contendo mistura de areia e terra, na proporção de 2:1, em câmara de crescimento a 30ºC. A irrigação foi realizada com freqüência de 2 em 2 dias e a quantidade de água necessária foi calculada para manter aproximadamente 70% da capacidade de campo do substrato. O IVE foi calculado segundo a fórmula proposta por Maguirre (1962) utilizando-se os resultados das avaliações diárias no teste de emergência, computando-se o número de plântulas emersas.
Teste de condutividade elétrica - Realizado com quatro subamostras de 25 sementes sem pergaminho, em copos plásticos contendo 75mL de água destilada. As sementes foram então colocadas para embeber em BOD, regulada para a temperatura de 25ºC, por período de 96 horas. As leituras foram realizadas em condutivímetro Digimed, modelo CD-21. Os resultados foram expressos em µmhos. cm-1.g de sementes-1.
Análise eletroforética de proteínas resistentes ao calor Para a extração das proteínas resistentes ao calor vinte sementes intactas de cada tratamento foram moídas em moinho refrigerado. Foram separados 200mg em microtubos, para aplicação de 300µL de tampão de extração (50mM tris-HCl-7,5; 500 mM NaCl; 5mM MgCl2; 1mM PMSF) e Antipain em proporção de 5mg para cada ml de tampão. Em seguida, os microtubos foram agitados em Vortex e levados para centrífuga a 14.000 g por 20 minutos a 4ºC. O sobrenadante foi então incubado em banho-maria a 85ºC por 15 minutos e novamente centrifugado por 30 minutos como acima referido. O sobrenadante foi vertido em microtubos e o pellet descartado. Antes da aplicação, os microtubos contendo 45mL do extrato de proteína + 23mL do tampão da amostra (5mL de glicerol, 2,5mL de solução tampão do gel concentrador, 2,5mg de azul de bromofenol, completando o volume para 25mL de água destilada) foram levados ao banho-maria em ebulição por 5 minutos. Foram, então, aplicados 40µL de cada amostra em gel de poliacrilamida SDS-PAGE a 12,5% (gel separador) e 6% (gel concentrador). A revelação para detecção das proteínas resistentes ao calor foi conduzida segundo metodologia descrita por Alfenas et al. (1991).
Atividade da enzima de endo-b-mananase - Para a extração da enzima endo-b-mananase foram moídas 10 sementes intactas de cada tratamento em moinho refrigerado. De cada tratamento foram pesados 200mg de cada material para adição de 600µL de tampão de extração (0,1 M Hepes/ 0,5M Nacl e ácido ascórbico (5mg de ácido ascórbico por ml de tampão), pH 8,0). Na etapa seguinte as amostras foram centrifugadas por 30 minutos a 10000g e 2µL do sobrenadante aplicados em gel contendo 6mL de LBG (Locust Bean Gum), 0,24 g de agarose e 24mL de tampão pH 5,0 (1M Ácido Cítrico/ 0,4M de Na2HPO4 2 H2O). As alíquotas foram aplicadas em furos de 2mm feitos no gel com auxílio de um furador O gel ficou incubado por 21h e revelado segundo metodologia proposta de Silva et al. (2004). A atividade da enzima endo-b-mananase foi calculada de acordo com Downie et al. (1994).
Procedimento estatístico - O delineamento experimental foi inteiramente casualizado, em esquema fatorial (2x3x3) com quatro repetições, sendo dois estádios de maturação dos frutos (verde cana e cereja), três métodos de secagem (sem secagem, secagem natural e secagem em secador) e três épocas de armazenamento (0, 4 e 8 meses). Para as comparações de médias foi utilizado o Teste de Scott Knott a 5% de probabilidade, transformando-se os dados de percentagem em [(arc sen X.100-1/2) + 0,5]. A análise dos dados foi realizada por meio do programa estatístico Sisvar (Ferreira, 2000).
RESULTADOS E DISCUSSÃO
De acordo com a Tabela 1, observa-se diferença significativa entre os valores de protrusão radicular em sementes colhidas nos diferentes estádios de maturação dos frutos, evidenciando a melhor qualidade fisiológica das sementes oriundas de frutos colhidos no estádio cereja. A protrusão radicular foi superior nas sementes secadas à sombra, no início e aos oito meses de armazenamento, quando comparado com a das sementes que foram secadas em secador e com as que não foram secadas, não tendo estas últimas diferido entre si (Tabela 2). Já aos quatro meses de armazenamento, as sementes que não foram secadas alcançaram protrusão radicular superior àquela observada naquelas sementes submetidas aos demais métodos de secagem, os quais não diferiram entre si.
Após oito meses de armazenamento, a menor protrusão radicular observada nas sementes não secadas em relação às sementes secadas à sombra, provavelmente seja devido às mudanças bioquímicas e metabólicas ocorridas em função do alto metabolismo que as sementes com alto teor de água (53%) desenvolveram e que as levaram a uma maior deterioração (Priestley, 1986 e Copeland e McDonald, 1995). Por ser uma secagem considerada mais branda, a secagem à sombra tenha proporcionado uma menor redução na protrusão radicular após 8 meses de armazenamento.
Pela Tabela 3, em sementes no estádio cereja, não houve diferenças significativas na germinação, independentemente do método de secagem e do período de armazenamento. Em sementes colhidas no estádio verde cana, nos períodos de zero e quatro meses, a menor germinação foi observada nas sementes secadas em secador, quando comparado à germinação das sementes secadas à sombra e das não submetidas à secagem, não tendo estas diferido entre si. Aos oito meses de armazenamento, a maior germinação foi observada nas sementes secadas à sombra e as menores nas sementes não secadas e nas secadas em secador.
Foi encontrada diferença significativa entre a germinação das sementes colhidas nos diferentes estádios de desenvolvimento do fruto (Tabela 3), para todos os tratamentos, com exceção para as sementes secadas à sombra e armazenadas por oito meses. Essa superioridade confirma os resultados de Guimarães (2000) que também observou maior germinação em sementes de cafeeiro colhidas em estádios mais avançados de maturação.
Pela Tabela 4 observa-se que, nas sementes que não foram armazenadas, houve diferença significativa entre os métodos de secagem tanto no estádio cereja quanto no estádio verde cana. No estádio cereja, a matéria seca de plântulas das sementes secadas em secador foi inferior à de plântulas de sementes não secadas e secadas à sombra, os quais não diferiram entre si. No estádio verde cana, a matéria seca de plântulas das sementes secadas à sombra foi superior àquelas encontradas para sementes secadas em secador e não secadas, não tendo estas diferindo entre si.
Aos quatro meses de armazenamento, no estádio cereja, a matéria seca de plântulas das sementes secadas em secador foi inferior à observada para as sementes não secadas que, as quais por sua vez, apresentaram valor menor que as sementes secadas à sombra. Nas sementes colhidas no estádio verde cana, a matéria seca de plântulas de sementes não secadas foi superior à observada para sementes secadas à sombra e em secador, não tendo estas diferido entre si.
Já nas sementes armazenadas por 8 meses, para ambos os estádios de maturação, a matéria seca de plântulas de sementes não secadas foi superior às encontradas para sementes secadas à sombra e em secador, não tendo estas diferido entre si.
Foi também observada maior matéria seca de plântulas em sementes no estádio cereja independente do método de secagem utilizado e do período de armazenamento, com exceção para o valor observado em sementes secadas à sombra, no início do armazenamento (Tabela 4), em que a matéria seca de plântulas das sementes no estádio cereja foi estatisticamente igual à das sementes verde cana. Também, de uma maneira geral, observa-se que a secagem em secador foi prejudicial ao desenvolvimento das plântulas, com reflexo na massa seca das mesmas.
Pela Tabela 5 nota-se que, nas sementes não armazenadas, foi observada diferença significativa entre os métodos de secagem, tanto no estádio cereja quanto no estádio verde cana. Em ambos os estádios de maturação dos frutos, o índice de velocidade de emergência das sementes secadas no secador foi inferior ao observado em sementes submetidas aos demais métodos de secagem. Brandão Júnior (2000) também observou redução do vigor em sementes de cafeeiro submetidas a esse método de secagem, o qual provocou intensas alterações, tais como formação de cristais, desaparecimento de vacúolo e de endomembranas, alterações estas observadas nas análises ultraestruturais.
Nas sementes colhidas no estádio cereja e armazenadas por 4 meses, não foi observada diferença significativa entre os métodos de secagem. No estádio verde cana, o índice de velocidade de emergência das sementes secadas no secador foram inferiores aos observados em sementes submetidas aos demais métodos de secagem.
Observa-se ainda nos resultados apresentados na Tabela 5, que menores IVE foram observados no início e aos oito meses de armazenamento, em sementes colhidas no estádio verde cana, secadas em secador e ainda naquelas não submetidas à secagem, no oitavo mês de armazenamento, quando comparadas às sementes colhidas no estádio cereja. Esses resultados provavelmente podem ser atribuídos à menor tolerância à dessecação de sementes colhidas no estádio verde cana, conforme também observado por Guimarães (2000).
Nas sementes que não foram submetidas à secagem não houve diferença significativa na condutividade elétrica em sementes colhidas nos diferentes estádios. Mas, naquelas secadas em secador e à sombra, menor condutividade foi observada em sementes colhidas no estádio cereja (Tabela 6).
Foi observada, ainda, maior condutividade elétrica em sementes colhidas no estádio verde cana e submetidas à secagem em secador. Já em sementes colhidas no estádio cereja não houve diferença significativa nessa variável quando as sementes foram submetidas aos diferentes métodos de secagem (Tabela 6). Pelos dados obtidos infere-se que sementes colhidas no estádio cereja já possuem um sistema de membranas mais organizado, o que parece proteger as sementes contra danos de secagem. Por outro lado, essa organização dos sistemas de membranas parece ser ainda deficiente em sementes colhidas no estádio verde cana.
Pela Tabela 6 observa-se também que a condutividade elétrica das sementes colhidas no estádio cereja foi inferior à observada para as colhidas no estádio verde cana. Esses resultados demonstram que a secagem, independente do método, pode causar danos aos sistemas de membranas celulares de sementes de cafeeiro, o que acontece de maneira diferenciada para cada estádio de desenvolvimento das sementes (Guimarães, 2000).
Pelos resultados das avaliações das sementes após o armazenamento, observa-se também que sementes colhidas no estádio cereja podem ser submetidas à secagem mais rápida, em secador, com menores perdas da germinação (Figura 1). Já as sementes colhidas no estádio verde cana são mais sensíveis a esse tipo de secagem, provavelmente, pelo fato de possuírem mecanismos de tolerância à dessecação menos desenvolvidos que as sementes no estádio cereja.
O desempenho fisiológico das sementes de cafeeiro colhidas nos estádios verde cana e cereja, observado durante o armazenamento, foi diferenciado em função dos métodos de secagem. Em geral, os valores de germinação e de vigor das sementes colhidas no estádio cereja foram superiores aos observados para as sementes colhidas no estádio verde cana. Essa relativa intolerância à dessecação das sementes de cafeeiro, quando colhidas em diferentes estádios de maturação, confirma o enquadramento dessas sementes em um grupo intermediário de classificação quanto à tolerância a dessecação (Ellis et al., 1990 e 1991; Hong e Ellis, 1995; Eira et al., 1999).
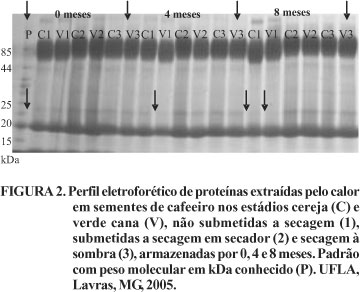
Pela Figura 2, pode-se observar, pelo perfil eletroforético de proteínas resistentes ao calor, menor intensidade ou até mesmo ausência da banda (indicada pelas setas), com peso molecular próximo a 25kDa, em sementes não submetidas à secagem, em todos os períodos de armazenamento. Como já observado em outras espécies, como o milho, a secagem parece induzir a expressão de alguns alelos, ocorrendo, dessa forma, o aparecimento ou intensificação de bandas (Faria, 2003). Um grupo de proteínas resistentes ao calor como as HSPs (Heat Shock Proteins), com baixo peso molecular pode se expressar sob condições de estresse (Vierling, 1991).
Trabalhando com sementes de cafeeiro colhidas nos estádios verde, verde cana e cereja, Guimarães (2000)observou presença de bandas de proteínas resistentes ao calor em estádios mais avançados de maturação. Além disso, o autor verificou que a ausência de determinadas bandas nas sementes sem secagem pode estar relacionada à intolerância à dessecação demonstrada por estas sementes. Na presente pesquisa não foi identificada no zimograma nenhuma banda que caracterizasse a menor intolerância das sementes de cafeeiro colhidas no estádio verde cana, quando comparado ao das sementes colhidas no estádio cereja.
A manutenção da qualidade fisiológica e a atividade da enzima endo-ß-mananase podem também estar relacionadas com a presença de proteínas resistentes ao calor nestas sementes que, segundo alguns autores, acumulam em embriões de sementes durante os estádios mais tardios do desenvolvimento, em fase correspondente à aquisição de tolerância à dessecação (Bostock e Quatrano, citados por Leprince et al., 1993).
Pelos resultados, observa-se que em sementes colhidas no estádio cereja existe maior atividade da enzima endo-â-mananase do que em sementes colhidas no estádio verde cana (Figura 3). As sementes colhidas no estádio cereja também apresentaram maiores valores de germinação e de vigor (Tabelas 3, 4 e 7 ).
Nota-se, ainda, que há um aumento gradativo da atividade da enzima endo-â-mananase à medida que se aumenta o tempo de armazenamento. Isto se deve ao fato da enzima endo-â-mananase estar envolvida na degradação de paredes na germinação das sementes de cafeeiro (Silva, 2002). No entanto, esta enzima é sintetizada na medida em que as sementes passam do processo de desenvolvimento para o processo de germinação nas sementes sem secagem. Portanto, por estarem mais adiantadas no processo de germinação, as sementes colhidas no estádio cereja, independente do tempo de armazenamento, apresentam maior atividade de endo-â-mananase, quando comparadas com a das sementes colhidas no estádio verde cana
Atividade considerável dessa enzima foi observada em sementes no estádio cereja secadas à sombra (Figura 3). A atividade da enzima nesta amostra deve-se, provavelmente, ao fato da secagem à sombra ter requerido um maior tempo, neste caso de 17 dias, possibilitando o avanço nessas sementes do processo de germinação. Esses resultados podem ser comparados com os observados nos testes de germinação e vigor dessas sementes (Tabelas 3 e 5).
CONCLUSÕES
Sementes de cafeeiro colhidas no estádio cereja têm maior potencial de armazenamento que no estádio verde cana.
Ocorre redução de germinação e vigor nas sementes de cafeeiro colhidas no estádio verde cana, quando submetidas à secagem rápida.
A presença ou intensidade de bandas de proteínas resistentes ao calor está associada à secagem das sementes.
Ocorre maior atividade da enzima endo-ß-mananase em sementes colhidas no estádio cereja que no estádio verde-cana.
Ocorre aumento da atividade da enzima endo-ß-mananase durante o armazenamento.
Submetido em 20/09/2005. Aceito para publicação em 28/11/2006.
trabalho financiado pela CAPES.
- ALFENAS, A.C.; PETERS, I.; BRUCE, W.; PASSADOS. G.C. Eletroforese de proteínas e isoenzimas de fungos e essências florestais Viçosa: UFV, 1991. 242 p.
- BEWLEY, J.D.; BLACK, M. Seeds: physiology of development and germination. New York: London Plenum Press, 1994. 367 p.
- BLACKMAN, S.A.; WETTLAUFER, S.H.; OBENDORF, R.L.; LEOPOLD, A.C. Maturation proteins associated with desiccation on tolerance in soybean. Plant Physiology, Rockville, v. 96, n. 3, p. 868-874. 1991.
- BLACKMAN, S.A.; WETTLAUFER, S.H.; OBENDORF, R.L.; LEOPOLD, A.C. Maturation proteins and sugars in desiccation tolerance of developing seeds. Plant Physiology, Rockville, v. 100, n. 1, p. 225-230, Sept. 1992.
- BRASIL. Ministério da Agricultura e Reforma Agrária. Secretaria Nacional de Defesa Agropecuária. Regras para análise de sementes Brasília, DF, 1992. 365 p.
- BRANDÃO JÚNIOR, D.da S. Marcadores da tolerância à dessecação de sementes de cafeeiro 2000. 144 p. Tese (Doutorado em Fitotecnia) Universidade Federal de Lavras, Lavras, MG.
- COPELAND, L.O.; McDONALD, M.B. Principles of seed science and technology 3. ed. Boston: KAP, 1995. 409 p.
- DOWNIE, B.; HILHORST, H.W.M.; BEWLEY, J.D. A new assay for quantifying endo-â-mananase activity using Congo Red dye. Phytochemistry, v. 36, p. 829-835, 1994.
- EIRA, M.T.S.; WALTERS, C.; CALDAS, L.S.; FAZUOLI, L.C.; SAMPAIO, J.B.; DIAS, M.C.L. L. Tolerance of coffea spp. seeds to desiccation and low temperature. Revista Brasileira de Fisiologia Vegetal, Londrina, v. 11, n. 2, p. 97-105, ago. 1999.
- ELLIS, R.H.; HONG, T.D.; ROBERTS, E.H. An intermediate category of seed storage behavior? I. Coffee. Journal of Experimental Botany, Ottawa, v. 41, n. 230, p. 1167-1174, Sept. 1990.
- ELLIS, R.H.; HONG, T.D.; ROBERTS, E.H. An intermediate category of seed storage behavior? II. Effects of provenance, immaturity, and imbibition on desiccation-tolerance in coffee. Journal of Experimental Botany, Ottawa, v. 42, n. 238, p. 653-657, May 1991.
- FARIA, M.A.V.R. Maturação de sementes de milho: aspectos físicos, bioquímicos e fisiológicos 2003. 129 p. Tese (Doutorado em Fitotecnia) Universidade Federal de Lavras, Lavras, MG.
- FARRANT, J. M.; PAMMENTER, N. W.; BERJAK, P. Recalcitrance a current assessment. Seed Science and Technology, Zurich, v. 16, n. 1, p. 155-166, 1988.
- FERREIRA, D. F. Análises estatísticas por meio do Sisvar para Windows Versão 4.0. In Reunião Anual da Região Brasileira da Sociedade Internacional de Biometria, 45., 2000, São Carlos-SP. Resumos... São Carlos-SP, 2000. p. 225-258.
- FINCH-SAVAGE, W. E. The role of developmental studies in research on recalcitrant and intermediate seeds. In: WORKSHOP ON IMPROVED METHODS FOR HANDLING AND STORAGE OF INTERMEDIATE/RECALCITRANT TROPICAL FOREST TREE SEEDS, 1995, Humlebaek, Denmark. Proceedings... Humlebaek, Denmark, 1996. p. 83-97.
- GUIMARÃES, R. M. Tolerância à dessecação e condicionamento fisiológico em sementes de cafeeiro (Coffea arabica, L.). 2000. 180 p. Tese (Doutorado em Fitotecnia) Universidade Federal de Lavras, Lavras, MG.
- HONG, T. D.; ELLIS, R. H. Development of desiccation tolerance in Norway maple (Acer platanoides L.) seeds during maturation drying. Seed Science Research, Wallington, v. 2, n. 2, p. 169-172, June 1992.
- HONG, T. D.; ELLIS, R. H. Interespecific variation in seed historage behaviour within two genera Coffea and Citrus. Seed science and Tecnology, Zurich, v. 23, n. 1 p. 165-181, 1995.
- LEPRINCE, O.; HENDRY, G. A. F.; MCKERSIE, B. D. The mehanisms of dessication tolerance in developing seeds. Seed Science Research, Wallingford, v. 3, n. 3,p. 231-246, Sept. 1993.
- MAGUIRRE, J. D. Speed of germination aid seedling emergence and vigor. Crop Science, Madison, v. 2, n. 2, p. 176-177, Mar./Apr. 1962.
- NAVRATIL, R. J.; BURRIS, J. S. Small-scale dryer designer. Agronomy Journal, Madison, v. 74, n. 1, p. 159-161, Jan./Feb. 1982.
- PAMMENTER, N. W.; BERJAK, P. A review of recalcitrant seed physiology in relation to dessication tolerance mechanisms, Seed Science Research, Wallingford, v. 9, n. 1, p. 13-37, Mar. 1999.
- PRIESTLEY, D. A. Seed Aging: implications of seed storage and persistence in the soil. Ithaca: Cornell University Press, 1986. 304 p.
- ROSA, S. D. V. F. da.; VON PINHO, E. V. R.; VIEIRA, M. G. G. C.; VEIGA, R. D. Indução de tolerância à alta temperatura de secagem em sementes de milho por meio de pré-condicionamento à baixa temperatura. Revista Brasileira de Milho e Sorgo, Sete Lagoas, v. 3, n. 2, p. 290-318, 2004.
- SILVA, E. A. A. da. Coffee (Coffee arabica L, cv Rubi) seed germination: mechanism and regulation 2002. 105 p. Thesis (Ph. D.) - Wageningen University, Wageningen.
- SILVA, E. A. A. da.; TOOROP, P. E.; VAN ELST, A. C.; HILHORST H. W. M. Abscisic acid controls embryo growth potential and endosperm cap weakening during coffee (Coffee arabica L., cv. Rubi) seed germination. Planta, Berlin, v. 220, n. 2, p. 251-261, Dec. 2004.
- THOMANN, E. B.; SOLLINGER, J.; WHITE, C.; RIVIN, C. J. Accumulation of group 3 late embryogenesis abundant proteins in Zea mays embryos. Roles of abscisic acid and the viviparous-1 gene product. Plant Physiology, Rockville, v. 99, n. 2, p. 607-614, June 1992.
- VIERLING, E. The role of heat shock proteins in plants. Annual Review of Plant Physiology and Plant Molecular Biology, Palo Alto, v. 42, p. 579-620, 1991.
- WOLKERS, W. F.; BOCHICCHIO, A.; SELVAGGI, G.; HOEKSTRA, F. A. Fourier transform infrared microscopy detects changes in protein secondary structure associated with desiccation tolerance in developing maize embryos. Plant Physiology, Rockville, v. 116, n. 3, p. 1169-1177, Mar. 1998.

Datas de Publicação
-
Publicação nesta coleção
22 Maio 2007 -
Data do Fascículo
Abr 2007
Histórico
-
Aceito
28 Nov 2006 -
Recebido
20 Set 2005