Life history and management of the tambaqui (Colossoma macropomum, Characidae): an important Amazonian food fish
Michael Goulding; Mírian Leal Carvalho
Museu Paraense Emílio Goeldi, Belém, Pará
INTRODUCTION
The genus Colossoma (Characidae) is widely distributed in South America, to which it is confined, and is known from the Orinoco (Mago Leccia, 1970), Amazon (Eigenmann, 1915), Bení-Mamoré-Guaporé (Pearson, 1937), Paraná-Paraguay-Pantanal (Britski, 1977) and La Plata (Ringuelet et al., 1967) drainage systems. The genus has not been adequately revised, but it appears to be represented by only two species in the Amazon Basin. The Amazonian species are much confused in the literature, but easily distinguished upon examination of live or dead specimens.
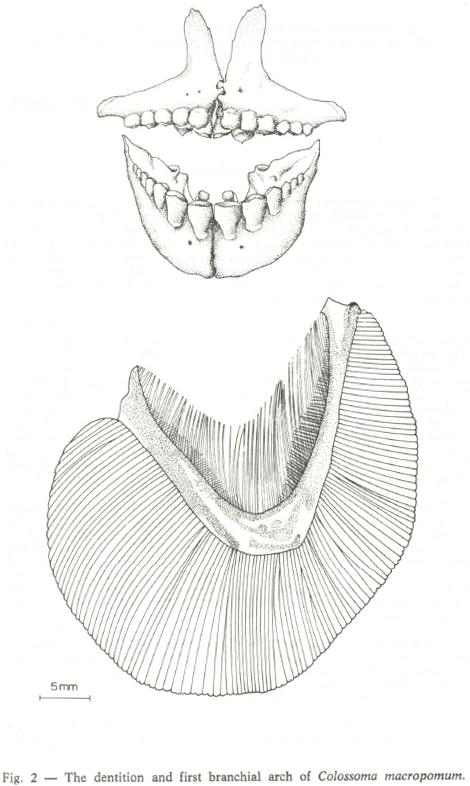
Colossoma macropomum is the second largest scaled fish (after Arapaima gigas, Osteoglossidae) in the Amazon Basin and reaches at least one meter in total length and 30 kg in weight (Fig. 1). The fish possesses the unique combination of molariform-like teeth adapted for crushing hard nuts and numerous elongated gillrakers that are employed to capture zooplankton (Fig. 2). Also noteworthy are its elongated opercula that support large fleshy flaps. The enlarged opercular appendages help increase water flow (with zooplankton) through the gills. Colossoma brachypomum is a smaller fish, but still reaches at least 90 cm in total length and 20 kg in weight. It is easily distinguished from C. macropomum by the triangular space between the first and second tooth rows in its premaxillae, its small gillrakers and the presence of teeth on the maxillary bones (C. macropomum has no maxillary teeth).
Young to pre-adult C. macropomum are ovoidal to rhomboidal shaped fishes, but during ontogeny into adults gradually become more elongated. In younger individuals (less than about 40 cm standard length) the cusps of the molariform-like teeth are higher and sharper than the lower and blunter projections of adults. Two conical teeth rest posteriorly against the symphyseal pair in the lower jaw, and give the impression of being supports for the front members. The premaxillae are endowed with two rows of teeth, and the four posterior elements - unlike in C. brachypomum - are set flush against the front members.
The gillrakers of C. macropomum range from about 85-100 in number on the first branchial arch. Each gillraker is endowed with microscopic ctenoid-like structures that alternate with one another on the internal side of each filament. Short, supplementary gillrakers reduce the space between the larger elements at the base of the branchial arches.
The stomach is well defined and very elastic in C. macropomum. Between the stomach and upper intestine are a larger number of pyloric caeca, whose function has yet to be determined. The intestines are relatively long and measure about 2.0 to 2.5 times the body length.
Colossoma macropomum displays distinct countershading, and is black dorsally and yellow to olive-green ventrally. The intensity of the hues is influenced by water transparency and color. In the blackwater, humic acid stained rivers, such as the Rio Negro, the fish is very dark, whereas in turbid water, such as the Rio Amazonas, it is much lighter in coloration and becomes almost a light golden-yellow ventrally.
Though the Amazon Basin has the most diverse freshwater fish fauna in the world, the life history of no species has been described from data based on field studies in the region. The present paper represents an attempt to describe and quantify as much as is known about the life history of C. macropomum, and more, to place the fish's ecology within an evolutionary and conservation framework.
STUDY AREA
The Amazon Basin
The Amazonian hydrographical network, the largest in the world, and embracing about six million km2, drains three main geological formations, namely the Andes, the Brazilian and Guiana Shields and the extensive lowlands between these (Fig. 3). Alfred Russel Wallace (1853) was the first naturalist to recognize the importance of the geological trio in the hydrochemistry and aquatic zoogeography of the Amazon Basin. Wallace (1853) described whitewaters, blackwaters and bluewaters (or clearwaters), and later Sioli (e. g. 1967, 1968), largely following this classification, clarified the relationship between geology and hydrochemistry in this region.
The major whitewater rivers include the Rio Solimões-Amazonas, Rio Madeira, Rio Purus and Rio Juruá. These rivers have headwaters in the eastern Andean slopes, and consequently they bring down huge quantities of silt that render them turbid and café-au-lait in color; hence they are called whitewater rivers. Secchi Disk readings in whitewater rivers are only a few centimeters as a rule, though the Rio Madeira, Rio Purus and Rio Juruá can clarify to about one meter transparency for a few weeks during the low water period. whitewater pH values taken in the channels range from slightly acid to about neutral. In comparison to the Amazonian Lowlands and Brazilian and Guiana Shields, the Andes have a complex mixture of rock types associated with much higher levels of basic salts (Gibbs, 1967). The best phytoplankton (Schmidt, 1973) and macrophyte (Junk, 1970) production in the Amazon Basin is found on the floodplains of the whitewater rivers and this is directly linked to nutrients of Andean origin.
Blackwater rivers for the most part rise in the Amazonian Lowlands, though many of them also have clearwater tributaries that drain the Guiana and Brazilian Shields. The humic acids that stain the Rio Negro, the largest blackwater river in the world, have been linked to low, open forest communities found on highly acidic white sands (Klinge & Ohle, 1964). In the so-called campina formations, many organic compounds are not decomposed in the soil litter and they are consequently leached into the streams and eventually the Rio Negro. The Rio Negro is extremely poor in nutrients, very acidic (pH about 4.4-4.9) and phytoplankton production is minimal (Sioli, 1967; Schmidt, 1973). Macrophyte communities in the Rio Negro and other blackwater rivers are poor in species in comparison to whitewater river floodplains and also much less extensive.
The clearwater rivers drain the Brazilian and Guiana Shields. The largest are the Rio Tocantins, Rio Xingu, Rio Tapajós and rightbank tributaries of the Rio Madeira. Clearwater rivers have a wide range of pH values, though they are generally very poor in nutrients with local exceptions (Sioli, 1967, 1968). Macrophyte communities are little developed and phytoplankton production, which has yet to be studied in detail, appears to be limited for the most part, judging from the lack of extensive blooms.
The three water types outlined above are not rigidly categorical, and many transitions exist. It should also be pointed out that the floodplains of turbid rivers (e.g. the Rio Solimões-Amazonas and Rio Madeira) have waterbodies that are filled with blackwaters and clearwaters draining the nearby terra firme. During the floods there is much mixing of water types on the whitewater river floodplains, and the hydrochemical interactions are complex and seasonal, and only recently are they being studied in any detail.
Rainfall in the Amazon Basin is neither temporally nor geographically distributed evenly, and thus all of the rivers do not rise and fall together. The different precipitation regimes of the northern and southern catchment basins interact to reduce the annual oscillation of the Rio Solimões-Amazonas, relative to its total discharge, in comparison to other large rivers of the world (Sternberg, 1975). In absolute terms, however, the annual oscillations are still striking. In the lower Rio Solimões there is about an 11 meter difference between the flood peak and low water trough but, as the floodplain becomes larger farther downstream near the mouth of the Rio Madeira, it is about 7 m (Fig. 4). Water level fluctuation in the lower courses of the tributaries of the Rio Solimões-Amazonas is controlled by the main river (except near the estuary where oceanic tides also have an influence). Water level data for Manaus (Fig. 4), for example, do not reflect the river level regime of the middle and upper Rio Negro, but of the Rio Solimões-Amazonas which dams back and extends the flooding season of the lower course of this large tributary. The southern affluents, such as the Rio Madeira, begin to fall earlier in the year than the main river, but their lower courses remain dammed back and thus display a more flattened and extended flood peak (Fig. 4).
Rainforest is the principal plant formation on Amazonian floodplains, and is found everywhere except in the lowest places where lakes and lagoons have developed, or where there has been deforestation. Floodplain rainforests are adapted to seasonal inundation. The length of time that riparian forests are inundated is determined by floodplain geomorphology and river level fluctuation in any particular year. The lower Rio Solimões and middle Rio Amazonas (near the mouth of the Rio Madeira) are inundated for about five to six months each year. In contrast, the Rio Madeira has a relatively high floodplain and for most of its length is only inundated for one to three months each year. The rightbank affluents of the Rio Madeira, however, have lower floodplains and their forest communities are inundated for about six months annually.
Study Sites
The investigation centers on the mid-western/central part of the Amazon Basin, with three main foci (see Fig. 3).
1) The lower Rio Solimões was investigated at Manaquiri, a floodplain area of about 15 km2 on the south side of the river, lying about 80 km upstream from the mouth of the Rio Negro (Fig. 5). This floodplain area contains two large, open waterbodies, namely Lago Manaquiri and Lago Grande; fish collections were made only in Lago Grande. The Manaquiri lakes are connected to the Rio Solimões by a 35 km long channel through which they are filled and drained with the seasonal oscillations of the principal river. At the height of the floods, Manaquiri waters meet those of adjacent floodplain areas, but during the low water period the lakes and lagoons become isolated except for their connections with the single channel that communicates with the river. As Rio Solimões water travels up the Manaquiri channel, it is largely decanted of its sediments, and thus the lake transparencies reach 1.7 m in comparison to the minimal ones of the highly turbid channel of the main river. Manaquiri lake waters are dark (but not black) in color, and pH is usually around 6.7. The Manaquiri floodplain is in constant ecological flux because of water level fluctuation. Especially notable are great biomass changes in macrophyte communities. During the low water period, wild rice (Oryza perennis) grows rapidly on the emerged lake and lagoon shores, forming verdant fields reminiscent of the domestic species in cultivation, but when water level rises with the floods, the plants are inundated and eventually become torn from the substrate by the winds (Fig. 6). Largely replacing rice at the time of the floods are much more extensive communities of floating herbaceous plants that develop very rapidly as the open waterbodies expand. The most abundant are the grasses Paspalum repens and Echinochloa polystachya. These species form dense floating meadows along with herbaceous taxa of several other families (Junk, 1970). Much of the floodplain rainforest at Manaquiri has been modified by agricultural activity and is in some stage of secondary succession. Both Alfred Russel Wallace (1853) and Richard Spruce (1908) visited Manaquiri, in 1850 and 1851, respectively, and though neither made reference to secondary growth at that time, their comments are highly suggestive. Wallace (1853) mentioned seeing cattle, sheep and cane fields at Manaquiri, while Spruce (1908) describes the local inhabitants gathering wild rice. Under natural conditions wild rice has a very limited extent on Amazonian floodplains, but appears to expand at the expense of deforestation or natural die-offs of shore vegetation.
Wallace (1853) stated that when he visited Manaquiri in May, June and July of 1850 that "the standing dish was tambaki," or the Colossoma macropomum of the present study. Interviews with the local fishermen indicated that the relatively heavy exploitation of C. macropomum at Manaquiri began in the 1950's in response to the growing Manaus market, but only larger individuals, or adults, were of commercial value and thus young fish were left almost unexploited. With shortages in the Manaus fish market in the late 1970's, young C. macropomum began to be exploited at Manaquiri. In summary, then, it may be said that both the floodplain rainforest and fish populations of Manaquiri have been modified by human action, and this is true of a large part of the Rio Solimões-Amazonas.
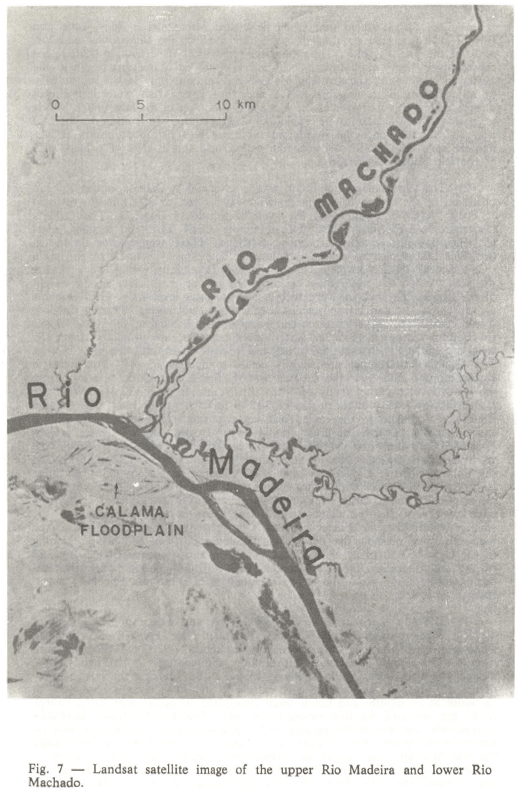
2) The Rio Madeira floodplain was investigated at Calama, about 700 km upstream of the mouth of the large whitewater river, or 100 km downstream of Porto Velho, the capital of the state of Rondônia (Fig. 7). The Calama floodplain is high but studded with numerous small lakes and lagoons, most of which are quasi-oxbows or linear in shape. These waterbodies lie in depressions that appear to be the remnants of abandoned channels of the main river. Secchi Disk transparencies of Calama floodplain waters vary during the year. When the waterbodies are invaded by the main river at the height of the floods, they become very turbid, but as water level falls, the sediments are decanted out and transparencies reach about two meters. During the low water period, the larger waterbodies become turbid because the wind mixes the bottom sediments. Smaller waterbodies, however, are protected from the wind by the high surrounding forest and remain clear until the new floods. The Calama waterbodies are slightly acidic to neutral in pH. For eight to eleven months each year the Calama waterbodies are landlocked, and during the lowest water period many of them shrink into large mudholes. With the floods, however, they attain depths of 7-12 m for a month or two. The Calama floodplain is linked to the main river via two small channels, about 10 km apart, that cut through the high alluvial levee. During the low water period, however, the connecting channels are dry and the floodplain is isolated from the river. Only in exceptionally high water years does the Rio Madeira invade the Calama floodplain over the high levee. The Calama floodplain rainforest is very high in stature with many species and, since there has been very little clearing in the area, large trees are found down to water's edge. There is a wide variety of floating plants, and during most of the year many of the smaller waterbodies are almost completely covered by a carpet of them, as the high rainforest protects the herbaceous communities from the wind. Fishing in the Calama floodplain is restricted mostly to subsistence activities because of the small size of the waterbodies, their difficult access (only by foot during most of the year) and dense populations of piranhas, especially Serrasalmus nattereri.
3) The Rio Machado is a clearwater river and enters the rightbank of the Rio Madeira opposite the Calama floodplain (see Fig. 7). Secchi Disk transparencies of the Rio Machado range from about 1.4-2.0 m, and pH values from 6.4 to slightly alkaline. Deforestation and erosion in the headwaters, along the Cuiabá-Porto Velho highway, appear to be reducing the transparency of the Rio Machado and may also be raising its pH. The river is about 400 m in width at its mouth, and has a small floodplain on either side for a stretch of 85 km, or until the first cataracts are met. The Rio Machado floodplain is relatively low and communicates with the main channel through numerous breaks in the levees. For a month or so, however, at the height of the floods, the Rio Machado rises above its levees and a continuous sheet of water is formed between the corresponding terra firme uplands, usually a distance of one to three km. The lakes and principal channel of the Rio Machado are almost devoid of the herbaceous plant communities associated with the whitewater rivers, though sedges colonize some beaches during the low water period. The floodplain rainforest of the Rio Machado is high in stature and little modified by human action. Commercial fishermen exploit the Rio Machado for the Porto Velho Market, and have done so since about 1959 (Goulding, 1979).
METHODS
Field observations of C. macropomum fisheries were made by the first author in July and September, 1973 and July, 1976 in the middle Rio Solimões and lower Rio Japurá region, and feeding habits, migrations and fisheries were investigated in detail in the Rio Madeira and Rio Machado between September, 1976 and February, 1981 (Goulding, 1970, 1980, 1981). Between 1976 and 1981 fish fauna surveys were also made of the Rio Madeira, Rio Machado, Rio Negro, Rio Solimões and Rio Tefé. The second author completed a detailed thesis on the feeding behavior of young C. macropomum and their interactions with zooplankton communities between April, 1979 and March, 1980 in the Manaquiri floodplain of the lower Rio Solimões (Carvalho, 1981). All together, year-round collections of C. macropomum were made at three different sites, namely the lower Rio Machado (1977 and 1978), the Calama floodplain of the upper Rio Madeira (1980) and the Manaquiri floodplain of the lower Rio Solimões (1979 and 1980), all of which are described above.
Gillnets were the principal gear used to capture C. macropomum in floodplain lakes and inundation forests. These were employed during both the day and night. Fishes examined from river channels were captured by commercial fishermen whom the first author accompanied to the middle Rio Madeira and lower Rio Japurá. In the river channels C. macropomum is captured mostly with seines, though these often have large meshes in which the fishes become entangled as in gillnets. Many specimens were also examined in the markets of Tefé (middle Rio Solimões), Manaus (lower Rio Negro) and Porto Velho (upper Rio Madeira), especially during the low water seasons when these fishes were being captured in the river channels.
The feeding behavior of C. macropomum was determined through stomach content analyses of 637 specimens ranging from 7-85 cm in standard length (taken from the tip of the snout to the end of the lateral line, which corresponds in this species with the approximate termination of the hypural bones). Three basic methods - occurrence, dominance and volume - were employed to evaluate the relative importance of the various items found in stomach contents. This analytical trio, or some variation of it, is standard procedure for studying the stomach contents of fishes (Hynes, 1950; Hyslop, 1980). Occurrence is a record of the number or individuals in which a particular food item appears. Dominance is calculated as the number of times a particular food item represented the largest part of the bulk. The volume of each food item was estimated on the basis of the total volume of an undistended stomach, when the stomach was less than full, or on the total quantity when the stomach was found distended. Colossoma macropomum has a fairly elastic stomach that can become quite distended, especially when larger individuals are feeding on fruits and seeds; however, it would have been too subjective to estimate the volumes of individual items in those specimens with less-than-full stomachs on the basis of a theoretically distended stomach. Estimates of the bulk of particular items were made on the basis of 1%, 5%, 10%, 25%, 50%, 75% or 100% of the total volume of the stomach as outlined above. Fruit and seeds eaten by C. macropomum were identified in the field (initially with the help of fishermen and rubber collectors), and then the tree, shrub or vine species from which they came was located in the flooded forest and herbarium specimens made for determination by professional taxonomists (Goulding, 1980). The contribution of different zooplankton taxa was determined by extracting 1 or 5 ml sub-samples (in solution) from the total bulk of this item in each stomach that contained it, and then counting the organisms under a microscope, at the lowest taxonomic level possible. The contribution of each zooplankton taxon to the total bulk was then extrapolated from these sub-samples.
In addition to the above three methods, we also measured mean fullness (based on the volumetric measurements outlined above, but including all specimens, even those with empty stomachs). For comparative purposes, the data were divided into the high and low water periods. The high water period was defined by the presence of inundated floodplain forest, and the complementary period as the low water season. This is more meaningful, ecologically, than selecting arbitrary points on a staff gauge measuring river level fluctuations in the channels.
MIGRATIONS AND DISTRIBUTION OF COLOSSOMA MACROPOMUM IN THE MID-WESTERN/CENTRAL AMAZON BASIN
Colossoma macropomum is one of about thirty or more species of Amazonian characins that migrate in the rivers in large schools. These migrations are well known to Amazonian fishermen and major fisheries are based on them (Goulding, 1979, 1980, 1981; Smith, 1979, 1981). Tagging experiments of Amazonian fishes have to date proved unsuccessful and migration information is from direct observation and fisheries data.
When water level begins to rise rapidly, large schools of C. macropomum are encountered moving upstream in the whitewater rivers, and at this time gonads are fully developed (Fig. 8). The exact spawning habitat of C. macropomum is still unknown, though it appears to be along the grassy levees that are being inundated with rising water. In the Rio Madeira region, the largest catches of C. macropomum are made when fishermen encounter the upstream moving schools just prior to the floods (Goulding, 1979, 1981). Subsequent to spawning, C. macropomum schools break up and the fishes begin entering floodplain forests of the blackwater, clearwater and whitewater rivers where they feed. Depending on floodplain morphology and water level, the fishes remain in flooded forest from four to seven months. When water level drops and the flooded forests are drained, most of the adult biomass of C. macropomum flees to the river channels, while much lesser quantities move into floodplain lakes. Populations that feed in the blackwater and clearwater river systems during the annual inundations migrate down these tributaries at the beginning of the low water period and subsequently enter whitewater rivers where they seek out woody shore areas. Likewise, populations that retreat from the floodplains of whitewater rivers also ensconce themselves in woody shore areas. Adult C. macropomum remain in these woody shore habitats until the commencement of the new floods, at which time they migrate upstream and spawn. The cycle is renewed when the spawned fishes migrate into the flooded forests to feed.
Our surveys of blackwater and clearwater tributaries revealed that immature C. macropomum are confined to the floodplains of whitewater rivers, though the exact number of years that the young reside in these nursery habitats before migrating is uncertain (occasionaly immature individuals are found in the floodplains of the lowermost reaches of some of the clearwater or blackwater rivers, but these fishes appear to be "strays" and to not prejudice our generalization). The smallest individuals that have been found with developed gonads were 56 cm (female, Rio Madeira) and 56 cm (female, Rio Solimões). It is still unclear whether young adults make their first migrations at the time of spawning or during the low water period.
Colossoma macropomum adults are widely distributed in the Amazon Basin, though rare or absent in the upper reaches of the larger blackwater and clearwater rivers. Judging from the rightbank tributaries of the Rio Madeira and the Rio Negro system, the species is only abundant in the clearwater and blackwater rivers within a distance of about 150 km from the whitewater rivers: beyond this, it is rare or disappears. The large characin is able to pass cataracts, and it has been witnessed by the first author negotiating the Teotônio rapids of the Rio Madeira above Porto Velho and the São Vicente rapids at the first turbulent stretch of the Rio Machado.
FEEDING BEHAVIOR
Manaquiri Floodplain, Rio Solimões
Of the 138 specimens (13-51 cm SL) examined from the high water period (March through August) at Manaquiri, only two had empty stomachs and mean fullness was 62 percent (Tab. 1). Fruits and seeds from trees or shrubs accounted for about 44 percent of the total bulk consumed by all of the specimens, followed by zooplankton with 30 percent and wild rice representing 22 percent. All together these three food items accounted for 96 percent of the total bulk of food eaten by the high water specimens. Of the 125 specimens examined from the low water period (September through February), 17 (14 percent) had completely empty stomachs and mean fullness dropped to 34 percent. Zooplankton accounted for 70 percent of the total bulk consumed, while not one of the other nine items eaten represented more than 10 percent.
The main species of fruits/seeds identified were Tabebuia barbata (Bignoniaceae), Mabea sp. (Euphorbiaceae), Cecropia sp. (Moraceae) and Vitex cymosa (Verbenaceae). All of these species are well represented in secondary vegetation of the Rio Solimões floodplain, and may be more abundant now than under more natural conditions when there was little human disturbance. Young C. macropomum appear to have a preference for the seeds of dry fruits. These fruits are usually capsules, such as Tabebuia barbata and Mabea spp., which dehisce and eject their seeds. Fleshy material must also be appreciated in the diet, however, as the seeds of fig syconia are so minute that most are not crushed during mastication and thus are ingested and defecated whole. Drupes, such as Vitex cymosa, with often sweet fleshy parts, are also ingested whole and the olive-sized seeds are not crushed, while in other species they are usually masticated before being swallowed. Perhaps the seeds of some fleshy fruits are toxic and thus the fishes avoid crushing them. The smallest individual that was found to have eaten fruits/seeds from trees or shrubs at Manaquiri was 19 cm SL, though specimens of less than 10 cm SL were captured in the floodplain forest of Janauacá, a site about 30 km downstream of the former. The minimum gillnet mesh-size used at Manaquiri was 9 cm (stretched), and thus C. macropomum individuals of less than about 13 cm were not susceptible to capture.
Because of deforestation wild rice (Oryza perennis) is undoubtedly more abundant in the Manaquiri floodplain than formerly when high forest was present around the margins. When water level rises rapidly at Manaquiri which, during the study period, was in late April and March, the rice fields growing on the low, open shores of the lakes, are flooded and young C. macropomum move into these areas to strip the seeds from the submerged stalks. Rice seeds were found in individuals ranging between 17-45 cm SL, though they were mostly dominant in specimens smaller than 30 cm SL.
Unlike tree fruits and rice seeds, zooplankton is available throughout the year for C. macropomum, and was the only food item that was important in both the high and low water periods. Using a theoretically 100 percent full stomach as the basis, then the mean contribution of zooplankton was 19 and 21 percent, respectively, during the high and low water periods at Manaquiri. Because fruits and seeds are largely unavailable during the low water period, this makes the relative contribution of zooplankton much higher than during the floods, even though about the same absolute quantities of the latter item were eaten during both periods. About 92 percent of the zooplankton consisted of cladoceran crustaceans, with Daphnia gessneri, Ceriodaphnia cornuta, Moina reticulata and Ceriodaphnia reticulata being the most important species (Tab. 2). The other eight percent was accounted for by copepods and ostracods. During the low water period Moina reticulata was the most important species eaten, followed by the copepod Notodiaptomus amazonicus, the latter of which were barely consumed during the floods.
To sum up for the year as a whole, 43 percent of the total bulk of food consumed by the immature specimens of C. macropomum captured at Manaquiri was represented by zooplankton, followed by 29 percent for tree fruits and seeds and 18 percent by rice seeds. This trio of food items, then, accounted for about 90 percent of the bulk of the diet of C. macropomum at this site.
Janauacá Floodplain, Rio Solimões
Janauacá is a large floodplain area downstream of and adjacent to Manaquiri (see Fig. 5). Janauacá is heavily fished for the Manaus market, and the local fishermen state that C. macropomum, until about 1970, was one of the most important species captured at the site. By 1976 it was relatively unimportant in the total catch recorded from Janauacá (Petrere, 1978). We include Janauacá here because it was the only site from which we obtained Colossoma macropomum specimens of 10 cm or less.
Five specimens ranging from 7-10 cm standard length were captured at the edge of a floating meadow in May, 1980. Filamentous algae was present in the stomachs of all five specimens, and represented the greatest volume (54 percent) of food ingested by the quintet (Tab. 3). Wild rice (Oryza perennis), zooplankton (Chydoridae and Ostracoda) and clam shrimps (Conchostraca) were each found in small amounts in four of the specimens. The mean fullness of the five specimens was 68 percent.
Calama Floodplain, Upper Rio Madeira
Of the 19 specimens (20-36 cm SL) examined from the high water period (March through mid-May) at Calama, none had empty stomachs and mean fullness was 41 percent (Tab. 4). Zooplankton and fruits/seeds accounted each for 42 percent of the total bulk consumed by all of the specimens, with about 4 items representing the remaining 16 percent. Of the 59 specimens examined from the extended low water period (mid-May through February), none had empty stomachs and mean fullness was 31 percent. Tree fruits/seeds accounted for 49 percent of the total bulk consumed by all of the low water specimens, followed by zooplankton with 24 percent and macrophyte remains with 17 percent; the remaining 10 percent was accounted for by about 8 items.
Although the Calama floodplain forest is inundated for only a short period of time (eight to ten weeks) each year, many fruits and seeds still fall into the waterbodies because of the large overhanging trees that completely surround the lake and lagoon margins. In floodplain areas that have been deforested and are in secondary succession, such as most of the lower Rio Solimões, the large shore trees are missing and thus there is little overhanging vegetation that would otherwise supply fruits and seeds to the waterbodies even before the onset of the floods. At Calama most of the floodplain fruit fall is during the rainy season, that is, between January and May. When the floodplain forest is inundated in late March and April, there is already much fruit on the ground, but it is unclear to what extent fishes may be able to find or utilize this. Most of the fruit material eaten by the Calama specimens was seeds and these were almost all masticated. The only two species that could be identified with certainty were Mabea sp. (Euphorbiaceae) and Hevea brasiliensis (Euphorbiaceae).
As at Manaquiri, Daphnia gessneri (Cladoura) was the most important zooplankton eaten by young C. macropomum at Calama during the high water period (Tab. 5). When the Rio Madeira invades the Calama floodplain the transparency of the waterbodies diminishes to a few centimeters because of the high turbidity of the main river. When water level falls and turbid water ceases to enter the floodplain, the sediments settle to the bottom and there are intensive plankton blooms (probably as a result of the introduction of nutrients in combination with relatively good transparencies, though these factors were not investigated at Calama). Daphnia gessneri is eaten throughout the year, but during the low water period is less important than Notodiaptomus spp. (Copepoda).
Macrophyte remains were eaten more at Calama than at Manaquiri, and this in part may be a reflection of the fact that the latter's waterbodies become largely covered with them during the lowest water period, the time of year when they were most consumed. Though the macrophyte remains could not be identified with any degree of confidence, many of them may have been from Eichhornia crassipes, which was the most abundant species in the Calama waterbodies investigated.
Floodplain Forest, Rio Machado
The high water period of the Rio Machado is between about mid-December and mid-June, and at this time of year adult C. macropomum migrate into the floodplain forests to feed on fruits and seeds (the following summary is from Goulding, 1980). Of the 96 specimens (44-75 cm SL) examined from the Rio Machado floodplain forests, only three had empty stomachs and mean fullness was 72 percent. Fruits and seeds accounted for about 94 percent of the total bulk of food eaten, with feces and fish representing the remaining four percent.
Colossoma macropomum select their favorite fruits and seeds from the floodplain flora within the sequence that they fall into the water during the inundation period. They have a preference for large seeds of both dry and fleshy fruits. Three examples will be discussed here. Rubber tree seeds (Hevea spruceana, Euphorbiaceae) alone accounted for 58 percent of the total fruit material eaten by the Rio Machado specimens. Hevea spruceana fruits are explosive capsules that, subsequent to maturation, burst open on hot days and eject their three or four seeds for several meters. The elongated, three to four cm seeds, have a hard nut wall, but this is easily broken by the powerful jaws and dentition of C. macropomum. The fishes wait beneath rubber trees whose seeds are falling into the water, and quickly snap the diaspores off the surface where they float. The second most important species found was a relatively large (1.5-2.0 cm diameter) palm fruit (Astrocaryum jauary) that has a very hard nut wall. Most of the A. jauary fruit consists of a single seed, and this is surrounded by a thin, yellow mesocarp and exocarp. Astrocaryum jauary seeds are heavy and sink to the bottom when they fall into the water. Colossoma macropomum, however, often grab them the moment they hit the water's surface, though they probably also forage them from the bottom. The seeds are almost always crushed and partly masticated, though occasionally entire fruits are ingested, in which case only the fleshy material is digested. Fruits of the genus Neolabatia (Sapotaceae) are relatively large (reaching about five to six cm in length) and are usually embedded in a very fleshy mesocarp that is sweet and edible by humans. Colossoma macropomum and other fishes rarely crush Neolabatia seeds but swallow them whole with the partially broken fleshy material. Since Neolabatia seeds could easily be crushed by the large characins, toxicity is suggested. Large fruits with small seeds (e. g. Genipa americana, Rubiaceae) are also eaten by C. macropomum adults, and in these cases most of the seeds pass through the digestive system without being destroyed. Amazonian fishes do not appear to be able to digest nut walls, and thus seeds swallowed whole are potentially dispersed, as has been shown through germination experiments (Goulding, 1980).
Floodplain Lakes of Rio Machado During the Low Water Period
When the floodplain forests of the Rio Machado are drained as river level falls, most of the C. macropomum population migrates downstream and into the Rio Madeira. Some individuals, however, are trapped in the floodplain lakes that become landlocked or connected to the main channel only by narrow channels that cut through the levees. Of the 27 specimens (55-79 cm SL) of C. macropomum captured in the Rio Machado floodplain lakes during the low water period, only one individual had more than a minute amount of food in its stomach, and in this case a characin (Curimata sp.) had been eaten. In the other specimens only traces of zooplankton, insects and fruits/seeds could be detected. The mean fullness of the low water specimens of the floodplain lakes of the Rio Machado was less than two percent.
River Channel, Rio Madeira
The low water period of the Rio Madeira is between about mid-June and mid-December, and at this time adult C. macropomum, that have been forced to migrate out of the floodplain forests, are found in the woody shore areas of, or migrating upstream in, the principal river. In September, 1977 the first author accompanied commercial fishermen exploiting the middle Rio Madeira and was able to examine 167 specimens (64-79 cm SL) captured in woody shore areas or while migrating upstream. About 75 percent of the specimens had completely empty stomachs, and mean fullness was less than one percent. Only very minute amounts of decomposing plant material, algae and occasional zooplankton were found in the stomachs that contained food.
Observations in Markets During Low Water Periods
During the low water periods between 1976 and 1981 the first author examined the stomachs of about 100 C. macropomum individuals that were for sale in the markets of Porto Velho, Humaitá (Rio Madeira) and Manaus. All of these fishes were reported to have been captured in the channels of whitewater rivers. Only one specimen was found with anything but the most minimal amount of food in its stomach, and in this case a small Curimata sp. (Curimatidae) had been swallowed whole.
EVOLUTIONARY ECOLOGY OF COLOSSOMA MACROPOMUM
The Amazon Basin, as we see it today, is a nutrient-poor ecosystem wherein have evolved biological mechanisms, especially in the rainforest root systems, that capture and recycle precious ions before they can be carried away to the streams and rivers. Communities of mycorrhizal fungi and root hyphae pick the nutrients out of the detritus, and it is this symbiotic sieve, recycling ions, that is thought to be largely responsible for allowing the rainforest to maintain its high biomass despite the poverty of basic salts in the underlying sediments and bedrock (e. g. Stark, 1971a, 1971b, 1972). The rainforest, in its floodplain counterpart, is directly linked to aquatic ecosystems and is a major, if not the main, supplier of food (fruits, seeds, insects and detritus) for many fish species. Floodplain rainforests are widely distributed in Amazonia and found in virtually every river system, and thus the nutrients cycled or recycled through them offer a very extensive contribution to the aquatic food chains. The other two principal sources of photosynthetic energy for Amazonian aquatic systems are phytoplankton and macrophyte communities. With a few exceptions, such as in the mouth-lakes of some of the clearwater rivers, high biomass development of phytoplankton and/or macrophyte communities is restricted to the floodplains of the whitewater rivers, or, in other words, to the fluvial geography of nutrients of Andean origin. In the following discussion we will focus our attention on C. macropomum as it revolves and has evolved around the trophic structure described above. Feeding behavior alone, of course, does not explain the evolutionary ecology of a fish it is the most convenient focal point in this case because the migratory and reproductive strategies fall more cleary into place within a trophic framework.
In the area considered in this paper (see Fig. 3) C. macropomum hatches are born mostly in the whitewater rivers at the commencement of the floods. We have no evidence of how long the larval fishes might remain in the river channels before entering floodplain areas, or how far they might be transported or migrate downstream; it is also remotely possible that larval fishes, at some stage soon after birth, are able to migrate short distances upstream in search of entrances to floodplains. The smallest individual that we have from a floodplain waterbody was seven cm in standard length. The birth of C. macropomum in the whitewater river channels, as opposed to other places, appears to have the following results. First, the newborn find themselves in close proximity to the whitewater river floodplain areas where they spend most of their immature lives and are nourished. Second, the river channels are much larger and deeper than floodplain waterbodies at the commencement of the floods, and thus egg and larval predation is probably decreased. In this view, there has been selection to spawn near, but not in, the whitewater river floodplain. As a consequence of this, it is hypothesized here that there is a general downstream displacement of larval fish soon after their birth, and we will return to this point later in the discussion.
The evidence presented earlier clearly shows that C. macropomum young are sustained on a diet consisting mostly of zooplankton and fruit material. Algae may also be important for larvae of less than 10 cm standard length, but more data are needed before this can be accepted. Even as pre-adults, C. macropomum is the largest known zooplanktivore in the Amazon Basin, and, of the characins, it appears to be the species that takes longest to reach maturity. We hypothesize that the zooplankton feeding adaptations of C. macropomum young have evolved - or have at least been maintained, if they arose for other reasons - in direct response to selection for the large size of the fish which gives it a special fruit and seed eating advantage as an adult (see below). Although at a very young age C. macropomum begin to eat fruits and seeds, these are not available in sufficient quantity throughout the year to sustain the nutritional requirements of the voracious young fishes, while zooplankton is in some abundance the year-round in the waterbodies receiving nutrients from the whitewater rivers* * Colossoma brachypomum, which lives sympatrically with C. macropomum, also reaches large size, but for most of its juvenile and all of its adult life does not feed on zooplankton. C. macropomum juveniles and adults feed heavily on the leaves of macrophytes and trees. The feeding behavior of the two species is distinctly different during the juvenile stages of their lives, but C. brachypomum will be dealt with in a later paper. . Zooplankton is not the only food item that is available throughout, or at least during most of, the year. Of the items known to be eaten by young C. macropomum, fish, insects/spiders, mollusks, crustaceans and macrophytes are present in some quantity during most or all of the year, but none of these is very important in the diet. Though young and adult C. macropomum occasionally eat fish, their relatively small mouths, heavy dentition and thick, fleshy lips, appear poorly adapted for piscivory, especially vis-à-vis the extremely diverse array of predatory fishes found in Amazonian waters. Insects and spiders are important in the diets of many Amazonian fishes (Knöppel, 1970, 1972; Almeida, 1980; Aragão, 1980; Goulding, 1980; Paixão, 1980; Soares, 1980), and there is undoubtedly much competition for them. The large size of C. macropomum would appear to be its main disadvantage for insect and spider eating on a larger scale than the data presented earlier revealed. Mollusks, crustaceans and bivalves are common in the floodplain waterbodies of whitewater rivers, though there are still too few fish species studied to know which taxa are mostly eating them (though the first author has examined doradid catfishes that had eaten large quantities of mollusks and sciaenids and siluroids that had fed heavily on shrimp). The ecological aspects of macrophyte grazing are very poorly understood in Amazonian waterbodies, though evidence has already been presented showing that the anostomid fishes Schizodon fasciatus, Rhytiodus argenteofuscus and R. microlepis (Santos, 1980) and the large doradid catfish Lithodoras dorsalis (Goulding, 1980) eat the roots and leaves of some aquatic herbaceous plants. Although almost no detailed information is available, the fact that fishes (and probably manatees as well) do not overgraze, or at least noticeably reduce the standing crop of floating meadows as a whole, suggests that the aquatic vertebrate herbivores are highly selective in the plant species and parts that are eaten and, in fact, that most of the macrophyte biomass may be of little value for direct consumption. Macrophyte-eating fishes may also require special physiological adaptations beyond those needed to digest fruits and seeds.
Approaching or reaching maturity (about 55 cm standard length?) a large, and we believe the largest, part of the young adult cohorts of C. macropomum migrate out of their nursery habitats in the waterbodies of the whitewater river floodplains, and with the floods, enter the nutrient-poor blackwater and clearwater tributaries where they then enter the floodplain forests to feed almost exclusively on fruits and seeds. There is no evidence to indicate whether the new recruits spawn in the same year that they migrate from their former nursery habitats, or only in the following year or two.
We hypothesize that the migrations of adult or nearly mature C. macropomum from the whitewater river floodplain nursery habitats to the rivers, subsequent to the annual floods, is directed towards two principal ecological strategies. First, the displacement of the larger size-classes away from the nursery habitats reduces intraspecific feeding competition for zooplankton during the low water period. The migration of adults to the rivers during the low water period is not absolute, as some older individuals do remain and in fact act like young fish in their feeding behavior, but much observation and fisheries data strongly suggest that these aberrant individuals represent only a very small part of the adult biomass. It is thus incorrect and misleading, in our view, to suggest, as Honda (1974) does, that zooplankton is important in the diet of adult C. macropomum (which she called C. bidens). Aberrant individuals may be important in particular communities, but they can give a very erroneous idea ot the larger life history of a taxon and the selective biophysical factors that have influenced the evolution of the species. Second, by migrating to the rivers after the floods, potential spawners do not run the risk of being cut off from breeding habitats by being trapped in landlocked waterbodies when the reproductive migrations begin later in the year at the commencement of the new floods.
Based on the evidence from the Rio Madeira, Rio Solimões-Amazonas and Rio Negro basins, there appear to be two principal geographical factors that influence the local distribution of adult C. macropomum in the mid-western/central part of the Amazon Basin, namely, the presence of floodplain forest for feeding and, in the case of the blackwater and clearwater tributaries, the distance from the whitewater rivers where the species breeds. As stated earlier, cataracts in themselves are not barriers to the distribution of C. macropomum, but may coincidentally mark the limits of the species because floodplain forests often become very restricted above rapids (as in the case of the rightbank tributaries of the Rio Madeira). In contrast to the rightbank tributaries of the Rio Madeira, whose first rapids lie within 50-200 km of the principal river, those of the Rio Negro are about 1,000 km distant (by way of the river) from the joining of the Rio Solimões and Rio Amazonas. The Rio Negro, however, has extensive areas of floodplain forest above the point (Rio Branco to Barcelos) where C. macropomum is no longer found, and our surveys showed that it is not replaced ecologically by another species (competitive exclusion can thus be excluded). The distribution of C. macropomum in the Rio Negro suggests that the species does not stray, even when floodplain forests are present, for more than about 200 km from whitewater rivers in the mid-western/central part of the Amazon Basin.
Adult C. macropomum turn to a diet consisting almost entirely of fruits and seeds in terms of bulk. Subsequent to spawning, and with the floods, the large characins migrate into the floodplain forests of all river types to feed on fruits and seeds. The inclusion of floodplain forests of the whitewater, blackwater and clearwater rivers in the trophic area of adults greatly increases their feeding range in comparison to immature fishes who are confined to the first. It is also safe to assume that a larger adult biomass can be sustained as consequence of being able to use the nutrient-poor river systems. In the floodplain forests size alone separates C. macropomum from all of the other frugivorous fishes, with the exception of C. brachypomum and one or two species of Brycon, that are able to crush large seeds, such as those of rubber trees (Hevea spruceana). The large size and powerful dentition of C. macropomum allow it to eat seeds that smaller frugivorous fishes cannot utilize, and this would appear to have been the main factor that selected for the robustness of this characin. Large size with a deep, thick body, and large internal cavity, has also made it possible for C. macropomum to store considerable quantities of fat during the period when it is in the floodplain forests feeding on fruits and seeds. The visceral fat reserves of C. macropomum account alone for about 10 percent of total body weight subsequent to the floods (Castelo, Amaya & Strong, 1980). Muscle and head fat represent at least another five percent (Castelo, pers. comum.). The laying down of large fat reserves is mainly an adaptation to the low water period when fruits and seeds are no longer available in quantity, but at which time considerable amounts of energy are needed for migratory and reproductive purposes.
As stated earlier, all the evidence indicates that subsequent to the floods most of the adult C. macropomum biomass migrates to the channels of the whitewater rivers where the fishes spend a large part of the low water period in woody shore areas. With the exception of the Rio Negro and upper Rio Japurá, all of the large rivers (Rio Solimões-Amazonas, Rio Madeira, Rio Purus, Rio Juruá and Rio Potomayo) of the western Amazon have banks built mostly from Andean alluvium. These rivers, of course, are the whitewater ones. As new banks are built up through time the floodplain rainforest colonizes them, but in the end, the soft, underlying alluvial foundations are undermined by the currents and eventually collapse, causing cave-ins with trees and all falling into the river. These bank cave-ins, or terras caídas as they are called in the Brazilian Amazon, have been noted by many naturalists (e.g. Bates, 1889) and are most common subsequent to the floods when river level is dropping and thus withdrawing the supporting wall of water. The Rio Madeira, in fact, literally means Wood River, and the name would be quite appropriate for other whitewater rivers because of the large amounts of trees that are brought downstream every year. Not all of the trees that fall into the river, or onto the collapsed banks, are transported downriver and it is these that form the woody shore area where adult C. macropomum reside during the low water period. The wood strewn banks of the whitewater rivers contrast sharply with those of the clearwater and blackwater rivers, whose banks are much more stable and less subject to undercutting and cave-ins. It is hypothesized here that the low-water retreat of C. macropomum to the whitewater river woody shore areas has evolved at least in part as a response to predation pressures.
During the low water period most of the blackwater and clearwater affluents are shallower than the large whitewater rivers into which they flow (the deep mouth-bay region of the lower Rio Negro is an exception), and, as mentioned above, they do not have extensive woody shore zones. The schooling fishes, of small to medium size, such as those in the genera Semaprochilodus, Curimata, Metynnis, Myleus and Hemiodus, appear to use shallow beach areas, especially at night, to escape predators, notably the large catfishes (Goulding, 1980). Colossoma macropomum, however, would be too large to move into these shallow beach areas to escape predators (though dolphins may be their most important predators). By migrating to the woody shore areas with their deeper waters, C. macropomum is offered protection from predators during the drawdown period.
From about one to two months before the floodplains are inundated, C. macropomum schools begin to quit the woody shore areas and then migrate upstream. The gonads are fully developed at this time. These upstream migrations counterbalance the displacement of larval fishes that will be carried downstream as a consequence of being born in or very near to the flowing river channels. The upstream migrations also serve to recruit adults into the nutrient-poor tributaries in the upper reaches of the whitewater rivers.
MANAGEMENT
Colossoma macropomum has been exploited commercially in the Amazon Basin since at least the end of the nineteenth century (Verissimo, 1895), but intensive and extensive fishing efforts aimed directly at the species appear to date only from the late 1960's when the region's urban centers began to grow rapidly and new gear became available. The traditional gear of C. macropomum fisheries consisted of fruit-baited trotlines, castnets, harpoons, small cotton seines and, by the 1930's, of bombs. By 1970, however, relatively cheap synthetic materials - especially nylon line or its equivalent - became available for constructing gillnets and better and larger seines, and these gear quickly replaced the more traditional methods for capturing the large characin in commercial operations.
Though no quantitative data are available, Arapaima gigas (Osteoglossidae), the pirarucu of the Brazilian Amazon, was probably the most important commercial fish species of inland Amazonia until about 1970, when it was replaced by C. macropomum.
Catch data of reasonable accuracy were collected for the first time in 1976, and in that year C. macropomum accounted for about 44 percent of the total catch of 30,800 tons landed at the principal market of Manaus (Petrere, 1978). Petrere's (1978) study monitored only fish that were consumed in Manaus and not exports (mostly of large catfishes), and thus C. macropomum actually represented somewhat less than 44 percent of the total catch of the Amazon's largest fishing fleet. Unpublished data collected by the Instituto Nacional de Pesquisas da Amazônia indicate that C. macropomum was still the most important species consumed in Manaus in 1980, but that catches had declined by about one-half since 1976.
Petrere (1978) showed that about 95 percent of the total gillnet catch arriving in the Manaus market was accounted for by C. macropomum, and that 80 percent of this species was taken with this gear. Colossoma macropomum received the brunt of gillnet effort in the 1970's because of its high market price, that constantly rose, and its robust size that allowed fishermen to use large meshes effectively. Small meshed gillnets were shunned because of the great damage that piranhas do to them and the large number of species that are often captured but of little market value. With declining catches in the late 1970's, commercial fishermen began to heavily attack young C. macropomum in the floodplain lakes of the whitewater rivers. As mentioned earlier, adolescent C. macropomum are larger than the adults of most other species, and thus have market value. Though the exploitation and sale of C. macropomum of less than 55 cm in length are prohibited by law, these fishes are still commonly sold in the many Manaus fish markets, especially those in the slums.
If the present, uncontrolled exploitation of C. macropomum continues for much longer, then it will become very difficult to detect and further elucidate the evolutionary ecology of this species, not to mention its disappearance as a food fish. The important question that begs an immediate answer is the following: In multi-species river and floodplain fisheries, as in Amazonia, is it possible to manage all of the species over the entire area, and, if not, which are the ones that should be protected to maintain sustained yields? We will not address the first part of this question in this paper, but cast a strong vote for C. macropomum as one of the major food fishes that needs and deserves protection.
If C. macropomum is decimated by the commercial fisheries, the most important trophic link between the fruits and seeds of the floodplain forests and human populations will also be destroyed. The only other species that could ecologically replace C. macropomum in eating the large fruits and seeds is its congener C. brachypomum, but this latter species has already been overexploited and is no longer very important in the commercial fisheries. The destruction of Colossoma populations, then, should be seen a double loss. First, overexploitation leads to commercial extinction, and second, with these fishes removed from the floodplain forest communities, a large quantity of fruit and seed energy will no longer be economically exploitable by man because there are no animals left to transform it into animal protein edible by humans.
Although direct exploitation is the major threat to C. macropomum at the present time, for completeness, the effects of floodplain deforestation and agricultural activities must also be discussed, as environmental destruction would probably have long-lasting and irreversible effects on C. macropomum and other fish populations. The data presented in this paper clearly show that both young and adult C. macropomum are dependent on the floodplain forests for fruits and seeds, and the evidence indicates that there are no alternative foods, other than perhaps zooplankton for immature individuals, that could in any significant way substitute these plant materials. The conclusion, then, is obvious: if the floodplain forests are destroyed, so too is the food source for the fruit and seed eating fishes.
The whitewater river floodplains are the main areas that are being eyed for large-scale agricultural activities, mainly because their soils are much better than those found along the blackwater and clearwater rivers. Let us assume, for the sake of argument, that young C. macropomum would be able to maintain large populations (with the aid of fish culturalists) in the whitewater river floodplain areas, to which they are restricted, even if these inundation forests were destroyed. Because of a lack of fruits and seeds, adults could no longer be nourished in the deforested floodplains of the whitewater rivers but, as the model we presented earlier suggests, mature fish would still be recruited into the less modified blackwater and clearwater river systems during the floods. Theoretically, then, large populations of C. macropomum could be maintained despite the destruction of the floodplain forests of whitewater rivers. Floodplain agriculture implies more than deforestation, however, and the principal threat to fish communities would be the herbicides, insecticides and other poisons that would undoubtedly be needed to control pests and weeds. Floodplain waterbodies would be especially susceptible to toxins during the low water period; when they become shallow and restricted in size, and fish and other animal populations are densely packed into them. It is not far-fetched to think that floodplain grain farmers would even end up using piscicides to kill rice-eating fishes, such as young C. macropomum, that might get into cultivated fields and attack crops before they matured and could be harvested.
An effective management and conservation program for the Amazon fish resource cannot be based on a single species, but must include entire animal and plant communities over large geographical areas, as the ichthyofauna is extremely diverse and many taxa, such as C. macropomum, have life histories that embrace immense areas in which several types of habitats are used. The South American countries that are the political inheritors of Amazonia have yet to take their fish resources seriously in the sense of implementing sound research, management and conservation programs, and yearly the problems become more exacerbated because of an exploding urban population that anachronistically and pathetically depends on precarious fisheries anchored to an extractive economy. For Amazonian fisheries, uncontrolled exploitation is ultimately a synonym for commercial extinction. The great Brazilian literary naturalist, José Veríssimo, perspicaciously sounded the alarm against the dangerous course on which Amazonian fisheries had already embarked in the nineteenth century (Veríssimo, 1895). His warning is yet to be fully heard though high fish prices (supply not satisfying demand) and shortages are malevolenty clearing the economic air for the final note that will announce serious protein shortages. At the end of the nineteenth century, Veríssimo (1895) tells us, tambaqui, the fish of the present investigation, was brought to the Manaus market in such excessive quantities that much of it could not be sold and surplus charitably found its way into the kitchen of the booming city's prison. When the curtains go down on the twentieth century, it is highly unlikely, unless major steps are taken immediately, that the free citizens of Manaus will be able to enjoy the large fruit eating fish that once graced the menu of the incarcerated.
ACKNOWLEDGEMENTS
The research for this paper was financed by the Instituto Nacional de Pesquisas da Amazônia (INPA) and World Wildlife Fund. Dr. Rosemary Lowe-McConnell is thanked for criticizing the manuscript. Mr. George Nakamura and Sra. Graça Overal are thanked for typing the manuscripts.
REFERENCES
Almeida, R. G., 1980. Aspectos taxonômicos e hábitos alimentares de três espécies de Triportheus (Pisces: Characoidei. Characidae), do Lago do Castanho, Amazonas. Master's Thesis. Instituto Nacional de Pesquisas da Amazônia, Manaus.
Aragão, L. P., 1980. Desenvolvimento embrionário e larval, alimentação e reprodução do aruanã, Osteoglossum bicirrhosum Vandelli, 1829, do Lago Ianauacá - Amazonas, Brasil. Master's Thesis. Instituto Nacional de Pesquisas da Amazônia, Manaus.
Britski, H. A., 1977. Sobre o gênero Colossoma (Pisces, Characidae). Ciênc. Cult., S Paulo 29:810.
Carvalho, M. L., 1981. Alimentação do tambaqui jovem (Colossoma macropomum Cuvier, 1818) e sua relação com a comunidade zooplanctônica do Lago Grande-Manaquiri, Solimões-AM. Master's Thesis. Instituto Nacional de Pesquisas da Amazônia, Manaus.
Castelo, F. P., D. R. Amaya & F. C. Strong, 1980. Aproveitamento e características da gordura cavitária do tambaqui, Colossoma macropomu. Acta amaz. 10 (3):557-576.
Eigenmann, C. H., 1915. The Serrasalminae and Mylinae. Ann. Carneg. Mus. 9:225-272.
Gibbs, R. J., 1967. The geochemistry of the Amazon river system. Part I. The factors that control the salinity and the composition and concentration of the suspended solids. Bull. geol. Soc. Am. 78:1203-1232.
Goulding, M., 1979. Ecologia da pesca do Rio Madeira. Conselho Nacional de Desenvolvimento Científico e Tecnológico, Instituto Nacional de Pesquisas da Amazônia, Manaus.
Goulding, M., 1980. The fishes and the forest. University of California Press, Los Angeles.
Goulding, M., 1981. Man and fisheries on an Amazon frontier. Junk Publishers, The Hague.
Honda, E. M. S., 1974. Contribuição ao conhecimento da biologia de peixes do Amazonas. II. Alimentação do tambaqui, Colossoma bidens (Spix). Acta amaz. 4:47-53.
Hynes, H. B. N., 1950. The food of fresh-water sticklebacks (Gasterosteus aculeatus and Pygosteus pungitius) with a review of methods used in studies of the food of fishes. J. Anim. Ecol. 19:35-38.
Hyslop, E. J., 1980. Stomach contents analysis - A review of methods and their application. J. Fish Biol. 17:411-429.
Junk, W. J., 1970. Investigations on the ecology and production-biology of the "floating meadows" (Paspalo-Echinochloetum) on the Middle Amazon. Part 1: The floating vegetation and its ecology. Amazoniana 2:449-495.
Klinge, H. & W. Ohle, 1964. Chemical properties of rivers in the Amazonian area in relation to soil conditions. Verh. int. Verein. theor. angew. Limnol. 15:1067-1076.
Knoppel, Hans-Armin, 1970. Food of Central Amazonian fishes: Contribution to the nutrient-ecology of Amazonian rain-forest streams. Amazoniana 2:257-352.
Knoppel, Hans-Armin, 1972. Zur Nahrung tropischer Süsswasserfische aus Südamerika. Ibid. 3:231-246.
Mago Leccia, F., 1970. Lista de los peces de Venezuela. Ministerio de Agricultura y Cría, Oficina Nacional de Pesca, Caracas.
Paixão, I. M. P., 1980. Estudo da alimentação e reprodução de Mylossoma duriventris Cuvier, 1818 (Pisces, Characoides), do Lago Janauacá, Am, Brasil. Master's Thesis, Instituto Nacional de Pesquisas da Amazônia, Manaus.
Pearson, N. E., 1937. The fishes of the Beni-Mamoré and Paraguay basins and a discussion of the origin of the Paraguayan fauna. Proc. Calif. Acad. Sci. 23:99-114.
Petrere Jr., M., 1978. Pesca e esforço de pesca no Estado do Amazonas. II. Locais de pesca, aparelhos de captura e estatísticas de desembarque. Acta amaz. 8 (Suppl. 2).
Ringuelet, R., R. Aramburu & A. A. Aramburu, 1967. Los peces argentinos de agua dulce. Comision de Investigacion Científica, Buenos Aires.
Santos, G. M., 1980. Estudo de reprodução e hábitos reprodutivos de Schizodon fasciatus, Rhytiodus microlepis e Rhytiodus argenteofuscus (Pisces, Anostomidae) do Lago Janauacá. Acta amaz. 10 (2):391-400.
Schmidt, G. W., 1973. Primary production of phytoplankton in the three types of Amazonian waters. III. Primary productivity of phytoplankton in a tropical flood plain lake of Central Amazonia, Lago do Castanho, Amazonas, Brazil. Amazoniana 4:379-404.
Schmidt, G. W., 1976. Idem. IV. On the primary productivity of phytoplankton in a bay of the Lower Rio Negro (Amazonas, Brazil). Ibid. 5:517-543.
Sioli, H., 1967. Studies in Amazonian waters. Atas Simp. Biota amaz. 3:9-50.
Sioli, H., 1968. Hydrochemistry and geology in the Brazilian Amazon region. Amazoniana 1 (3):267-277.
Smith, N. J. H., 1979. Pesca no Rio Amazonas. Conselho Nacional de Desenvolvimento Científico e Tecnológico, Instituto Nacional de Pesquisas da Amazônia, Manaus.
Smith, N. J. H., 1981. Man, fishes and the Amazon. Columbia University Press, New York.
Soares, M. G. M., 1979. Aspectos ecológicos (alimentação e reprodução) dos peixes do Igarapé do Porto, Aripuanã, MT. Acta amaz. 9 (2):325-352.
Spruce, R., 1908. Notes of a botanist on the Amazon and Andes, 2 vols. Macmillan, London.
Stark, N., 1971a. Nutrient distribution in some Amazonian soils. Trop. Ecol. 12 (1):24-50.
Stark, N., 1971b. Nutrient recycling. II: Nutrient distribution in Amazonian vegetation. Ibid. 12 (2):177-201.
Stark, N., 1972. Nutrient cycling pathways and litter fungi. Bioscience 22 (6):355-360.
Sternberg, H. O., 1975. The Amazon river of Brazil. Erdk. Wissen 40.
Verissimo, J., 1895. A pesca na Amazônia. Livraria Classica de Alves, Rio de Janeiro.
Wallace, A. R., 1853. A narrative of travels on the Amazon and Rio Negro. Reeve, London.
- Almeida, R. G., 1980. Aspectos taxonômicos e hábitos alimentares de três espécies de Triportheus (Pisces: Characoidei. Characidae), do Lago do Castanho, Amazonas. Master's Thesis. Instituto Nacional de Pesquisas da Amazônia, Manaus.
- Aragão, L. P., 1980. Desenvolvimento embrionário e larval, alimentação e reprodução do aruanã, Osteoglossum bicirrhosum Vandelli, 1829, do Lago Ianauacá - Amazonas, Brasil. Master's Thesis. Instituto Nacional de Pesquisas da Amazônia, Manaus.
- Britski, H. A., 1977. Sobre o gênero Colossoma (Pisces, Characidae). Ciênc. Cult., S Paulo 29:810.
- Carvalho, M. L., 1981. Alimentação do tambaqui jovem (Colossoma macropomum Cuvier, 1818) e sua relação com a comunidade zooplanctônica do Lago Grande-Manaquiri, Solimões-AM. Master's Thesis. Instituto Nacional de Pesquisas da Amazônia, Manaus.
- Castelo, F. P., D. R. Amaya & F. C. Strong, 1980. Aproveitamento e características da gordura cavitária do tambaqui, Colossoma macropomu. Acta amaz. 10 (3):557-576.
- Eigenmann, C. H., 1915. The Serrasalminae and Mylinae. Ann. Carneg. Mus. 9:225-272.
- Gibbs, R. J., 1967. The geochemistry of the Amazon river system. Part I. The factors that control the salinity and the composition and concentration of the suspended solids. Bull. geol. Soc. Am. 78:1203-1232.
- Goulding, M., 1979. Ecologia da pesca do Rio Madeira. Conselho Nacional de Desenvolvimento Científico e Tecnológico, Instituto Nacional de Pesquisas da Amazônia, Manaus.
- Goulding, M., 1980. The fishes and the forest. University of California Press, Los Angeles.
- Goulding, M., 1981. Man and fisheries on an Amazon frontier. Junk Publishers, The Hague.
- Honda, E. M. S., 1974. Contribuição ao conhecimento da biologia de peixes do Amazonas. II. Alimentação do tambaqui, Colossoma bidens (Spix). Acta amaz. 4:47-53.
- Hynes, H. B. N., 1950. The food of fresh-water sticklebacks (Gasterosteus aculeatus and Pygosteus pungitius) with a review of methods used in studies of the food of fishes. J. Anim. Ecol. 19:35-38.
- Hyslop, E. J., 1980. Stomach contents analysis - A review of methods and their application. J. Fish Biol. 17:411-429.
- Junk, W. J., 1970. Investigations on the ecology and production-biology of the "floating meadows" (Paspalo-Echinochloetum) on the Middle Amazon. Part 1: The floating vegetation and its ecology. Amazoniana 2:449-495.
- Klinge, H. & W. Ohle, 1964. Chemical properties of rivers in the Amazonian area in relation to soil conditions. Verh. int. Verein. theor. angew. Limnol. 15:1067-1076.
- Knoppel, Hans-Armin, 1970. Food of Central Amazonian fishes: Contribution to the nutrient-ecology of Amazonian rain-forest streams. Amazoniana 2:257-352.
- Mago Leccia, F., 1970. Lista de los peces de Venezuela. Ministerio de Agricultura y Cría, Oficina Nacional de Pesca, Caracas.
- Paixão, I. M. P., 1980. Estudo da alimentação e reprodução de Mylossoma duriventris Cuvier, 1818 (Pisces, Characoides), do Lago Janauacá, Am, Brasil. Master's Thesis, Instituto Nacional de Pesquisas da Amazônia, Manaus.
- Pearson, N. E., 1937. The fishes of the Beni-Mamoré and Paraguay basins and a discussion of the origin of the Paraguayan fauna. Proc. Calif. Acad. Sci. 23:99-114.
- Petrere Jr., M., 1978. Pesca e esforço de pesca no Estado do Amazonas. II. Locais de pesca, aparelhos de captura e estatísticas de desembarque. Acta amaz. 8 (Suppl. 2).
- Ringuelet, R., R. Aramburu & A. A. Aramburu, 1967. Los peces argentinos de agua dulce. Comision de Investigacion Científica, Buenos Aires.
- Santos, G. M., 1980. Estudo de reprodução e hábitos reprodutivos de Schizodon fasciatus, Rhytiodus microlepis e Rhytiodus argenteofuscus (Pisces, Anostomidae) do Lago Janauacá. Acta amaz. 10 (2):391-400.
- Schmidt, G. W., 1973. Primary production of phytoplankton in the three types of Amazonian waters. III. Primary productivity of phytoplankton in a tropical flood plain lake of Central Amazonia, Lago do Castanho, Amazonas, Brazil. Amazoniana 4:379-404.
- Sioli, H., 1967. Studies in Amazonian waters. Atas Simp. Biota amaz. 3:9-50.
- Sioli, H., 1968. Hydrochemistry and geology in the Brazilian Amazon region. Amazoniana 1 (3):267-277.
- Smith, N. J. H., 1979. Pesca no Rio Amazonas. Conselho Nacional de Desenvolvimento Científico e Tecnológico, Instituto Nacional de Pesquisas da Amazônia, Manaus.
- Smith, N. J. H., 1981. Man, fishes and the Amazon. Columbia University Press, New York.
- Soares, M. G. M., 1979. Aspectos ecológicos (alimentação e reprodução) dos peixes do Igarapé do Porto, Aripuanã, MT. Acta amaz. 9 (2):325-352.
- Spruce, R., 1908. Notes of a botanist on the Amazon and Andes, 2 vols. Macmillan, London.
- Stark, N., 1971a. Nutrient distribution in some Amazonian soils. Trop. Ecol. 12 (1):24-50.
- Stark, N., 1972. Nutrient cycling pathways and litter fungi. Bioscience 22 (6):355-360.
- Sternberg, H. O., 1975. The Amazon river of Brazil. Erdk. Wissen 40.
- Verissimo, J., 1895. A pesca na Amazônia. Livraria Classica de Alves, Rio de Janeiro.
- Wallace, A. R., 1853. A narrative of travels on the Amazon and Rio Negro. Reeve, London.
Publication Dates
-
Publication in this collection
28 Aug 2009 -
Date of issue
1982