Resumos
Avaliou-se o efeito da suplementação de dois níveis de proteína bruta (PB), 14 e 16%, e dois de vitamina E (VE), 25 e 250mg/kg, na dieta sobre o desempenho e composição do ovo de matrizes de frangos de corte, entre 24 e 54 semanas de idade e sobre o desempenho de suas progênies. Foram alojadas 480 fêmeas e 48 machos da linhagem Ross, com 18 semanas de idade, em 16 boxes com 30 fêmeas e 3 machos em cada um. As dietas foram à base de milho e farelo de soja, sendo todas isocalóricas (2750kcal de EM/kg). O delineamento experimental foi o inteiramente ao acaso, constituído de quatro tratamentos em esquema fatorial 2×2, sendo dois níveis de PB (NPB) e dois níveis de VE (NVE), representados por quatro repetições cada um. A produção de ovos/ave/dia, o número de ovos/ave-alojada, o peso do ovo, a viabilidade, o peso corporal das fêmeas no final do experimento e a percentagem de gema no ovo não foram influenciados (P>0,05) pelos tratamentos e nem pela interação entre NVE × NPB. Menor percentagem de casca (P<0,05) e maior percentagem de albúmen (P<0,05) foram obtidos em ovos de aves que receberam o maior NPB na dieta, independente do NVE suplementado na dieta. Maior concentração de alfa-tocoferol na gema foi obtida em ovos de matrizes que receberam dieta suplementada com 250mg de VE/kg (P<0,05). O rendimento de incubação em ambas as idades estudadas (32ª e 52ª semanas) resultou em efeitos não significativos (P>0,05) dos NPB e dos NVE, representados pelo peso do ovo incubado, pela taxa de eclosão, pela razão entre o peso do ovo e o peso da progênie e pelo desempenho da progênie até a idade de abate. Porém, maior peso ao nascer (P<0,01) foi encontrado na progênie proveniente de reprodutoras com 32 semanas de idade, que receberam dieta com maior NPB e maior NVE. Dieta com 14% de PB e suplementada com 25mg de VE/kg é suficiente para assegurar o desempenho produtivo e reprodutivo satisfatório de matrizes de frangos de corte durante a fase de produção.
Matriz pesada; proteína bruta; vitamina E; produção de ovos; composição do ovo
This study was carried out with the objective of evaluating the effects of supplementation of two levels of crude protein (14 and 16% CP) and vitamin E (25 and 250mg VE/kg) in the diet on the performance and egg composition of broiler breed hens and on the performance of their progeny during the laying period (24-to-54 weeks of age). Thus 480 female and 48 male Ross breeder 18-week-old birds were randomly distributed in 16 floor pens, with 30 females and three males in each one. The diets were based on corn and soybean meal, and all them were isoenergetics (2750kcal ME/kg). The experimental design was a complete randomized block and the treatments were arranged in 2 × 2 factorial (CP × VE levels) with two repetitions each. Egg production/hen/day, egg weight, egg number/hen housed, viability, final female body weight and egg yolk percentage were not affected (P>0.05) by the treatments, and no interaction between CP and VE was observed. Hens fed diet with 16% CP, supplemented with 25 or 250mg VE/kg, had smaller egg shell and greater albumen percentage (P<0.05). Diet with 250mg VE/kg resulted in greater alpha tocopherol concentration in the yolk (P<0.05). No effects of CP or VE levels were observed on incubation performance in both studied ages (32nd and 52nd week), i.e., on incubated egg weight, on eclodibility, on relationship between egg weight and neonatal chick weight, and on subsequent performance of the progeny. However, greater neonatal chick weight (P<0.01) was observed in those from 32 week-old broiler breed hens fed diet containing 16% CP and 250mg VE/kg. It can be concluded that a 14% CP with 25mg VE/kg diet provides sufficient CP and VE to assure the productive and reproductive performance of broiler breeder hens during the laying period.
Broiler breed hen; protein; vitamin E; egg production; egg composition
Níveis de proteína e de vitamina E para matrizes de frango de corte. 1. Efeito sobre o desempenho das matrizes, composição do ovo e desempenho da progênie
(Protein and vitamin E levels for broiler breed hens. 1. Effects on broiler breed performance, egg composition and performance of their progeny)
S.L.T. Barreto1, W.M. Ferreira1* * Recebido para publicação, após modificações, em 25 de janeiro de 1999. Autor para correspondência , T.M. Gonçalves2
1Departamento de Zootecnia da Escola de Veterinária da UFMG
Caixa Postal 567
30123-970 - Belo Horizonte, MG
2Departamento de Zootecnia da UFLA
RESUMO
Avaliou-se o efeito da suplementação de dois níveis de proteína bruta (PB), 14 e 16%, e dois de vitamina E (VE), 25 e 250mg/kg, na dieta sobre o desempenho e composição do ovo de matrizes de frangos de corte, entre 24 e 54 semanas de idade e sobre o desempenho de suas progênies. Foram alojadas 480 fêmeas e 48 machos da linhagem Ross, com 18 semanas de idade, em 16 boxes com 30 fêmeas e 3 machos em cada um. As dietas foram à base de milho e farelo de soja, sendo todas isocalóricas (2750kcal de EM/kg). O delineamento experimental foi o inteiramente ao acaso, constituído de quatro tratamentos em esquema fatorial 2´2, sendo dois níveis de PB (NPB) e dois níveis de VE (NVE), representados por quatro repetições cada um. A produção de ovos/ave/dia, o número de ovos/ave-alojada, o peso do ovo, a viabilidade, o peso corporal das fêmeas no final do experimento e a percentagem de gema no ovo não foram influenciados (P>0,05) pelos tratamentos e nem pela interação entre NVE ´ NPB. Menor percentagem de casca (P<0,05) e maior percentagem de albúmen (P<0,05) foram obtidos em ovos de aves que receberam o maior NPB na dieta, independente do NVE suplementado na dieta. Maior concentração de a-tocoferol na gema foi obtida em ovos de matrizes que receberam dieta suplementada com 250mg de VE/kg (P<0,05). O rendimento de incubação em ambas as idades estudadas (32ª e 52ª semanas) resultou em efeitos não significativos (P>0,05) dos NPB e dos NVE, representados pelo peso do ovo incubado, pela taxa de eclosão, pela razão entre o peso do ovo e o peso da progênie e pelo desempenho da progênie até a idade de abate. Porém, maior peso ao nascer (P<0,01) foi encontrado na progênie proveniente de reprodutoras com 32 semanas de idade, que receberam dieta com maior NPB e maior NVE. Dieta com 14% de PB e suplementada com 25mg de VE/kg é suficiente para assegurar o desempenho produtivo e reprodutivo satisfatório de matrizes de frangos de corte durante a fase de produção.
Palavras-Chave: Matriz pesada, proteína bruta, vitamina E, produção de ovos, composição do ovo
ABSTRACT
This study was carried out with the objective of evaluating the effects of supplementation of two levels of crude protein (14 and 16% CP) and vitamin E (25 and 250mg VE/kg) in the diet on the performance and egg composition of broiler breed hens and on the performance of their progeny during the laying period (24-to-54 weeks of age). Thus 480 female and 48 male Ross breeder 18-week-old birds were randomly distributed in 16 floor pens, with 30 females and three males in each one. The diets were based on corn and soybean meal, and all them were isoenergetics (2750kcal ME/kg). The experimental design was a complete randomized block and the treatments were arranged in 2 ´ 2 factorial (CP ´ VE levels) with two repetitions each. Egg production/hen/day, egg weight, egg number/hen housed, viability, final female body weight and egg yolk percentage were not affected (P>0.05) by the treatments, and no interaction between CP and VE was observed. Hens fed diet with 16% CP, supplemented with 25 or 250mg VE/kg, had smaller egg shell and greater albumen percentage (P<0.05). Diet with 250mg VE/kg resulted in greater alpha tocopherol concentration in the yolk (P<0.05). No effects of CP or VE levels were observed on incubation performance in both studied ages (32nd and 52nd week), i.e., on incubated egg weight, on eclodibility, on relationship between egg weight and neonatal chick weight, and on subsequent performance of the progeny. However, greater neonatal chick weight (P<0.01) was observed in those from 32 week-old broiler breed hens fed diet containing 16% CP and 250mg VE/kg. It can be concluded that a 14% CP with 25mg VE/kg diet provides sufficient CP and VE to assure the productive and reproductive performance of broiler breeder hens during the laying period.
Keywords: Broiler breed hen, protein, vitamin E, egg production, egg composition
INTRODUÇÃO
Vários estudos têm sido realizados com o objetivo de verificar o efeito de diferentes níveis de proteína bruta (NPB) e de vitaminas no desempenho de poedeiras comerciais e de frangos de corte, mas poucos são os que foram desenvolvidos com matrizes de frangos de corte na fase de produção.
Entretanto, sobre algumas vitaminas, e particularmente sobre a vitamina E (VE), existem poucos trabalhos que possam ser utilizados como base para a determinação da exigência de VE para matrizes pesadas, já que dentre os encontrados na literatura, a maioria data dos anos 50 e 60 e se refere a poedeiras comerciais.
O consumo de quantidades inadequadas de VE pode provocar problemas de fertilidade, de viabilidade embrionária e até mesmo na produção. Esse fato se agrava quando as aves são submetidas ao estresse calórico, provocado pela rápida elevação na temperatura ambiente, com repercussão na redução do consumo de ração, além de elevar e acelerar perdas na concentração de VE na dieta (Scheideler & Froning, 1994).
Embora a concentração de a-tocoferol (AT) na gema do ovo possa ser facilmente elevada pelo aumento na suplementação de VE na dieta (Frigg et al., 1992; Grobas et al., 1997ab; Barreto, 1998), poucos são os trabalhos encontrados na literatura que se preocuparam em avaliar a eficiência de transferência de VE presente no ovo para o embrião e, conseqüentemente, para a progênie após o nascimento e seus efeitos sobre o desempenho zootécnico da prole.
Leeson & Summers (1997) recomendam 25mg de VE/kg de dieta para matrizes pesadas durante a fase de produção. Entretanto, o Manual... (1996) recomenda um NVE oito vezes maior para reprodutoras pesadas (200mg de VE/kg de dieta) durante a fase de produção, sendo este nível 20 vezes superior ao nível sugerido pelo Nutrient... (1994).
Por outro lado, as tabelas do Nutrient... (1994) preconizam a exigência de 19,5g de proteína/ave/dia para matrizes de frangos de corte durante todo o ciclo de produção. Controvérsias existem e entre os trabalhos publicados muitos sugerem níveis mais elevados para a melhora do desempenho produtivo e níveis mais baixos para a melhora do desempenho reprodutivo das aves.
A maioria das dietas práticas que vêm sendo utilizadas para essas aves apresenta nível de 16% de proteína bruta (PB), fornecendo até 26g de proteína/ave/dia durante o pico de produção. Segundo vários estudos, esse consumo é elevado e desnecessário para o ótimo desempenho produtivo e reprodutivo do plantel, além de elevar o custo de produção (Brum, 1994; Lopez & Leeson, 1995). Além disso, se forem comparadas as exigências nutricionais para matrizes de frangos de corte, citadas pelo Nutrient... (1994), com as recomendações contidas nos manuais da maioria das linhagens, pode-se verificar que estes sugerem níveis nutricionais superiores àqueles recomendados pelo Nutrient... (1994), provavelmente como margem de segurança em função das diversas regiões onde são criadas as reprodutoras.
Trabalhos como os de Irving & Buntz-Olson (1955) e de Hirahara et al. (1979) sugerem que a proteína e a VE apresentam um ponto comum no metabolismo, indicando que o aumento do NPB na dieta resulta em aumento da exigência de VE, havendo também necessidade de maior suplementação de VE quando se eleva o NPB na dieta.
Todavia, a respeito do nível ideal de PB e VE na dieta de matrizes de frangos de corte ainda existem controvérsias entre os pesquisadores, manuais de criação e tabelas de recomendações de exigência nutricional. E em função disso, existe uma diversidade de recomendações de VE e proteína sendo empregadas no Brasil.
Com base no que foi relatado, este trabalho teve como objetivos estudar os efeitos de diferentes NPB e da suplementação de VE na dieta sobre o desempenho produtivo e reprodutivo e sobre a composição do ovo de matrizes de frangos de corte, entre 24 e 54 semanas de idade, e o desempenho da progênie.
MATERIAL E MÉTODOS
Este experimento foi conduzido no setor de Avicultura da Fazenda Experimental Professor Hélio Barbosa, da Escola de Veterinária da UFMG. Utilizou-se um aviário convencional, constituído de 60 boxes com dimensões de 3,00 ´ 2,20m, coberto com maravalha (±10cm) sobre o piso. Cada boxe foi provido de um bebedouro pendular e de três comedouros, sendo que dois permitiam o acesso somente das fêmeas, e outro, pendurado mais alto, permitia somente o acesso dos machos. Em cada boxe experimental foram utilizados ninhos convencionais de dois andares com cinco bocas por andar, de chapa galvanizada, com dimensões de 0,30 ´ 0,30 ´ 0,30m, a 0,30m acima do nível da "cama", sendo uma boca para cada três fêmeas.
Foram utilizados 528 reprodutores de frangos de corte, sendo 480 fêmeas e 48 machos, ambos da linhagem Ross. As aves foram selecionadas na 18ª semana de idade, a partir de um plantel de reprodutores, dentro de uma faixa de peso de 1850 a 1950g para as fêmeas, e de 2450 a 2550g para os machos, obtendo-se pesos médios de 1890g e 2500g para fêmeas e machos, respectivamente. As aves foram distribuídas aleatoriamente em 16 boxes, em grupo de 30 fêmeas e 3 machos por boxe.
O período experimental teve início na 24ª semana, quando o plantel alcançou 5% de produção, e finalizou na 54ª semana de idade das aves.
O acasalamento foi realizado na 18ª semana de idade das aves, na proporção de um macho para cada 10 fêmeas. Na ocorrência de morte ou descarte do macho procedeu-se à sua substituição.
O programa de iluminação, que permaneceu o mesmo até o final do período experimental, teve início na 19ª semana de idade, com fornecimento inicial de 14 horas de luz diária, e com aumentos semanais de 30 minutos de luz até o fornecimento de 17 horas de luz diária na 25ª semana de idade das aves.
O fornecimento de ração foi limitado e controlado diariamente, e o de água foi ad libitum. A quantidade de ração foi oferecida para as fêmeas em função do nível de produção e do peso corporal e, para os machos, em função do peso corporal (Tab. 1), sendo o peso controlado semanalmente até a 35ª semana, e quinzenalmente, a partir da 36ª semana de idade das aves até o final do experimento.
Os ovos foram colhidos diariamente em bandejas de polpa de papelão descartável, três vezes ao dia, registrando-se o número de ovos de cada colheita segundo o tratamento e a repetição.
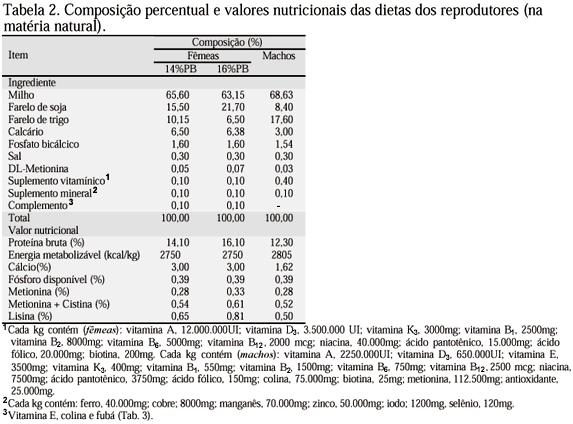
As exigências nutricionais utilizadas para a formulação das dietas das fêmeas e machos foram baseadas nas recomendações contidas no Manual... (1996). A composição percentual, química e os valores nutricionais das dietas utilizadas encontram-se na Tab. 2.
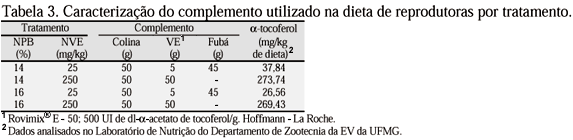
Os tratamentos foram definidos de acordo com o NPB e o NVE suplementados nas rações das fêmeas, conforme apresentado nas Tab. 2 e 3, e constaram de quatro dietas diferenciadas: 14% de PB e 25mg de VE/kg; 14% de PB e 250mg de VE/kg; 16% de PB e 25mg de VE/kg; 16% de PB e 250mg de VE/kg.
A partir da 23ª semana de idade as fêmeas passaram a receber as dietas experimentais constituídas da dieta básica (Tab. 2), suplementadas com dois NVE (Tab. 3). A dieta básica das fêmeas continha suplemento vitamínico sem VE e sem colina. Os machos passaram a receber uma única dieta, conforme mencionado na Tab. 2, durante todo o período experimental.
A produção média de ovos de cada repetição, expressa em percentagem durante todo o período experimental, foi obtida computando-se o número de ovos produzidos semanalmente, inclusive os quebrados, trincados e os anormais, e dividindo-se pelo número de fêmeas no mesmo período. O total de ovos produzidos/ave alojada de cada repetição, da 24ª a 54ª semana de idade das aves, foi calculado pela divisão entre o número de ovos produzidos e o número de aves alojadas na mesma repetição.
Todos os ovos íntegros produzidos em cada repetição foram pesados durante três dias consecutivos de cada semana, para avaliação do peso médio do ovo durante todo o período experimental.
Quinzenalmente, 10 ovos de cada repetição foram coletados aleatoriamente, sendo identificados e posteriormente incubados. Com 24 horas de incubação, todos os ovos foram quebrados para avaliação da fertilidade, mediante diferenciação do disco germinativo, computando-se o percentual de fertilidade por unidade experimental.
Na 54ª semana de idade, todas as aves de cada repetição foram pesadas em grupo para avaliação do peso médio final no período experimental.
A mortalidade das fêmeas de cada repetição foi registrada diariamente para obtenção da percentagem de sua viabilidade no final do período experimental. A viabilidade de machos não foi avaliada, havendo substituição de macho para cada ocorrência de morte.
A cada 15 dias, cinco ovos de cada repetição foram coletados aleatoriamente e identificados. No dia seguinte, os ovos foram pesados em grupo por repetição e, posteriormente, no mesmo dia, quebrados. As cascas foram lavadas e secadas em temperatura ambiente e pesadas no dia seguinte. O peso médio do albúmen de cada repetição foi obtido pela diferença entre o peso médio do ovo e o peso médio da gema somado ao peso médio da casca de cada repetição. Esses dados foram transformados em percentagens e analisados.
Para a avaliação do conteúdo de a-tocoferol (AT) na gema do ovo de reprodutoras, dois ovos de cada repetição foram coletados aleatoriamente na 30ª; 40ª e 50ª semanas de idade das aves. O conteúdo de AT na gema foi obtido em triplicatas e estimado em mg de AT/g de amostra, segundo a metodologia proposta por Ueda & Igarashi (1990).
Para a avaliação do rendimento de incubação foram realizadas duas incubações, uma na 32ª semana de idade das aves e outra na 52ª semana. Para cada incubação foram coletados 60 ovos de cada repetição durante quatro dias consecutivos nas duas idades mencionadas. Um total de 960 ovos foram pesados e incubados em cada idade avaliada, de acordo com o tratamento e repetição, e avaliadas as seguintes características: peso médio do ovo incubado, peso médio dos pintos ao nascer, percentual do peso da progênie ao nascer em relação ao peso do ovo, taxa de eclosão e desempenho da progênie até ao 42º dia de idade.
O delineamento experimental foi o inteiramente ao acaso, em esquema fatorial 2 ´ 2 (NPB ´ NVE) com 4 repetições de 30 fêmeas e 3 machos por unidade experimental. Para a análise de variância, utilizou-se o programa estatístico (SAS..., 1985). Quando as interações entre dois fatores tiveram efeitos significativos foram feitos os desdobramentos das interações estudando-se os efeitos de NVE dentro de NPB na dieta, mediante o teste F.
RESULTADOS E DISCUSSÃO
Pelos resultados obtidos, apresentados na Tab. 4, observa-se que não houve interação significativa (P>0,05) entre os NPB e os NVE, assim como também não houve diferenças significativas (P>0,05) na produção de ovos/ave/dia, no número de ovos acumulados por ave alojada, no peso do ovo, na viabilidade e no peso corporal de reprodutoras em função do NPB e do NVE na dieta entre 24 e 54 semanas de idade. No entanto, verificou-se maior fertilidade nos ovos de matrizes que receberam dieta com 25mg de VE/kg.
Trabalhos conduzidos com matrizes mostram que o NPB da dieta exerce pouca ou nenhuma influência sobre o desempenho produtivo das aves, desde que haja suplementação de aminoácidos essenciais quando se reduz a proteína da dieta (Brum, 1994; Lopez & Leeson, 1995).
A redução do NPB de 16 para 14% não exerceu nenhum efeito (P>0,05) sobre o desempenho das aves, visto que a dieta formulada para 14% de PB continha 0,54% de metionina + cistina, sendo capaz de atender à exigência desses aminoácidos segundo o Nutrient... (1994). Contudo, o nível de proteína da dieta de matrizes de frangos de corte na fase de produção pode ser reduzida de 16 para 14% sem afetar o desempenho do plantel, desde que as exigências dos aminoácidos essenciais sejam supridas. A dieta com 14% de PB proporcionou um consumo médio diário de 23g de proteína e de 891mg de metionina + cistina /ave na fase de pico de produção e de 21g de proteína e de 810mg de metionina + cistina /ave na fase final de produção.
Com relação aos resultados de desempenho obtidos em função dos NVE suplementados na dieta de reprodutoras (Tab. 4), verifica-se que a suplementação de 10 vezes a mais de VE na dieta de reprodutoras não trouxe nenhum efeito para a melhora do desempenho, conforme foi sugerido pelo Manual... (1996). Tais achados concordam com os encontrados por Jiang et al. (1994) e Grobas et al. (1997a) que também não encontraram efeito do NVE da dieta sobre as características de desempenho. No entanto, discordam com Scheideler & Froning (1994) que encontraram melhora de 2% na produção de ovos de poedeiras alimentadas com dieta com 50mg de VE/kg.
Leeson & Summers (1997) sugerem a exigência de 25, 20 e 10mg de VE/kg de dieta para reprodutoras pesadas, reprodutoras leves e poedeiras leves, respectivamente. Por outro lado, Waldroup (1997) traz, como exigência, 10mg de VE/kg de dieta para reprodutoras e 5mg de VE/kg para poedeiras, enquanto que o Manual... (1996) menciona níveis de 200mg de VE/kg de dieta, sendo este nível 20 vezes superior ao nível sugerido pelo Nutrient... (1994).
Acredita-se que a VE traz melhora ao desempenho de matrizes apenas quando for suplementada a dieta que contenha níveis inferiores ao sugerido pelo Nutrient... (1994), ou seja, menos que 10mg de VE/kg de dieta.
De acordo com os dados da Tab. 5, observa-se que não houve interação significativa (P>0,05) do NPB x NVE para as percentagens de gema, de albúmen e de casca, como também não houve efeito do NVE (P>0,05) sobre essas variáveis. Entretanto, verificou-se que as percentagens de albúmen (P<0,05) e de casca (P<0,01) foram significativamente influenciadas pelos NPB, porém o mesmo não foi observado para a percentagem de gema.
Os resultados mostram que quando as aves foram alimentadas com dieta com 16% de PB foi obtido 0,9% a mais na percentagem de albúmen. No entanto, maior percentagem de casca (2,2% a mais) foi notada em ovos de reprodutoras que receberam dieta com o menor NPB.
Embora se saiba da importância da proteína e da VE na dieta das aves poucos são os trabalhos realizados com reprodutoras pesadas, e dentre os existentes, raros são os que avaliaram a composição do ovo. Entre estes, existe grande controvérsia quanto a essa característica.
Com relação à concentração de AT na gema do ovo, observou-se que não houve interação significativa (P>0,05) do NVE ´ NPB, como também não houve efeito (P>0,05) do NPB sobre a deposição de AT na gema. Entretanto, houve aumento significativo (P<0,01) na concentração de AT na gema com a utilização de 250mg de VE/kg de dieta (Tab. 5).
Segundo Rosebrough et al. (1982), o aumento no nível de PB na dieta resulta em redução na lipogênese e, dessa forma, esperava-se que poderia haver menor deposição de vitamina na gema de aves que receberam dieta com 16% de PB.
Trabalhando com poedeiras comerciais alimentadas com dieta de 250mg, em média, de VE/kg, Frigg et al. (1992) também encontraram deposição de AT que variou de 400 a 600mg/g de gema, dependendo da idade das aves e do tipo de óleo utilizado na dieta. Ao contrário, Jiang et al. (1994) verificaram deposição de 134,8 e de 245,8mg de AT/g de gema em ovos de aves alimentadas com dieta de 27 e 200mg de VE/kg, respectivamente.
Ao suplementar a dieta das aves deste experimento com 25 ou 250mg de VE/kg, verificou-se deposição média de 117,9 e 702,7mg de AT/g de gema, respectivamente, ou seja, um aumento de 6 vezes. No entanto, verifica-se que as concentrações de AT na gema foram superiores àquelas encontradas por Jiang et al. (1994), quando utilizaram NVE bem próximos ao deste experimento.
Com relação aos resultados do rendimento de incubação dos ovos de matrizes de 32 semanas (Tab. 6), observa-se que o peso do ovo incubado, o percentual do peso da progênie ao nascer em relação ao peso do ovo e a taxa de eclosão não foram influenciados (P>0,05) pela interação entre o NPB e o NVE, como também pelo NPB e pelo NVE utilizados na dieta (P>0,05). Porém, houve interação significativa entre o NPB e o NVE (P<0,05) para o peso da progênie ao nascer. O resultado dessa interação mostrou que a dieta com o maior NPB (16%) proporcionou 3,5% a mais no peso da progênie ao nascer, quando esta dieta foi suplementada com o maior NVE (250mg/kg).
O Manual... (1996) menciona eclodibilidade padrão de 88% em ovos de reprodutoras Ross com 32 semanas de idade. Com base nos resultados obtidos neste experimento, observou-se que eclodibilidade maior do que 88,0% só não foi obtida em ovos de reprodutoras que consumiram dieta com 16% de PB e 250mg de VE/kg.
Embora se saiba que a VE é essencial para a manutenção de níveis ótimos da fertilidade e da eclodibilidade e que o aumento da suplementação dessa vitamina na dieta resulta em elevação na concentração de AT nos tecidos (Maurice et al. 1996) e na gema do ovo (Maurice et al., 1996; Jiang et al., 1994; Grobas et al., 1997a,b), trabalhos publicados mostram que, quando a VE é suplementada na dieta em níveis mais elevados do que o nível sugerido pelo Nutrient... (1994), não se observa nenhuma melhora na fertilidade e na eclodibilidade dos ovos (Arnold et al., 1974; Bartov et al., 1965).
Analisando os valores médios referentes ao rendimento de incubação de ovos de matrizes de 52 semanas de idade (Tab. 7), verifica-se que o peso do ovo incubado, o peso da progênie ao nascer, o percentual do peso da progênie em relação ao peso do ovo, e a taxa de eclosão dos ovos não foram influenciados (P>0,05) pelos NPB e pelos NVE utilizados na dieta das aves. Também não foi observada interação significativa (P>0,05) entre os NPB e os NVE.
Embora fosse observado menor percentagem de casca (P<0,05) em ovos de matrizes que receberam dieta com 16% de PB (Tab. 5), verifica-se que esta ocorrência não afetou significativamente (P>0,05) a taxa de eclosão dos ovos de matrizes no pico e na fase final de produção (Tab. 6 e 7).
Sabe-se que com o avanço na idade das aves ocorrem mudanças no comportamento fisiológico, evidenciando mudanças na atividade hormonal e, conseqüentemente, resultando em aumento no tamanho do ovo, maior peso da progênie ao nascer e maior razão entre o peso da progênie e o peso do ovo. Porém, observa-se queda na fertilidade e na eclodibilidade. Dessa forma, esperava-se que ao suplementar a dieta de reprodutoras de 52 semanas de idade com níveis de 250mg de VE/kg se pudesse obter melhora na fertilidade e na taxa de eclosão, conforme o Manual... (1996), que sugere a suplementação de 200mg de VE/kg de dieta.
Segundo Pearson & Herron (1982), baixa eclodibilidade tem sido associada com a utilização de elevado nível protéico na dieta, principalmente quando o consumo de energia for baixo. Porém, esse efeito não foi observado quando o consumo de energia foi elevado (470kcal de EM/ave/dia), ou quando as aves consumiram 21,3g de PB/ave/dia.
Enquanto deficiências vitamínicas só deveriam ocorrer sob circunstâncias incomuns, existe indicação de eclodibilidade mais baixa relacionada ao metabolismo das vitaminas quando são fornecidas dietas com elevado nível protéico, o que sugere problemas com a adequação de vitaminas para o ovo (Leeson, 1995). No entanto, pelos resultados obtidos, a VE parece não se enquadrar nesse grupo de vitaminas, visto que não foi observada interação significativa entre os NVE e os NPB utilizados neste trabalho.
Os NVE e os NPB utilizados na dieta de matrizes de 32 e 52 semanas de idade não exerceram efeito significativo (P>0,05) no consumo de ração, no peso corporal aos 42 dias, no ganho de peso do 1º ao 42º dia de idade e na conversão alimentar da progênie (Barreto, 1998).
Estudos mostram que um grama a mais no peso do pinto ao nascer resulta em 10 a 12g a mais no peso corporal na idade de abate (Al-Murrani, 1978). Entretanto, embora neste trabalho tenha havido diferença significativa de 2,08g a mais no peso ao nascer da progênie de reprodutoras com 32 semanas de idade que receberam 16% de PB (Tab. 6), não se verificou diferença significativa no peso aos 42 dias dessa progênie (Barreto, 1998).
Os resultados do presente experimento reforçam a idéia de Brum (1994) e de Lopez & Leeson (1994) que afirmaram não terem encontrado diferença significativa no desempenho da progênie proveniente de matrizes com mais de 45 semanas de idade e alimentadas com dieta com diferentes NPB.
Embora houvesse aumento significativo na concentração de AT na gema do ovo de reprodutoras que receberam dieta com o nível mais elevado de VE (Tab. 5), observou-se que esse efeito não exerceu nenhuma influência na fertilidade, na eclodibilidade e no peso da progênie ao nascer (Tab. 6 e 7), e no subseqüente desempenho da progênie proveniente de reprodutoras no pico e na fase final de produção (Barreto, 1998). Com isso, pode-se concluir que dentre os níveis avaliados, 25mg de VE/kg de dieta e dieta com 14% de PB foram suficientemente adequados na alimentação de reprodutoras com base no desempenho da progênie.
Os valores médios de AT na carne de peito da progênie aos 45 dias de idade também não foram influenciados (P>0,05) pelos NVE ou pelos NPB contidos na dieta das progenitoras, como também não houve influência do sexo da progênie sobre a concentração (Barreto, 1998). Desse modo, a suplementação de VE em níveis mais elevados na dieta de reprodutoras, durante a fase de produção, não mantém a concentração elevada de AT na carne de peito da progênie no momento do abate.
Bartov et al. (1965) demonstraram que a concentração de AT na gema do ovo, no saco vitelínico e no fígado de pintos de um dia foram influenciados pelos NVE na dieta das progenitoras. No entanto, experimentos com perus mostraram que a reserva de tocoferol decresce rapidamente durante as primeiras semanas de vida. Essa queda ocorre independentemente do aumento do NVE ou de antioxidante na dieta, pois as aves apresentam baixa capacidade de armazenamento prolongado de AT nos tecidos (Soto-Solanova et al., 1993). Dessa forma, a suplementação de níveis elevados de VE na dieta de reprodutoras não contribui para melhora do desempenho da progênie, assim como não mantém aumento na concentração de AT na carcaça com o objetivo de prolongar o seu armazenamento após o abate.
CONCLUSÕES
Com base nestes resultados pode-se concluir que: 1 - o fornecimento de dieta com 14% de PB e 25mg de vitamina E/kg supre a exigência desses nutrientes para matrizes de frangos de corte quanto ao desempenho produtivo e reprodutivo no período de 24 a 54 semanas de idade; 2 - o aumento de 25 para 250mg de vitamina E/kg de dieta eleva em seis vezes a concentração de a-tocoferol na gema do ovo, independentemente do nível de PB da dieta; 3 - não é viável suplementar a dieta de matrizes de frangos de corte com níveis superiores a 25mg de vitamina E/kg de dieta e 14% de PB, tanto na fase de pico como na fase final de produção, visto que não ocorrem alterações na concentração de a-tocoferol na carne de peito e no desempenho da progênie até a idade de abate.
- AL-MURRANI, K.W. Maternal effects on embrionics and post embryonic growth in poultry. Br. Poult. Sci., v.19, p.277-281, 1978.
- ARNOLD, R.L., OLSON, O.E., CARLSON, C.W. Tissue selenium content and serum tocopherol is influenced by dietary type, selenium and vitamin E. Poult. Sci., v.53, p.2185-2192, 1974.
- BARRETO, S.L.T. Níveis de proteína e vitamina E para matrizes de frangos de corte na fase de produçăo Belo Horizonte: Escola de Veterinária da UFMG, 1998. 173p. (Tese, Doutorado).
- BARTOV, I., BUDOWSKI, P., BORNSTEIN, S. The relation between a-tocopherol content of the breeder diet and that of the newly hatched chick. Poult. Sci., v.44, p.1489-1494, 1965.
- BRUM, P.A.R. Exigęncia de proteína em dietas para reprodutoras de frangos de corte durante a fase de produçăo. Porto Alegre: UFRGS, 1994. 189p. (Tese, Doutorado).
- FRIGG, M., WHITEHEAD, C.C., WEBER, S. Absence of effects of dietary a-tocopherol on egg yolk pigmentation. Br. Poult. Sci., v. 33, p. 347-353, 1992.
- GROBAS, S., MÉNDEZ, J., BLAS, C. et al. Influence of dietary vitamins E and A on performance, egg quality and a-tocopherol content of yolks. Poult. Sci., v.76, p.93, 1997a. (Supplement, 1).
- GROBAS, S., MÉNDEZ, J., BLAS, C. et al. Influence of level of vitamin E and type of dietary fat on concentration of a-tocopherol, oxidative status, and fatty acid profile of egg yolks. Poult. Sci., v.76, p.93, 1997b. (Supplement, 1)
- HIRAHARA, F., TAKAI, Y., IWAO, H. The effects of vitamin E on protein metabolism in the aging process of rats. Nutr. Rep. Int., v.20, p.261-267, 1979.
- IRVING, J.T., BUNTZ-OLSEN, O.E. The relation between the action of vitamin E and protein in the body, and the influence of various fish-liver oils in the diet upon vitamin E activity in vivo. Br. J. Nutr. v.9, p.301-309, 1955.
- JIANG, Y.H., McGEACHIN, R.B., BAILEY, C.A. a-tocopherol, b-carotene, and retinol enrichment of chicken eggs. Poult. Sci., v.73, p.1137-1143, 1994.
- LEESON, S. Nutriçăo e eclodibilidade em reprodutoras de corte. In: CONFERĘNCIA APINCO 1995 DE CIĘNCIA E TECNOLOGIA AVÍCOLAS, 1995, Campinas. Trabalhos de pesquisa... Campinas: Facta, 1995. p.51-60.
- LEESON, S., SUMMERS, J.D. Commercial poultry nutrition. Guelph: University Books, 1997. 350p.
- LOPEZ, G., LEESON, S. Response of older broiler breeders to medium-high intakes of protein. J. Appl. Poult. Res., v.3, p.157-163, 1994.
- LOPEZ, G., LEESON, S. Response of broiler breeders to low-protein diets. 1. Adult breeder performance. Poult. Sci., v.74, p.685-695, 1995.
- MANUAL de manejo de matrizes AG ROSS. Rio Claro: Agroceres Ross Melhoramento Genético de Aves, 1996. 87p.
- MAURICE, DV., ELZUBIER, E.A., LIGHSEY, S.F. Breeder diet vitamin E concentration and post-hatch changes in tissue vitamin E status of chickes. Poult. Sci., v. 75, p.128, 1996. (Supplement, 1).
- NUTRIENT requirements of poultry. 9.ed., Washington: National Academy Press, 1994. 155p.
- PEARSON, R.A., HERRON, K.M. Effects of maternal energy and protein intakes on the incidence of malformations and malpositions of the embryo and time of death during incubation. Br. Poult. Sci., v. 23, p. 71-77, 1982.
- ROSEBROUGH, R.W., STEELE, N.C., FROBISH, L.T. Studies on the role of lysine and protein in the regulation of lipogenesis by the turkey poult. Poult. Sci., v.61, p.2056-2059, 1982.
- SAS. User’s guide: statistics, Cary: SAS Institute, 1985. 956p.
- SCHEIDELER, S.E., FRONING, G. Dietary flaxseed improves egg production and incorporation of omega 3 fatty acids in eggs. Poult. Sci., v.73, p.50, 1994. (Supplement 1)
- SOTO-SALANOVA, M.F., SELL, J.L. Efficacy of dietary and injected vitamin E for poults. Poult. Sci., v.73, p.14, 1994. (Supplement, 1)
- UEDA, T., IGARASHI, O. Determination of vitamin E biological specimens and foods by HPLC-Pretreatment of samples and extraction of tocopherols. J. Micronutr. Anal., v.7, p.79-96, 1990.
Datas de Publicação
-
Publicação nesta coleção
17 Abr 2001 -
Data do Fascículo
Abr 1999
Histórico
-
Recebido
25 Jan 1999