Resumos
Amostras do vírus da artrite-encefalite caprina (CAEV) de animais dos estados de Minas Gerais e Pernambuco foram isoladas a partir de explantes de membrana sinovial (MS) (amostras BrMg 1-01, BrMg 2-01, BrMg 2-02, BrMg 2-03 e BrPe 1-01) ou de co-cultivo de leucócitos com MS (BrMg 1-02). As amostras foram identificadas pelo efeito citopático-ECP (formação de sincícios), imunofluorescência direta (IFD) e reação em cadeia de polimerase (PCR) aperfeiçoada para amplificação de parte do gene gag da amostra CAEV Cork. O estudo do ECP revelou a presença de amostras pouco (BrMg 1-01 e BrMg 1-02) ou fortemente indutoras de ECP (BrMg 2-01, BrMg 2-02, BrMg 2-03 e BrPe 1-01). A IFD mostrou resultado positivo em células de todas as monocamadas infectadas pelas amostras isoladas e em células de referência (CAEV Cork e visna/maedi K 1514). A PCR do DNA de células infectadas resultou na amplificação específica de um fragmento de DNA de 286pb da amostras testadas, exceto do visna/maedi K 1514.
Caprino; artrite-encefalite; reação em cadeia de polimerase
This paper describes the isolation of caprine arthritis-encephalitis virus (CAEV) in Minas Gerais and Pernambuco States, Brazil, using goat synovial membrane (GSM) explants (isolates: BrMg 1-01, BrMg 2-01, BrMg 2-02, BrMg 2-03 and BrPe 1-01) or co-culture of blood mononuclear cells with GSM (BrMg 1-02). The isolates were identified by cytopathic effect (CPE), direct immunofluorescence (DIF) and an improved polimerase chain reaction (PRC) specific to CAEV (strain Cork) genome. It was found isolates of low (BrMg 1-01 and BrMg 1-02) and of high lytic (BrMg 2-01, BrMg 2-02, BrMg 2-03 and BrPe 1-01) phenotypes. The DIF revealed positive results in cells infected by all isolates tested and the reference strains of CAEV (Cork) and visna-maedi (K1514). The used PCR amplified a 286 pb fragment of DNA from cells infected with all isolates and CAEV Cork but not with visna-maedi K1514.
Caprine; arthritis encephalitis; polimerase chain reaction
Isolamento e identificação pela imunofluorescência direta e reação em cadeia de polimerase do vírus da artrite-encefalite caprina
(Caprine arthritis encephalitis virus isolation and identification using fluorescent antibody and polimerase chain reaction)
R.S. Castro1, R.C. Leite2, M. Resende3, A. Martins3, A.M.G. Gouveia2
1Departamento de Medicina Veterinária, Universidade Federal Rural de Pernambuco
Rua D. Manoel de Medeiros s/n, Dois Irmãos
52171-900 - Recife, PE.
2Escola de Veterináriada Universidade Federal de Minas Gerais
3Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais
Recebido para publicação em 8 de abril de 1998.
Colaboradores: B.P. Drumond, S.A. Nascimento
RESUMO
Amostras do vírus da artrite-encefalite caprina (CAEV) de animais dos estados de Minas Gerais e Pernambuco foram isoladas a partir de explantes de membrana sinovial (MS) (amostras BrMg 1-01, BrMg 2-01, BrMg 2-02, BrMg 2-03 e BrPe 1-01) ou de co-cultivo de leucócitos com MS (BrMg 1-02). As amostras foram identificadas pelo efeito citopático-ECP (formação de sincícios), imunofluorescência direta (IFD) e reação em cadeia de polimerase (PCR) aperfeiçoada para amplificação de parte do gene gag da amostra CAEV Cork. O estudo do ECP revelou a presença de amostras pouco (BrMg 1-01 e BrMg 1-02) ou fortemente indutoras de ECP (BrMg 2-01, BrMg 2-02, BrMg 2-03 e BrPe 1-01). A IFD mostrou resultado positivo em células de todas as monocamadas infectadas pelas amostras isoladas e em células de referência (CAEV Cork e visna/maedi K 1514). A PCR do DNA de células infectadas resultou na amplificação específica de um fragmento de DNA de 286pb da amostras testadas, exceto do visna/maedi K 1514.
Palavras-Chave: Caprino, artrite-encefalite, reação em cadeia de polimerase
ABSTRACT
This paper describes the isolation of caprine arthritis-encephalitis virus (CAEV) in Minas Gerais and Pernambuco States, Brazil, using goat synovial membrane (GSM) explants (isolates: BrMg 1-01, BrMg 2-01, BrMg 2-02, BrMg 2-03 and BrPe 1-01) or co-culture of blood mononuclear cells with GSM (BrMg 1-02). The isolates were identified by cytopathic effect (CPE), direct immunofluorescence (DIF) and an improved polimerase chain reaction (PRC) specific to CAEV (strain Cork) genome. It was found isolates of low (BrMg 1-01 and BrMg 1-02) and of high lytic (BrMg 2-01, BrMg 2-02, BrMg 2-03 and BrPe 1-01) phenotypes. The DIF revealed positive results in cells infected by all isolates tested and the reference strains of CAEV (Cork) and visna-maedi (K1514). The used PCR amplified a 286 pb fragment of DNA from cells infected with all isolates and CAEV Cork but not with visna-maedi K1514.
Keywords: Caprine, arthritis encephalitis, polimerase chain reaction
INTRODUÇÃO
Lentivírus é um grupo de vírus da família Retroviridae responsável por enfermidades de grande importância médica e veterinária, de curso geralmente longo e irreversível. Nesse grupo estão classificados os vírus da imunodeficiência humana (HIV), símia (SIV), felina (FIV) e bovina (BIV), anemia infecciosa eqüina (EIA), visna/maedi do ovino (V/M) e artrite-encefalite caprina (CAEV) (Clements & Payne, 1994).
As primeiras suspeitas etiológicas de lentivírus em caprinos foram descritas por Weinhold et al. (1974), que observaram, pela microscopia eletrônica, partículas virais semelhantes em forma e tamanho às do vírus visna/maedi, em células de cultura de plexo coróide de uma cabra afetada por sintomatologia nervosa. Essa suspeita foi confirmada pelo isolamento de um vírus sincicial do plexo coróide e líquido cérebro-espinhal de uma cabra clinicamente afetada por paresia e paralisia progressiva (Weinhold & Triemer, 1978), do explante do timo de um animal inoculado experimentalmente com suspensão cerebral de um caso natural de encefalite caprina (Narayan et al., 1980) e de membrana sinovial de articulações de animais artríticos, passando o agente a ser denominado CAEV (Crawford et al., 1980).
Depois do isolamento do vírus, estudos sorológicos têm sido conduzidos em diversos países demonstrando a ampla distribuição geográfica da doença (Adams et al., 1984), assim como tentativas bem-sucedidas de isolamento do vírus, inclusive no Brasil (Hotzell et al., 1993). Com os avanços nas técnicas de biologia molecular, têm sido possível o seqüenciamento e o aprofundamento dos conhecimentos sobre o genoma do vírus, que apresenta a estrutura básica dos lentivírus, com genes codantes para proteínas estruturais (gag e env) e os de controle da replicação e expressão viral (pol, tat, rev e vif) (revisão de Clements & Payne, 1994). Isso tem tornado possível lançar mão de recursos modernos como a reação em cadeia de polimerase (PCR) como instrumento potencial na identificação de amostras isoladas e no diagnóstico da doença (Rimstad et al., 1993; Leroux et al., 1997).
Este trabalho relata o isolamento de amostras do CAEV de caprinos dos estados de Minas Gerais e Pernambuco e sua caracterização preliminar pelo comportamento em cultivo celular, imunofluorescência direta (IFD) e PCR.
MATERIAL E MÉTODOS
Foram selecionadas para isolamento de amostras do CAEV cabras soropositivas a um teste imunoenzimático (Lab-ELISA) para pesquisa de anticorpos séricos anti-CAEV (Castro et al., 1999), pertencentes a rebanhos comerciais, monitorados em programas de controle da doença nos estados de Minas Gerais e de Pernambuco (Tab. 1). Ao contrário dos demais, um dos rebanhos não apresentava histórico de alterações típicas de CAEV. Os animais foram examinados clinicamente, com ênfase nos sistemas comumente afetados pela CAEV. Dos necropsiados foram colhidos espécimens de membrana sinovial (MS), pulmões e glândula mamária, fixados em formalina tamponada a 10%, para exame histopatológico, realizado após coloração pela técnica de H&E, como colhidas foram também as articulações carpo-metacarpianas para cultivo in vitro de MS. Dos animais não sacrificados foram colhidas amostras de sangue em tubos com heparina1 1 Vacutainer; Becton Dickinson, USA. para obtenção de leucócitos para co-cultivo com células de MS.
As células para multiplicação das amostras foram obtidas de explantes de MS de cabritos soronegativos, filhos de cabras também soronegativas ao lab-ELISA para CAEV, de acordo com Abreu et al. (1998). O isolamento das amostras virais dos animais necropsiados foi feito também por explantação de MS, enquanto que nos não sacrificados foi feito por co-cultivo de leucócitos com MS. Os leucócitos foram obtidos por centrifugação de 5,0ml do sangue heparinizado, em gradiente de ficol2 1 Vacutainer; Becton Dickinson, USA. e tratamento com 5,0ml cloreto de amônia a 0,84%. Após lavagens em PBS, foram inoculadas em garrafas de 25cm2 de área de cultivo celular e placas de 24 poços. Após uma hora de incubação a 37ºC e 5% de CO2, as garrafas e placas foram lavadas para remoção das células que não aderiram à superfície de cultivo e acrescentadas as células de MS, passadas na diluição 1:3. Finalmente, as culturas foram incubadas e examinadas para verificação do desenvolvimento celular e surgimento de fusão, com formação de sincícios, que eram confirmados por coloração das placas com cristal violeta a 0,1%. Quando necessário as culturas foram passadas como de rotina, sempre preparando-se garrafas e placas.
As amostras isoladas e as padrão do vírus CAEV3 1 Vacutainer; Becton Dickinson, USA. (amostra Cork) e do vírus visna/maedi4 1 Vacutainer; Becton Dickinson, USA. (K1514) foram multiplicadas em células de MS. Antes da inoculação, os estoques virais foram titulados em microplacas por diluições decimais e o título, calculado pelo método de Reed & Muench (1938), definido como a recíproca da diluição que apresentou, 14 dias após inoculação, sincícios em 50% dos poços inoculados, e expresso como uma dose formadora de sincícios (DFS).
A identificação das amostras foi feita pela IFD e PCR, realizadas em células do próprio explante ou após inoculação em MS. Como controles foram usadas células de MS não infectadas e células infectadas com amostra-padrão dos vírus CAEV e visna/maedi. A IFD foi efetuada utilizando-se conjugado preparado a partir de imunoglobulinas de soro ovino anti-visna/maedi, purificadas pela precipitação com ácido caprílico e sulfato de amônia (McKinney & Parkinson, 1987) e marcadas por diálise contra solução a 0,1mg de isotiocianato de fluoresceína/ml de tampão carbonato 0,05M em salina (Clark & Shepard, 1963). A reação foi feita em lâminas, onde distribuíam-se, por poço, 25µl da suspensão celular (2´106/ml) obtida por tripsinização da monocamada infectada. Após secagem e fixação em acetona a - 20ºC, durante 10 minutos, processou-se a reação com o conjugado diluído 1:3 (Goldman, 1968).
As amostras de DNA para PCR foram preparadas a partir de suspensões celulares obtidas por tripsinização das monocamadas ou dos fragmentos dos explantes destacados das culturas. Após uma centrifugação (1000rpm/5 minutos; Centrifugador Exelsa-25 1 Vacutainer; Becton Dickinson, USA. ; rotor 8´15ml) e duas lavagens com 10ml de PBS, as células foram ressuspensas em tampão hipertônico (0,32M sacarose; 10mM Tris.HCl, pH 7,5; 5,0mM MgCl2; 1,0% Triton X 100), para lise do citoplasma, durante alguns minutos na temperatura ambiente. Em seguida, o material foi centrifugado (2500rpm/10 minutos; Centrifugador Exelsa-2, rotor 8´15ml), o sedimento lavado com PBS e ressuspenso em 250µl de tampão PCR (50mM KCl; 1,0mM MgCl2; 10mM Tris.HCl, pH 8,0; 5% glicerol; 0,05% Tween 20) e tratado com 100µg de proteinase K/ml, durante 60 minutos a 56ºC. Finalmente, a proteinase K foi inativada termicamente (cerca de 100ºC durante 10 minutos) e as amostras acondicionadas em geladeira e, posteriormente, em freezer a 20ºC.
A PCR utilizada foi a descrita por Reddy et al. (1993) para amplificação de um fragmento de 286pb do gene gag da amostra CAEV Cork com aperfeiçoamentos nas condições da reação. Foram avaliadas cinco possibilidades de combinações de ciclos (Minicycler)6 1 Vacutainer; Becton Dickinson, USA. , permanecendo invariáveis os ciclos iniciais (1 ciclo de: 94ºC - 5 min; 5 ciclos de: 94ºC - 1 min, 52ºC - 1 min, 72ºC - 1 min) e o final (1 ciclo de 72ºC - 5min e 4ºC - até a colheita das amostras). Nos 25 ciclos intermediários (inicialmente de 92ºC - 1 min, 52ºC - 1 min e 72ºC - 1 min) foram testadas variações na temperatura de desnaturação (90ºC e 87ºC) e, mantendo-se a temperatura de desnaturação de 87ºC, na de anelamento (52ºC, 56ºC e 60ºC). Em paralelo foram feitas reações nas condições descritas por Reddy et al. (1993).
Em todos os casos, a reação foi conduzida em volume final de 100µl, constando de 0,25 a 0,5µg de DNA (estimado com auxílio do GeneQuant)7 1 Vacutainer; Becton Dickinson, USA. 1,0 UI de Taq DNA polimerase8 8 CENBIOT/RS, Brasil. , 50pmol de cada oligonucleotídeo9 8 CENBIOT/RS, Brasil. , 200µM de cada dNTP10 8 CENBIOT/RS, Brasil. (dATP, dCTP, dGTP e dTTP), 10mM Tris.HCl (pH 8,0), 50mM KCl, 1,5mM MgCl2, 0,01% (W/V) gelatina, coberto com 80µl de óleo mineral. O produto da reação foi submetido a eletroforese em agarose a 1,5% e, após coloração com brometo de etídio (10mg/ml), examinado comparativamente com marcadores de peso molecular11 11 BRL Life Techonogies, USA. (F´174 e 100pb Ladder), com auxílio de transiluminador12 11 BRL Life Techonogies, USA. O resultado foi registrado por fotografia.13 11 BRL Life Techonogies, USA.
RESULTADOS
Os animais estudados apresentavam aspecto aparentemente normal (animal BrMg 1-02) ou parcialmente debilitado sem alterações articulares (BrPe 1-01) ou com aumento das articulações (BrMg 1-01, BrMg 2-01, BrMg 2-02, BrMg 2-03) (Tab. 1). Ao exame pós-morte foram observadas lesões de artrite e periartrite, com aumento das articulações, espessamento da cápsula articular e conteúdo abundante, variando de seroso a sanguinolento, de aspecto achocolatado. O animal BrPe 1-01 apresentou áreas discretamente marmóreas na região dos lobos diafragmáticos dos pulmões que, ao corte, se aprofundavam pelo parênquima. Ao exame microscópico observou-se artrite crônica típica de CAEV (BrMg 1-01, BrMg 2-01, BrMg 2-02, BrMg 2-03) e discreta pneumonia intersticial (BrPe 1-01), caracterizada por infiltração linfocítica difusa.
O estudo do ECP revelou a presença de amostras pouco (BrMg 1-01 e BrMg 1-02 ) ou fortemente indutoras de ECP (BrMg 2-01, BrMg 2-02, BrMg 2-03 e BrPe 1-01) com raros e pequenos ou numerosos e grandes sincícios, respectivamente (Fig. 1; Tab. 2). A imunofluorescência direta mostrou coloração de parte do citoplasma celular nas células de todas as monocamadas e das infectadas com CAEV Cork e visna/maedi K 1514 (Fig. 2; Tab. 2).
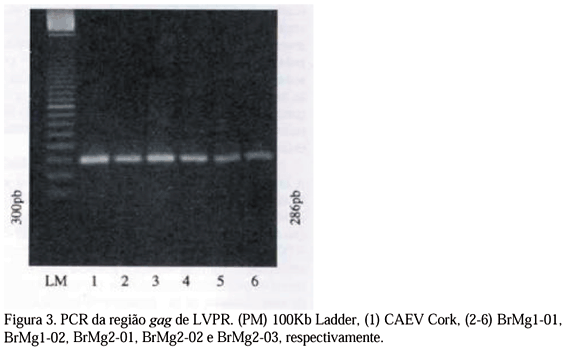
Durante a padronização da PCR evidenciou-se que a diminuição, nos ciclos intermediários, da temperatura de desnaturação de 92ºC para 90ºC e 87ºC resultou na obtenção de bandas mais evidentes, não sendo perceptíveis diferenças entre as temperaturas de 90ºC e 87ºC. No que diz respeito à temperatura de anelamento, não foi observada diferença nas temperaturas de 52ºC, 56ºC e 60ºC. Dessa forma, foram escolhidas as temperaturas de 87ºC e 60ºC como, dentre as testadas, ideais para desnaturação e anelamento, respectivamente. A PCR padronizada, nessas condições, resultou em produtos com bandas mais fortes que as observadas com os ciclos descritos por Reddy et al. (1993). As PCR das amostras testadas e da CAEV Cork produziram fragmentos do tamanho esperado (286 pb), enquanto que não foi observada amplificação do DNA obtido de células de MS não infectadas ou infectadas com visna/maedi K1514 (Fig. 3; Tab. 2).
DISCUSSÃO
Os achados nos rebanhos bem como as alterações registradas nos animais são, em linhas gerais, típicos das observações feitas no Brasil com estudos epidemiológicos e anátomo-histopa-tológicos mais detalhados, onde tem-se observado alta prevalência de animais soropositivos em rebanhos de raças européias (Saraiva Neto et al., 1995) e lesões típicas de CAEV (Pereira, 1995). Entretanto, o que chamou a atenção foi a ausência de histórico clínico típico de CAEV no rebanho três, apesar da sua soropositividade. Isso pode indicar a presença de uma amostra pouco indutora de lesões de CAEV ou, alternativamente, o uso de práticas de manejo adequadas que minimizaram a incidência de alterações clinicamente detectáveis. Além disso, merece atenção o isolamento de amostras fortemente indutoras de ECP, ao contrário do que mostraram os estudos com a amostra padrão CAEV Cork (Querat et al., 1984).
Os rebanhos usados para o isolamento do vírus eram formados basicamente por animais de linhagens importadas da Europa, pois os rebanhos nativos que não tiveram contato com animais de origem européia, testados até o momento, têm apresentado resultados negativos. Recentemente, tem sido demonstrado que animais SRD apresentam baixa soroprevalência de CAEV (Castro et al., 1999). Do ponto de vista evolutivo, seria interessante isolar e comparar o genoma de amostras desse tipo de animal com o de amostras isoladas de rebanhos de origem importada. O isolamento do vírus de um rebanho mantido fechado (rebanho nº 3) desde a importação sugere a presença do agente antes mesmo das primeiras descrições clínicas e sorológicas da CAEV no Brasil.
A estratégia utilizada para aperfeiçoar a PCR foi a de diminuir a temperatura de desnaturação e, assim, aumentar a meia-vida da Taq DNA polimerase, o que foi conseguido, pois as temperaturas de 87ºC e 90ºC mostraram-se mais eficientes. Para amplificação do fragmento do genoma do CAEV foram empregadas células obtidas por tripsinização de monocamadas ou de explantes. A utilização de fragmentos de explantes representa a possibilidade de identificação precoce de amostras virais, mesmo antes do surgimento de ECP perceptível e da formação de monocamadas, com número de células suficiente para passagens e para realização de outros ensaios, necessários para identificação do CAEV. Além disso, pode ser útil nos estudos de amostras do CAEV, diminuindo as chances de haver significativa seleção in vitro de variantes genéticas, o que é de relevante significado para estudos filogenéticos de lentivírus de pequenos ruminantes (Leroux et al., 1995).
Resultados positivos à IFD foram observados em células infectadas com todas as amostras isoladas, com amostra CAEV Cork e com o vírus visna/maedi K 1514. Entretanto, a PCR, apesar de ter amplificado o segmento do gene gag de todas as amostras testadas e de CAEV, não amplificou o do vírus visna/maedi. Isto demonstra relação antigênica entre essas amostras, a CAEV Cork e visna/maedi K 1514, bem como a existência de variação da seqüência do visna/maedi K 1514 na região usada para a definição dos "primers", em relação às demais amostras estudadas. A diversidade genética é uma característica típica dos lentivírus (Leroux et al., 1995, 1997), e sua possível relação com o curso das lesões e suas implicações epidemiológicas precisam ser investigadas.
AGRADECIMENTOS
À Fundação Coordenação de Aperfeiçoamento de Pessoal de Nível Superior-CAPES pela concessão de bolsa de Doutorado a Roberto Soares de Castro. Este trabalho foi financiado em parte pela Financiadora de Estudos e Projetos (FINEP).
NOTAS
2 Histopaque 1077, Sigma, USA.
3 Institut Pourquier, Montpellier, France
4 Gentilmente cedidas por Dr. Chebloune do Laboratoire Associé de Recherches sur les Lentivirus chez Les Petits Ruminants - INRA ENVL, France.
5 FANEN, Brasil.
6 MJ Research Inc, USA.
7 Pharmacia, Sweden.
9 BRL Life Technologies, USA
10 Promega, USA.
12 Pharmacia, Sweden.
13 Photodyne, Polaroid, UK.
- ABREU, S.R.O., CASTRO, R.S., NASCIMENTO, S.A. et al. Produçăo de antígeno para diagnóstico da artrite-encefalite caprina e comparaçăo com o Maedi-Visna através da imunodifusăo em ágar gel. Pesq. Vet. Bras., v.18, p.57-60, 1998.
- ADAMS, D.S., OLIVER, R.E., AMEGHINO, E. et al. Global survey of serological evidence of caprine arthritis-encephalitis virus infection. Vet. Rec., v.115, p.493-495, 1984.
- CASTRO, R.S., LEITE, R.C., RESENDE, M. et al. A labelled avidin-biotin ELISA to detect antibodies to caprine arthritis-encephalitis virus in goats sera. Vet. Res. Commun., 1999. (No prelo).
- CLARK, H.F., SHEPARD, C.C. A dialysis technique for preparing fluorescent antibody. Virology, v.20, p.642-644, 1963.
- CLEMENTS, J.E., PAYNE, S. Molecular basis of the pathobiology of lentiviruses. Virus Res., v.32, p.97-109, 1994.
- CRAWFORD, T.B., ADAMS, D.S., CHEEVERS, W.P. et al. Chronic arthritis in goats caused by a retrovirus. Sience, v.207, p.997-999, 1980.
- GOLDMAN, M. Labeling agents and procedures for conjugation. In: FLUORESCENT antibody methods. New York: Academic, 1968. p.97-117.
- HOTZELL, I, BASTOS, S.E., RAVAZZOLO, A.P. et al. Caprine arthritis-encephalitis virus: isolation and identification in Rio Grande do Sul, Brazil. Braz. J. Med. Biol. Res., v.26, p.1175-1179, 1993.
- LEROUX, C., CHASTANG, J., GREENLAND, T. et al. Genomic heterogeneity of small ruminant lentiviruses: existence of heterogeneous populations in sheep and of the same lentiviral genotypes in sheep and goats. Arch. Virol, v.142, p.1125-1137, 1997.
- LEROUX, C., VUILLERMOZ, S., MORNEX, J.F. et al. Genomic heterogeneity in the pool region of ovine lentivirus obtained from bronchoalveolar cells of infected sheep from France. J. Genet. Virol., v.76, p.1533-1537, 1995.
- McKINNEY, M.M., PARKINSON, A. A simple, non-chromatographic procedure to purify immunoglobulins from serum and ascitic fluid. J. Immunol. Methods, v.96, p.271-278, 1987.
- NARAYAN, O., CLEMENTS, J.E., STRANDBERG, J.D. et al. Biological characterization of virus causing leukoencephalitis and arthritis in goats. J. Genet. Virol., v.50, p.69-79, 1980.
- PEREIRA, M.F. Artrite-encefalite caprina a vírus (CAE) - estudo anatomopatológico e imuno-histoquímico em cabras naturalmente infectadas. Belo Horizonte: Escola de Veterinária da UFMG, 1995. 64p. (Dissertaçăo, Mestrado).
- QUERAT, G., BARBAN, V., SAUZE, N. et al. Highly lytic and persistent lentiviruses naturally present in sheep with progressive pneumonia are genetically distinct. J. Virol, v.52, p.672-679, 1984.
- REDDY, P. G., SAPP, W. J., HENEINE, W. Detection of caprine arthritis-encephalites virus by polymerase chain reaction. J. Clin. Microbiol., v.31, p.3042-3043, 1993.
- REED, L. J., MUENCH, H. A simple method of estimating fifty percent end points. Am. J. Hyg., v.27, p.493-497, 1938.
- RIMSTAD, E., EAST, N.E., TORTEN, M. et al. Delayed seroconversion following naturally acquired caprine arthritis-encephalitis virus infection in goats. Am. J. Vet. Res., v.54, p.1858-1862, 1993.
- SARAIVA NETO, A. O, CASTRO, R.S, BIRGEL E.H. et al. Estudo soro-epidemiológico da artrite-encefalite caprina em Pernambuco. Pesq. Vet. Bras., v.15, p.121-124, 1995.
- WEINHOLD, E., MÜLLER, A., LEUCHTE, S. Visna-virus-ähnliche partikel in der kultur von plexus chorioideus-zellen einer ziege mit visna-symptomen. Zb. Vet. Med. B., v.21, p.32-36, 1974.
- WEINHOLD, E., TRIEMER, B. Visna bei de ziege. Zb. Vet. Med. B., v.25, p.525-538, 1978.
Datas de Publicação
-
Publicação nesta coleção
19 Abr 2001 -
Data do Fascículo
Jun 1999
Histórico
-
Recebido
08 Abr 1998