Abstracts
A western blotting (WB) procedure has been developed for detecting antibodies to bovine leukosis virus (BLV) in cattle sera. Two hundred and thirty three serum samples from naturally infected cattle with BLV virus and serial bleedings from experimentally BLV infected cows were used. An agar gel immunodiffusion test (AGID) was used for comparing with the results obtained by WB. The AGID positive sera showed a different degree of reactivity by WB test against the two most important viral antigens (gp51 and p24), or against one of them. Other proteins (gp30, p15, p12 and p10) were not detected with any AGID positive sera, being observed occasionally three bands corresponding to the p24 protein. Using sera obtained by BLV experimental inoculation, the antibodies directed to p24 appeared early (between the 2nd and 4th week post inoculation) and thereafter antibodies to gp51were detected in some animals. The analysis of field serum samples by AGID as compared to WB showed an agreement of 90.9%. Only 1.7% of sera were negative by AGID and positive by WB and 7.2% that were not conclusive by AGID and were defined by WB (4.2% as positive and 3.0% as negative).
Bovine leukosis; western blotting; diagnosis
Um sistema de western blotting (WB) foi desenvolvido para detecção de anticorpos contra o vírus da leucose em soros de bovinos. Foram utilizadas amostras de soros de 233 animais naturalmente infectados e soros de vacas experimentalmente infectadas. O teste de imunodifusão em ágar (AGID) foi usado para comparação dos resultados. Graus diferentes de reatividade foram observados em soros positivos ao AGID, quando testados em WB frente a um ou aos dois antígenos mais importantes (gp51 e p24). Outras proteínas (gp30, p15, p12 e p10) não foram detectadas por nenhum soro positivo ao AGID, sendo que três bandas correspondentes à proteína p24 foram observadas ocasionalmente. Em soros obtidos por inoculação experimental, anticorpos contra a proteína p24 foram detectados entre a segunda e a quarta semanas após a inoculação e, em alguns animais, detectaram-se anticorpos anti-gp51 mais tardiamente. O estudo de soros de campo com AGID e WB mostrou concordância de 90,9% sendo que apenas 1,7% dos soros negativos pelo AGID foram positivos ao WB e 7,2% dos resultados não conclusivos por AGID foram definidos por WB (4,2% como positivos e 3% como negativos).
Leucose bovina; western blotting; diagnóstico
Evaluation of western blotting for the diagnosis of enzootic bovine leukemia (Avaliação da técnica de western blot no diagnóstico da leucose enzoótica bovina)
E.T. Gonzalez1, G.A. Oliva1, J. Norimine2, V. Cid de la Paz1, M.G. Echeverría1
1Laboratorio de Virologia, Faculdad de Ciencias Veterinarias
Universidad Nacional de La Plata
La Plata, Argentina
2Plant and Animal Biotechnology Laboratory, Urbana, IL, USA
E-mail: etgonzal@fcv.medvet.unlp.edu.ar
ABSTRACT
A western blotting (WB) procedure has been developed for detecting antibodies to bovine leukosis virus (BLV) in cattle sera. Two hundred and thirty three serum samples from naturally infected cattle with BLV virus and serial bleedings from experimentally BLV infected cows were used. An agar gel immunodiffusion test (AGID) was used for comparing with the results obtained by WB. The AGID positive sera showed a different degree of reactivity by WB test against the two most important viral antigens (gp51 and p24), or against one of them. Other proteins (gp30, p15, p12 and p10) were not detected with any AGID positive sera, being observed occasionally three bands corresponding to the p24 protein. Using sera obtained by BLV experimental inoculation, the antibodies directed to p24 appeared early (between the 2nd and 4th week post inoculation) and thereafter antibodies to gp51were detected in some animals. The analysis of field serum samples by AGID as compared to WB showed an agreement of 90.9%. Only 1.7% of sera were negative by AGID and positive by WB and 7.2% that were not conclusive by AGID and were defined by WB (4.2% as positive and 3.0% as negative).
Keywords: Bovine leukosis, western blotting, diagnosis
RESUMO
(WB) foi desenvolvido para detecção de anticorpos contra o vírus da leucose em soros de bovinos. Foram utilizadas amostras de soros de 233 animais naturalmente infectados e soros de vacas experimentalmente infectadas. O teste de imunodifusão em ágar (AGID) foi usado para comparação dos resultados. Graus diferentes de reatividade foram observados em soros positivos ao AGID, quando testados em WB frente a um ou aos dois antígenos mais importantes (gp51 e p24). Outras proteínas (gp30, p15, p12 e p10) não foram detectadas por nenhum soro positivo ao AGID, sendo que três bandas correspondentes à proteína p24 foram observadas ocasionalmente. Em soros obtidos por inoculação experimental, anticorpos contra a proteína p24 foram detectados entre a segunda e a quarta semanas após a inoculação e, em alguns animais, detectaram-se anticorpos anti-gp51 mais tardiamente. O estudo de soros de campo com AGID e WB mostrou concordância de 90,9% sendo que apenas 1,7% dos soros negativos pelo AGID foram positivos ao WB e 7,2% dos resultados não conclusivos por AGID foram definidos por WB (4,2% como positivos e 3% como negativos).Palavras-Chave: Leucose bovina, western blotting, diagnóstico
INTRODUCTION
Bovine leukemia virus (BLV) belongs to the Retroviridae family, Oncornavirinae subfamily and is the etiologic agent of the enzootic bovine leukemia (EBL). According to the genome structure of the proviral DNA nucleotide sequence and size and amino acids sequence of the structural and not structural proteins, BLV is more related to the virus producing the human and monkeys leukemias (HTLV-1, HTLV-2, STLV-1) than other oncoviruses (Gallo & Wong- Staal, 1990; Johnson & Kaneene, 1992; Kettmann et al., 1994)
EBL is a chronic and contagious disease that remains asymptomatic for a long period (1 to 8 years), with tumors developing in only a small number of infected animals. The seroprevalence of the infection in the world is variable (Di Giacomo, 1992; Schwartz & Levy, 1994) and the infection incidence seems to be related to the size of the herds, geographical area, race and age of the animals (Kettmann et al., 1994).
The infection with BLV is persistent with blockade at transcriptional level of the virus expression, due apparently to a block factor existing in the plasma, but not in the serum of the infected animals (Gupta & Ferrer, 1982) The transmission of the disease can occur with the inoculation of a very small volume (2 to 10m l) of total blood from an infected animal (Di Giacomo, 1992; Klintevall et al., 1994; Schwartz & Levy, 1994). B-lymphocytes are the white cells population targets for the integration of the viral genome as provirus (Mirsky et al., 1996). The humoral response is strong and persistent and will not permit the elimination of the infection, neither prevents the subsequent progression of the disease to the tumoral phase that is, in all cases, fatal and occurs in a low percentage of animals (Di Giacomo, 1992; Gallo & Wong-Staal, 1990; Heeney et al., 1998; Johnson and Kaneene, 1992)
Several methods have been employed for the diagnosis of EBL such as serum neutralization, complement fixation, radioimmunoassay (RIA), agar gel immunodifussion (AGID), syncytia infectivity assay, variations of the enzyme linked immunosorbent assay (ELISA), polimerase chain reaction (PCR) and western blot (WB) (Miller & Olson, 1972; Miller & Van der Maaten, 1974; McDonald & Ferrer, 1976; Guillemain et al., 1978; Bex et al., 1979; Walker et al., 1987; Ballagi-Pordany et al., 1992; Grover & Guillemain, 1992; Moreno et al., 1992; Jimenez et al., 1995; Ward, 1995; Kittelberger et al., 1996). The serological methods more widely used today are AGID and ELISA (Di Giacomo, 1992; Ward, 1995). The indirect methods are not useful for diagnosis in a recent infection or in the case of young animals with maternal antibodies, or immunodepressed for various causes, including the infection by the virus of the bovine virus diarrhea (BVD).
The envelope glycoprotein gp51, product of the gene env, and core protein p24, product of the gene gag, are the preferential targets against those which the immune system develops antibodies (Kettman et al., 1994)
The objective of this study is to evaluate the use WB technique for serological diagnosis of EBL and to study the kinetics of immune response to viral antigens in an experimental infection, using the AGID technique as reference.
MATERIALS AND METHODS
For this study sera from different sources have been used: 1. positive controls sera from four AGID kits (Pittman Moore INC. Mundelein, USA; Institut Pourquier, Montpellier, France; FCV, Argentine; National Institute of Animal Health, NIAH, Tsukuba, Japan) five sera from naturally infected animals (two of which developing tumor), a BLV negative reference serum and a BVD positive reference serum (Kindly provided by Dra. Lager, INTA, Instituto Nacional de Tecnología Agropecuaria Argentina); 2. field serum samples (233) from naturally infected cattle ranging between two to ten years old. Cattle breeds included Holstein (Holando-Argentine), Hereford, and Aberdeen-Angus, obtained from dairy herds, cabins and beef breeding herds; 3. one hundred and two serum samples from BLV experimentally infected cattle were obtained.
This experiment was carried out with Aberdeen Angus animals from negative herds, three times with intervals of two months previous to the inoculation, analyzed by AGID test. Animals were inoculated with 10ml of heparinized peripheral whole blood diluted in phosphate buffered saline (PBS) from a cow with lymphosarcoma confirmed positive for BLV by AGID test. There were infected six cows by the subcutaneous route and two animals remained uninfected as negative controls. Sera were collected immediately prior to inoculation and then weekly for the first 9 weeks post inoculation (PI) and intermittently after up to 16 weeks.
Cell line FLK (fetal lamb kidney) persistently infected with BLV (kindly provided for Dr. Y. Ando, JICA, Japan) was propagated using Falcon bottles with MEM (Nissui, Pharmaceutical Co. LTD. Japan), supplemented with 0.03g% glutamine, 0.28% tryptose phosphate broth, 100U/ml penicillin, 100mg/ml streptomycin, without fetal calf serum. The supernatant fluid of cultures was harvested every five days up to 10 times consecutively. The supernatant was clarified by centrifugation at 8,500g for 30min. Ammonium sulfate was used for overnight precipitation of viral proteins. After centrifugation at 8.500g for 50min this precipitate was dialyzed against PBS and concentrated with polyethylene glycol (PEG 6000) at 0.1% of the original volume. This crude antigen was used in SDS-PAGE (Walker, 1987). Three crude antigens were available and protein concentration was determinated by a protein assay (Bio-Rad Lab., Richmond, CA, USA).
The antigens were diluted 1:2 in sample buffer under non-reducing conditions (62mM Tris-HCl pH 6.9, 5% SDS, 10% glycerol and 1% bromophenol blue). Electrophoresis was carried out on homogeneous 12% polyacrilamide gel of 1.5mm thickness using a discontinuous tris-glicine buffer system (Grover & Guillemain, 1992). They were subjected to 100V, transferred to nitrocellulose paper NC (Bio-Rad 0,45m ) with blotting apparatus (Advantec ETB-15) overnight at 4°C at 20V. Membranes were blocked 1h at 37°C in TBS (Tris buffer saline) containing 3% gelatin. After washings with TBS, the NC papers were incubated with serum samples diluted 1/100 in TTBSG (TBS 0.02% Tween 20 and 1% gelatin) at 37°C during 2h. Then the NC were washed again and incubated with peroxidase conjugated rabbit anti-cow immunoglobulin (Sigma) diluted 1/1000, during 1h at 37°C. The reaction was developed with 30mg diaminobenzidine in 100ml of TBS containing 20ml of 30% H2O2, placing the sheet paper membrane in running tap water for 5min to stop the reaction.
All serum samples were analyzed by agar gel immunodiffusion (AGID) using diagnostic equipment produced by our Laboratory (FCV, UNLP, Argentine, approved by the National Service of Animal Health, SENASA Argentine).
RESULTS
The crude antigen preparation, which was used in WB test, consisted of concentrated cell culture fluid. It contained culture medium without serum. We used three different batches of antigens containing between 33-60mg of protein per ml. All of the antigens were positive by AGID against the reference positive sera, but differences within the three batches were observed when assayed by in WB. Antigens 1 and 3 showed a stronger reaction for the most important proteins (p24 and gp51) than antigen 2, which exhibited poor staining for these bands. Fig. 1 shows reactivity profile of the reference sera using antigen 1 under non-reducing condition, showing correspondence with positive control serum used in AGID. The intensity of the reaction for the different antigen proteins was variable but all reacted for the p24 and the gp51 regions. No reactivity was observed with negative control serum. None of the protein bands reacted with bovine sera specific to BVD.
Reactivity against the internal proteins p15, p12, p10 was not detectable in WB with any sera (including seropositives symptomatic or asymptomatic animals).
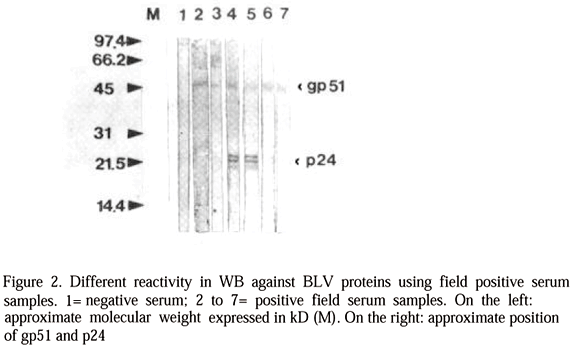
A different degree of reactivity of the positive sera by WB to the most important viral antigens (gp51 and p24), being observed in some instances three bands corresponding to p24 (Fig. 2). Moreover gp30 was not detected with any positive serum, being variable the reactivity for a greater molecular weight protein (70 KD).
The analysis of 233 field serum samples by AGID and WB showed that in 90.9% of the cases there was an agreement of both techniques. There were 1.7% sera that were negative by AGID and positive by WB and 7.2% that were non-conclusive by AGID and were defined by WB (4.2% as positive and 3% as negative) as shown in Table 1.
Table 2 shows the WB results corresponding to the BLV experimentally infected catle. The antibodies for the p24 appeared between the second and fourth week PI. The antibodies aganst the gp51 were detected between the second and the sixth week PI. The same sera were analyzed by AGID and started to be positive between the fourth and sixth week PI. One of them remained weakly positive during all the experiment. The control animals were always negative for both techniques.
DISCUSSION
The WB protein profiles identified in this study corresponded to the genes gag and env BLV products as described previously (Grover & Guillemain, 1992; Johnson & Kaneene, 1992; Kettmann et al., 1994). The gag gene expression, results in the synthesis from four non-glycosilated proteins (p24, p14, p12 and p10) as identified by polyacrylamide gel electrophoresis and RIA (Mamoun et al., 1983; Uckert et al., 1986). For these non-glycosilated proteins, p24 region was the only detected by WB. In conformity with other authors (Mager et al., 1994; Jimenez et al., 1995), the reactivity of some sera for the p24 region permitted to differentiate one to three bands employing the same batch of antigen. This suggests small differences in the corresponding molecular weight of this protein, associated with structural variations of the polipeptide chain, due to special cleavage in the protein synthesis, which would be detected with different intensity by different serum samples according to the particular immune response involved. On the other hand, working with one positive serum and different antigen batches, it was observed variability in the detection of these bands. This variability probably is due to molecular weight heterogeneity of these proteins, possibly by an alteration in the antigen production process and/or subsequently by WB. In conformity with this, Mager et al. (1994) detected up to four bands using monoclonal antibodies against p24. Storage of the antigen for long periods at 20°C resulted in the appearance of two additional bands of 26 and 28 kDa (Walker et al., 1987)
In respect to the product of gene env the detection of two glycoproteins, gp51 and gp30, components of the viral envelope and involved directly in the viral infectivity (Johnson & Kaneene, 1992; Kettmann et al., 1994; Schwartz & Levy, 1994) is expected. In this experiment the detection of the gp51 was not a constant finding for the positive serum and we could not detect the gp30 with any serum. This is not in line with the observation of Moreno et al. (1992) and Jimenez et al. (1995) which detected gp30 together with other bands.
Some positive sera had reactivity for a greater molecular weight protein (70 kD). It is possible that this protein represents an intermediary of the gp51 and gp30 that is released in the cellular culture during assembly of the whole particles (Heeney et al., 1998). Kittelberger et al. (1996) also points out that this band is unrelated to the true serological status.
Analyzing 232 field serum samples by the AGID and WB techniques, the results demonstrate that all positive sera for AGID were positive by WB. We detected antibodies directly against the envelope glycoprotein gp51 and/or against the internal protein p24. This is in agreement with Kettmann et al. (1994), which indicated that the gp51 together with the p24 are the most potent antigens. However, in our results the degree of reactivity for these proteins was not the same in all cases. Furthermore, WB defined undetermined sera by AGID test, as positive or negative.
Although the WB antigen clearly contained the viral surface glycoprotein gp51, it was not usually detected in all the field positive serum samples. The known immunodominat gp51 failed as a reliable indicator for the serological status of the sera in blots when compared to the results on the same sera applying AGID as reference test. According to Grover & Guillemain (1992), a strong reaction was detected against the major viral core antigen p24. The reactivity bands against the internal proteins, p15, p12, and p10, were not detectable in WB for any sera (including seropositives symptomatic or asymptomatic animals). This is in contrast with Heeney et al. (1998) which indicated that the presence of antibodies to proteins of low molecular weight is occasional for asymptomatic animals, appearing however constant in animal with symptomatology.
Serum of animal associated with lymphoma showed a stronger reactivity than normal infected cattle, being related this fact to the chronic state of the infection and to the greater possibility of provirus expression.
Using sera obtained by experimental inoculation, the antibodies directed to the p24, appeared earlier (between the 2nd and 4th week PI) and thereafter to the gp51 in some animals, being of variable intensity for the latter. This would be in agreement with Kittelberger et al. (1996), which compared WB with ELISA and AGID, and demonstrated a loss of reactivity of many sera against gp51 by WB, due probably to the labile nature of this antigen molecule. The gp51 molecule is highly glycosylated and serum antibodies in cattle seem to be directed only against a limited number of such glycosylated sites (Johnson & Kaneene, 1992; Kettmann et al., 1994). If these sites are composed of labile molecules, antibody reactivity may be lost through denaturation. It is likely that during sample preparation, which includes boiling in the presence of SDS at pH 8.9, and electrophoresis, produced hydrolysis of labile bonds between sugar residues. The differences between the reaction in AGID, in which the antigen has a predominance of gp51 and the reaction in blots, are probably due to such denaturation processes.
The same sera analyzed by AGID were WB positive between the 4th and 6th week PI. In one animal the AGID results were weakly positive during all the experiment, but were clearly positive for WB from the 2nd week onwards.
CONCLUSIONS
1. The reactivity of AGID positive BLV sera in WB analysis is clear for the major proteins (gp51 and p24), against both or against only one of them. Other proteins (gp30, p15, p12 and p10) were not detected with any positive serum. 2. The presence of p24 protein can be demonstrated by the appearance of 1 to 3 bands and the reactivity against gp51 is dependent on the integrity of this labile molecule. The integrity of gp51 can depend on the antigen batch and/or degradation during electrophoresis. 3. WB is an alternative technique that can elucidate some doubtful results obtained by other serological methods like AGID.
ACKNOWLEDGEMENTS
The authors are grateful to Dr. Alejandro Valera, from the Veterinary Diagnostic Center. This work was supported by grants of the Secretary of Sciences and Technology, La Plata University and Japan International Cooperation Agency (JICA).
REFERENCES
- BALLAGI-PORDANY, A., KINTEVALL, K., MERZA, M. et al. Direct detection of bovine leukemia virus infection: practical applicability of a double polymerase chain reaction. J. Vet. Med. Ser. B., v.39, p.69-77, 1992.
- BEX, F., BRUCK, C., MAMMERICKX, M. et al. Humoral antibody response to bovine leukemia virus infection in cattle and sheep. Canine Res., v.39, p.1118-1123, 1979.
- DI GIACOMO, R.F. The epidemiology and control of bovine leukemia virus infection. Vet. Med, v.3, p.248-257, 1992.
- GALLO, R, WONG-STAAL, F., (Eds). Retrovirus biology and human disease New York, 1990.
- GROVER, Y.P., GUILLEMAIN, B. An immunoblotting procedure for detection of antibodies against Bovine Leukemia virus in cattle. J. Vet. Med. Ser. B, v.39, p.48-52, 1992.
- GUILLEMAIN, B., MAMOUN, R., LEVY, D. et al. Early polykaryocytosis inhibition (EPI). A simple quantitative test for the detection of bovine leukemia virus infection in cattle. Eur. J. Cancer, v.4, p.811-827, 1978.
- GUPTA, P., FERRER, J. Expression of bovine leukemia virus genome is blocked by a non inmunoglobulin protein in plasma from infected cattle. Science, v.215, p.405-407, 1982.
- HEENEY, J., VALLI, V.E., MONTESANI, J. Alterations in humoral immune response to bovine leukemia virus antigens in cattle with lymphoma. J. Gen. Virol, v.69, p.659-666, 1998.
- JIMENEZ, C., BONILLA, J.A., DOLZ, G. et al. Bovine leukemia-virus infection in Costa Rica. Zentralb. Veterinaermed. B., v.42, p.385-390, 1995.
- JOHNSON, R., KANEENE, J. Bovine Leukemia Virus and Enzootic Bovine Leukosis. Vet. Bull, v.62, p.287-312, 1992.
- KETTMANN, R., BURNY, A., CALLEBAUT, I. et al. The retroviridae, New York: Plenum 1994. p.39-81.
- KITTELBERGER, B.J., LAYBOURN, B.J., DIACK, D.S. et al. Evaluation of electrophoretic immunoblotting for the detection of antibodies against the bovine leukosis virus in cattle J. Virol. Methods, v.61, p.7-22, 1996.
- KLINTEVALL, K., BALLAGY-PORDANY, A., NÄSLUND, K. et al. Bovine leukemia virus: rapid detection of proviral DNA by nested PCR in blood and organs of experimentally infected calves. Vet. Microbiol, v.42, p.191-204, 1994.
- MAGER, A., MASENGO, R., MAMMERICKX, M. et al. T cell proliferative response to bovine leukemia virus (BLV): identification of T cell epitopes on the major core protein (p24) in BLV-infected cattle with normal haematological values. J. Gen. Virol, v.75, p.2223-2231, 1994.
- MAMOUN, R.Z., ASTIER, T., GUILLERMAIN, B. et al. Bovine lymphosarcoma: expression of BLV-related proteins in cultured cells. J. Gen. Virol, v.64, p.1895-1905, 1983.
- MCDONALD, C., FERRER, J.F. Detection, quantitation and characterization of the major internal virion antigen of the bovine leukemia virus by radioimmunoassay. J. Nat. Cancer Inst, v.57, p.875-882, 1976.
- MIRSKY, M.L., OLMSTEAD, C.A., DA, Y. et al. The prevalence of proviral bovine leukemia virus in peripheral blood mononuclear cells at two subclinical stages of infection. J. Virol, v.4, p.2178-2183, 1996.
- MILLER, J.M., OLSON, C. Precipitating antibody to an internal antigen of the C-type virus associated with bovine lymphosarcoma. J. Nat. Cancer Inst, v.49, p.1459-1462, 1972.
- MILLER, J.M., VAN DER MAATEN, M.J. A complement-fixation test for the bovine leukemia (C-type). J. Nat. Cancer Inst, v.53, p.1699-1702, 1974.
- MORENO, E., DOLZ, G., BONILLA, J. et al. Serological studies on Bovine Leukemia Virus (BLV) infection in Costa Rica by ELISA, Immunodiffusion and Western Blot tests. In: REGIONAL NETWORK FOR LATIN AMERICA ON ANIMAL DISEASE DIAGNOSIS USING IMMUNOASSAY AND LABELLED DNA PROBE TECHNIQUES, 1990, Heredia. Proceedings.. Vienna: International Atomic Energy Agency, 1992. p.99-114.
- SCHWARTZ, I., LEVY, D. Pathobiology of bovine leukemia virus. Vet. Res, v.25, p.521-536, 1994.
- UCKERT, W., HERTLING, I., KRAFT, R. et al. Structural components of bovine leukemia virus: further biochemical and immunological characterization of major structural proteins and glycoproteins. Virus Res, v.4, p.343-356, 1986.
- WALKER, P.J., MOLLOY, J.B., RODWELL, B.J. A protein immunoblot test for detection of bovine leukemia virus p24 antibody in cattle and experimentally infected sheep. J. Virol. Methods, v.15, p.201-211, 1987.
- WARD, M.P. Serological studies of bovine leukemia virus infection in Queensland beef cattle. Aust. Vet. J, v.72, p.71-72, 1995.
Publication Dates
-
Publication in this collection
18 Apr 2001 -
Date of issue
Aug 1999
History
-
Received
07 May 1998