Resumos
Foram estudadas silagens de quatro híbridos de sorgo, com diferentes teores de umidade no colmo combinados com teor alto ou baixo de tanino, em silos de laboratório feitos de tubos PVC, lacrados com fita crepe e dotados de uma válvula tipo Bunsen. Usaram-se 24 tratamentos com quatro repetições cada, sendo quatro híbridos (T1=colmo suculento e baixo tanino, T2=colmo seco e baixo tanino, T3=colmo seco e alto tanino, T4= colmo suculento e alto tanino) combinados com seis tempos para abertura do silo, sendo o primeiro tempo antes de ensilar (P1), seguido de 1, 7, 14, 28 e 56 dias de ensilagem. Determinaram-se os teores de matéria seca (MS), proteína bruta (PB), nitrogênio amoniacal (N-NH3) e tanino. Utilizou-se o teste de Tukey para comparação entre médias. Todos os híbridos produziram N-NH3 ao longo da fermentação. Os valores obtidos da fração N-amoniacal/N-total encontram-se abaixo dos valores citados na literatura. Silagens de híbridos com tanino alto apresentaram menores valores de N-amoniacal e maiores teores de matéria seca (P<0,05) que silagens de baixo tanino.
Silagem; sorgo; tanino; composto nitrogenado
Four hybrid forage sorghums with different tannin concentrations and moisture in the stem were ensiled in laboratory silos made of PVC tubes. The silos were opened 1 (P2), 7 (P3), 14 (P4), 28 (P5) e 56 (P6) days after ensiling, in order to evaluate fermentation patterns and others parameters. Twenty-four treatments were used, with four repetitions each: 4 hybrids (T1=moist stem/low tannin, T2=not moist stem/low tannin, T3=not moist stem/high tannin, T4= moist stem/high tannin) and with six periods, since the forages before ensiling were also studied. Dry matter content (DM), crude protein (CP), ammonia nitrogen (N-NH3) and tannin were determined. Data were evaluated by analysis of variance, with mean separation achieved using Tukey statistical test. All sorghums had N-NH3 production during the fermentation, which was lower in high tannin sorghums. The N-NH3 concentrations were lower than those in the literature. High tannin sorghums had higher dry matter contents.
Silage; sorghum; tannin; nitrogenous compound
Silagem de sorgo de porte baixo com diferentes teores de tanino e de umidade no colmo. III- Quebra de compostos nitrogenados
(Forage sorghum silage with different tannin concentrations and moisure in the stem. III- Effect on nitrogenous compounds)
L.C. Gonçalves1, N.M. Rodriguez1, F.S. Nogueira2, A.L.C.C. Borges1*, C.P. Zago3
1Escola de Veterinária da UFMG
Caixa Postal 567
30123-970 Belo Horizonte, MG
2 Med. Vet., MZ, Belo Horizonte, MG
3Pesquisador Sementes Agroceres, Capinópolis, MG
Recebido para publicação em 2 de março de 1999.
Colaborador: I. Borges
Autor para correspondência
E-mail: anaglaub@vet.ufmg.br
RESUMO
Foram estudadas silagens de quatro híbridos de sorgo, com diferentes teores de umidade no colmo combinados com teor alto ou baixo de tanino, em silos de laboratório feitos de tubos PVC, lacrados com fita crepe e dotados de uma válvula tipo Bunsen. Usaram-se 24 tratamentos com quatro repetições cada, sendo quatro híbridos (T1=colmo suculento e baixo tanino, T2=colmo seco e baixo tanino, T3=colmo seco e alto tanino, T4= colmo suculento e alto tanino) combinados com seis tempos para abertura do silo, sendo o primeiro tempo antes de ensilar (P1), seguido de 1, 7, 14, 28 e 56 dias de ensilagem. Determinaram-se os teores de matéria seca (MS), proteína bruta (PB), nitrogênio amoniacal (N-NH3) e tanino. Utilizou-se o teste de Tukey para comparação entre médias. Todos os híbridos produziram N-NH3 ao longo da fermentação. Os valores obtidos da fração N-amoniacal/N-total encontram-se abaixo dos valores citados na literatura. Silagens de híbridos com tanino alto apresentaram menores valores de N-amoniacal e maiores teores de matéria seca (P<0,05) que silagens de baixo tanino.
Palavras-Chave: Silagem, sorgo, tanino, composto nitrogenado
ABSTRACT
Four hybrid forage sorghums with different tannin concentrations and moisture in the stem were ensiled in laboratory silos made of PVC tubes. The silos were opened 1 (P2), 7 (P3), 14 (P4), 28 (P5) e 56 (P6) days after ensiling, in order to evaluate fermentation patterns and others parameters. Twenty-four treatments were used, with four repetitions each: 4 hybrids (T1=moist stem/low tannin, T2=not moist stem/low tannin, T3=not moist stem/high tannin, T4= moist stem/high tannin) and with six periods, since the forages before ensiling were also studied. Dry matter content (DM), crude protein (CP), ammonia nitrogen (N-NH3) and tannin were determined. Data were evaluated by analysis of variance, with mean separation achieved using Tukey statistical test. All sorghums had N-NH3 production during the fermentation, which was lower in high tannin sorghums. The N-NH3 concentrations were lower than those in the literature. High tannin sorghums had higher dry matter contents.
Keywords: Silage, sorghum, tannin, nitrogenous compound
INTRODUÇÃO
Do nitrogênio total das forragens, 75 a 90% encontram-se na forma de proteína. A fração não protéica é composta principalmente de aminoácidos livres e amidas (glutamina e asparagina) e pequenas concentrações de outros compostos sem significado nutricional. A concentração de N-amoniacal em forragens é usualmente menor que 1% (Oshima & McDonald, 1978; McDonald et al., 1991). No entanto, a degradação protéica por enzimas das plantas e a ação de bactérias lácticas, entéricas e clostrídios alteram a composição da fração nitrogenada na silagem (Van Soest, 1994; Rodriguez et al., 1998).
A extensão da hidrólise protéica por enzimas depende do potencial proteolítico da forragem e da velocidade de queda do pH. O potencial proteolítico é a medida da atividade total das proteinases, da disponibilidade das proteínas e da afinidade das proteinases pela proteína das plantas (McKersie, 1985). O conteúdo de matéria seca e a temperatura também são importantes, segundo Henderson (1993).
O nitrogênio amoniacal (N-amoniacal) das silagens origina-se da ação de bactérias sobre a fração solúvel e sobre as proteínas. No primeiro caso, há uma estreita correlação entre essas bactérias e a produção de lactato (Moisio & Heikonen, 1994). A ação de bactérias sobre as proteínas é geralmente associada a fermentações secundárias. Embora se tenha como certo que essas bactérias são inibidas em pH próximo de 4,0, Luis & Ramirez (1988) observaram que, além das leveduras, alguns grupos de bactérias entéricas, também produtoras de N-amoniacal, permaneciam ativas em pH abaixo de 4,0.
A produção de grandes quantidades de N-solúvel altera o curso da fermentação, uma vez que os aminoácidos básicos, aminas e N-amoniacal, produtos da hidrólise protéica, podem retardar a queda do pH. Os níveis de N-amoniacal em silagens bem fermentadas são usualmente inferiores a 10% do nitrogênio total (N-total), sendo a amônia derivada principalmente da deaminação de aminoácidos específicos e amidas, e da redução de nitratos pelas bactérias lácticas (Fairbairn et al., 1992). No entanto, em silagens com evidências de fermentações secundárias, o nitrogênio solúvel (N-solúvel) e o N-amoniacal advêm da ação de microrganismos responsáveis por essas fermentações. As concentrações desses metabólitos são conseqüência da extensão da atividade de cada grupo de microrganismos no ambiente do silo (Van Soest, 1994). Nesses casos, o catabolismo dos aminoácidos ocorre por três processos bioquímicos: deaminação, descarboxilação e reações de oxidação e redução, com liberação de amônia (McDonald et al., 1991).
O tanino dos alimentos tem a capacidade de formar complexos com a fração protéica, reduzindo sua degradabilidade, e conseqüentemente sua disponibilidade (Van Soest, 1994). Além disso, os taninos podem provocar inibição de enzimas bacterianas celulolíticas e ureolíticas (Kumar & Singh, 1984; Marinho, 1984) e alterações entre os ácidos nos processos fermentativos do rúmen (Nuñez-Hernandez et al., 1991; Van Hoven & Furstenburg, 1992). De fato, Albrecht & Muck (1991), estudando aspectos fermentativos de silagens de leguminosas, observaram que os taninos inibiram a proteólise. O mesmo foi observado por Borges (1995) e Rodriguez et al. (1998).
Este trabalho teve como objetivo avaliar as variações nos valores de tanino e sua interferência na quebra de compostos nitrogenados na ensilagem.
MATERIAL E MÉTODOS
O material utilizado e o processo de coleta de amostras foram descritos por Rodriguez et al. (1999). As amostras pré-secas foram moídas em moinho estacionário Thomas-Wiley, modelo 4, utilizando-se peneira de 1mm. Em seguida, foram guardadas em vidros com tampa para as demais determinações de laboratório. Parte do material retirado de cada silo foi prensado, utilizando-se uma prensa hidráulica Carver, modelo C, para extração do suco, que foi utilizado imediatamente para análise de N-amoniacal, pela utilização de óxido de magnésio e cloreto de cálcio (AOAC, 1980). Nas amostras pré-secas, determinaram-se a matéria seca em estufa a 105oC (AOAC, 1980), a proteína bruta (método de Kjeldhal, segundo AOAC, 1980) e os taninos (Watterson & Butler, 1983) sem a utilização de PVP (polivinilpirrolidona).
Os híbridos (tratamentos) divididos em quatro grupos, de acordo com o teor de tanino e de suculência do colmo, utilizando-se seis tempos para a abertura dos silos, e as análises estatísticas foram descritos por Rodriguez et al. (1999).
Adotou-se o esquema 4´ 5 para N-amoniacal (tratamentos ´ tempos) e 4 ´ 6 para as demais características. As correlações entre as variáveis dependentes estudadas foram determinadas utilizando-se a comparação entre médias.
RESULTADOS E DISCUSSÃO
A Tab. 1 mostra os teores de proteína bruta das silagens. Os valores protéicos no P1 encontram-se próximos aos citados por Zago (1991) para híbridos de sorgos graníferos no estádio de grão farináceo.
Não houve diferença estatística (P>0,05) na concentração de proteína bruta entre as médias dos tempos de abertura, embora os híbridos tenham apresentado diferentes valores protéicos. No entanto, essa informação terá maior significado nutricional quando acompanhada de um método que leve em conta as alterações nos diferentes componentes da fração nitrogenada. Nas silagens de forma geral, parte do nitrogênio total (N-total) encontra-se na forma solúvel (N-solúvel), produto da atividade enzimática das plantas, e parte na forma de N-amoniacal, produto da atividade microbiana sobre o N-solúvel (Henderson, 1993; Moisio & Heikonen, 1994). A utilização do fator 6,25 para transformar o N-total em proteína bruta não leva em conta essa proteólise, superestimando a fração proteína em silagens (Sigurd et al., 1987).
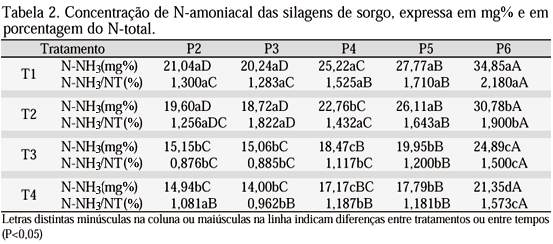
Na Tab. 2 encontram-se as concentrações de N-amoniacal dos híbridos, de P2 a P6. Mesmo constatando que todos os valores obtidos da fração N-amoniacal/N-total encontram-se abaixo dos valores citados na literatura, não se pode deixar de observar que as silagens de variedades de sorgo com tanino alto apresentaram menores valores de N-amoniacal (P<0,05) que as silagens de baixo tanino. Os valores expressos em N-amoniacal/N-total foram obtidos pela relação entre os valores em mg% e o N-total obtido pela determinação da proteína bruta.
Concentrações de 6% de N-amoniacal/N-total nas silagens de sorgo com 23,6% de matéria seca, feitas em tambores de 200 litros, foram observadas por Tjandraamadja et al. (1993). Ojeda & Diaz (1992), utilizando microssilos com capacidade para 400g de forragem verde encontraram 14,6% de N-amoniacal/N-total em silagens obtidas de sorgo ensilado com 8,45% de proteína bruta e 28,32% de matéria seca. Fairbairn et al. (1992), ensilando milho em microssilos de PVC com 25cm de comprimento e 7,6cm de diâmetro encontraram 2,57; 3,23; 3,94; 5,4 e 7,1 de N-amoniacal/N-total aos 1, 7, 14, 42 e 90 dias de ensilagem, respectivamente. Bolsen et al. (1993), ensilando milho e sorgo, encontraram 100 e 70 mg% de N-amoniacal, respectivamente. No entanto, Meeske et al. (1993) relatam valores de N-amoniacal/N-total inferiores a 0,5% para silagens de sorgo com 8,19% de proteína bruta e 27,6% de matéria seca, utilizando como silos jarras de vidro com capacidade para 1,5 litro, providas de válvula para escape de gases.
A influência dos teores de matéria seca sobre a proteólise foi revisada por McKersie (1985), Muck (1988) e McDonald (1991). Entretanto, observando os valores de proteína bruta e matéria seca dos híbridos na Tab. 3, não se pode creditar as diferenças nas concentrações de N-amoniacal entre os tratamentos apenas às diferenças nos teores de matéria seca, uma vez que a correlação da matéria seca com a proteólise se fez mais presente em forragens com baixos teores de carboidratos fermentáveis (Meeske et al., 1993), com concentrações protéicas acima de 15% (Moisio & Heikonen, 1994), ou com teores de matéria seca total abaixo de 20% (McKersie, 1985; Fisher & Burns, 1987; Henderson, 1993). Meeske et al. (1993) obtiveram silagens com menores valores de N-amoniacal quando ensilaram sorgos, embora estes tivessem menores teores de matéria seca (21,4 ´ 27,6% matéria seca). No presente trabalho, o T1, de colmo suculento, apresentou valores superiores de N-amoniacal, provavelmente devido a sua maior concentração de carboidratos solúveis. Entretanto, o mesmo não foi observado para o T4, também de colmo suculento.
A inexistência aparente de fermentações secundárias (Van Soest, 1994) e a relativamente baixa concentração protéica do material ensilado (Moisio & Heikonen, 1994) parecem ter sido os fatores que explicam as pequenas concentrações de N-amoniacal encontradas.
Neste trabalho, os híbridos T3 e T4, de alto tanino, apresentaram silagens com teores significativamente inferiores de N-amoniacal/N-total. A ação dos taninos na redução da relação N-amoniacal/N-total poderia se fazer sentir por três principais mecanismos: diminuição da disponibilidade da proteína a ser solubilizada e/ou inibição da ação da enzima das plantas, e ainda inibição da atividade das enzimas bacterianas. Nos dois primeiros casos, a queda nas concentrações de N-solúvel traria como conseqüência a queda nas concentrações de N-amoniacal. No entanto, especulações sobre a deficiência de substrato N-solúvel como limitante para produção de amônia parecem infundadas. Mesmo em silagens bem preservadas, as concentrações de N-solúvel são altas, uma vez que a atividade enzimática persiste, mesmo que em pequena escala, em pH inferior a 4, sendo intensa e de difícil controle nas primeiras 48h de ensilagem (McKersie, 1985). Contudo, não se questiona a inibição das enzimas proteolíticas das bactérias (Robbins et al., 1987). Essas poderiam ter sua capacidade de produção de N-amoniacal reduzida pelos taninos. A redução da proteólise, com menores concentrações de N-amoniacal, também foi constatada por Albrech & Muck (1991), que ensilaram leguminosas com altos teores de taninos. Assim, as menores concentrações de N-amoniacal nos híbridos T3 e T4, quando comparados com T1 e T2, provavelmente foram devidas aos maiores teores de tanino dos primeiros, assim como de seus conteúdos de matéria seca superiores. Entretanto, a influência dos taninos na diminuição da proteólise no processo de ensilagem pode também estar influenciando o teor de matéria seca.
Fairbairn et al. (1992), ao estudarem os padrões de degradação protéica no milho, encontraram concentrações de N-amoniacal de 0,86% no momento da ensilagem, 1,64% após 8h de ensilagem, 2,42% após 16h, 3,23% após sete dias e 5,43 após 42 dias. Levando-se em conta que as silagens de sorgo e milho teriam padrões de fermentação semelhantes, a inexistência de dados sobre a concentração de N-amoniacal em tempos anteriores a 24h de ensilagem e sobre a fração N-solúvel limita a discussão sobre as interações entre taninos e frações nitrogenadas.
CONCLUSÕES
Todos os híbridos apresentaram produção de N-NH3 ao longo da fermentação. Os valores obtidos da fração N-amoniacal/N-total encontram-se abaixo dos valores citados na literatura. Silagens de híbridos com tanino alto apresentaram menores valores de N-amoniacal (P<0,05) que silagens de baixo tanino. Da mesma forma, os conteúdos de matéria seca foram superiores nos híbridos de alto tanino.
REFERÊNCIAS BIBLIOGRÁFICAS
- ALBRECH, K.A., MUCK, R.E. Proteolysis in ensiled forage legumes that vary in tannin concentration. Crop Sci., v.31, p.464-469, 1991.
- AOAC. Official methods of analysis 13.ed. Washington: AOAC, 1980, 1015p.
- BOLSEN, K.K. et al. Rate and extent of top spoilage losses in horizontal silos. J. Dairy Sci., v.76, p.2940-2962, 1993.
- BORGES, A L.C.C. Qualidade de silagens de híbridos de sorgo de porte alto com diferentes teores de tanino e de umidade no colmo, e seus padrões de fermentação Belo Horizonte: Escola de Veterinária da UFMG, 1995. 104p. (Tese, Mestrado).
- FAIRBAIRN, R.L., ALLI, I., PHILLIP, L.E. Proteolysis and amino acid degradation during ensilage of untreated or formic acid-treated lucerne and maize. Grass For. Sci., v.47, p.382-390, 1992.
- FISHER, D.S., BURNS, J.C. Quality analysis of summer-annual forages. II. Effects of forage carbohydrate constituents on silage fermentation. Agron. J., v.79, p.242-248, 1987.
- HENDERSON, N. Silage additives. Anim. Feed Sci. Technol., v.45, p.35-56, 1993.
- LUIS, L., RAMIREZ, M. Evolución de la flora microbiana en un ensilaje de king grass. Pastos For., v.11, p.249-253, 1988.
- KUMAR, R., SINGH, M. Tannins: their adverse role in ruminant nutrition. J. Agric. Food Chem., v.32, p.447-453, 1984.
- MARINHO, A.A.M. Influência dos taninos no comportamento dos microrganismos e suas implicações nas transformações microbianas no trato gastrintestinal dos ruminantes. Rev. Port. Ciênc. Vet., v.79, p.5-21, 1984.
- McDONALD, P, HENDERSON, A.R., HERON, S. The biochemistry of silage 2.ed. Marlow: Chalcombe, 1991. 340p.
- McKERSIE, B.D. Effect of pH on proteolysis in ensiled legume forage. Agron. J., v.77, p.81-86, 1985.
- MEESKE, R., ASHBELL, G., WEINBERG, Z.G. et al. Ensiling forage sorghum at two stages of maturity with the addition of lactic acid bacterial inoculants. Anim. Feed Sci. Technol., v.43, p. 165-175, 1993.
- MOISIO, T., HEIKONEN, M. Lactic acid fermentation in silage preserved with formic acid. Anim. Feed Sci. Technol., v.47, p.107-124, 1994.
- MUCK, R.E. Factors influencing silage quality and their implications for management. J. Dairy Sci., v.71, p.2992-3002, 1988.
- NUÑEZ-HERNANDEZ, G., WALLACE, J.D., HOLECHEK, J.L. et al. Condensed tannins and utilization by lambs and goats fed low-quality diets. J. Anim. Sci., v.69, p.1167-1177, 1991.
- OJEDA, F., DIAZ, D. Estudio de diferentes proporciones de sorgo y dolichos ensilados com y sin preservantes. Pastos For., v.15, p.77-87, 1992.
- OSHIMA, M., McDONALD, P. A review of changes in nitrogenous compounds in herbages during ensiling. J. Sci. Food Agric., v. 29, p.497-505, 1978.
- ROBBINS, C.T. et al. Role of tannins in defending against ruminants: reduction in dry matter digestion. Ecology, v.68, p.1606-1615, 1987.
- RODRIGUEZ, N.M., BORGES, A.L.C.C., GONÇALVES, L.C. et al. Silagens de sorgo de porte alto com diferentes teores de tanino e de umidade no colmo. III Quebra de compostos nitrogenados. Arq. Bras. Med. Vet. Zoot, v.50, p.161-165, 1998.
- RODRIGUEZ, N.M. GONÇALVES, L.C., NOGUEIRA, F.S. et al. Silagem de sorgo de porte baixo com diferentes teores de tanino e de umidade no colmo. I pH e teores de matéria seca e de ácidos graxos durante a fermentação. Arq. Bras. Med. Vet. Zootec., v.51, p.485-490, 1999.
- SAS. Users guide: statistics Cary, NC: SAS Institute Inc., 1990 (versão 6.0).
- SIGURD, B., BECH-ANDERSEN, S., EGGUM, B.O. A critical view on the conversion factor 6,25 from total nitrogen to protein. Acta Agric. Scandinavia, v.37, p.299-304, 1987.
- TJANDRAATMADJA, M., McRAE, I.C., NORTON, B.W. Intake and digestibility of sorghum silage by goats. Anim. Feed Sci. Technol., v.41, p.171-179, 1993.
- VAN HOVEN, W., FURTENBURG, D. The use of purified condensed tannin as a reference in determining its influence on rumen fermentation. Comparative Biochemistry and Physiology, v.101, p.381-385, 1992.
- VAN SOEST, P.J. Nutritional ecology of the ruminant 2.ed. Ithaca, New York: Cornell University Press, 1994. 476 p.
- WATTERSON, J.J., BUTLER, L.G. Ocucurence of an unusual leucoanthocyanidin and absence of proanthocynidins in sorghum leaves. J. Agric. Food Chem., v.31, p.41-45, 1983.
- ZAGO, C.P. Cultura de sorgo para produção de silagem de alto valor nutritivo. In: SIMPÓSIO SOBRE NUTRIÇÃO DE BOVINOS, 4, 1991, Piracicaba. Anais... Piracicaba: FEALQ. 1991. p.169-217.
Datas de Publicação
-
Publicação nesta coleção
05 Abr 2001 -
Data do Fascículo
Dez 1999
Histórico
-
Recebido
02 Mar 1999