Resumos
Avaliou-se o desempenho de fêmeas bovinas doadoras de embrião nos primeiros dois anos de trabalho em um centro do programa multiple ovulation and embryo transfer (MOET). Utilizaram-se novilhas prenhes de recente importação de clima moderado para o Nordeste Brasileiro. Compararam-se os resultados de superovulação em 1996, primeiro ano de adaptação dos animais, e 1997 com AUSA-Superov (FSH) e Folligon (eCG). O número de CL contados no ovário foi 3,3 (FSH) e 4,2 (eCG) em 1996 e 8,2 (FSH) e 9,6 (eCG) em 1997, o de óvulos e embriões degenerados coletados de 2,8 (FSH) e 3,8 (eCG) em 1996 e 6,5 (FSH) e 7,4 (eCG) em 1997, o de embriões de boa qualidade de 1,9 (FSH) e 2,3 (eCG) em 1996 e 5,5 (FSH) e 4,0 (eCG) em 1997. A eficiência da coleta foi de 40% (FSH) e 35% (eCG) em 1996 e 85% (FSH) e 83% (eCG) em 1997. Todos os valores citados para 1996 e 1997 foram significativamente diferentes (P<0,05). Em 1997 foram utilizados dois produtos novos, Pluset e Folltropin-V, ambos à base de pFSH. Com Pluset o número de óvulos e embriões degenerados lavados foi de 11,7 unidades/doadora e o número de embriões de boa qualidade foi de 5,1 unidades/lavagem. Com Folltropin-V, o número foi de 4,5 unidades/lavagem e o de embriões viáveis de 1,9 unidades/doadora. Melhores resultados foram obtidos com Pluset (P<0,05).
Bovino; Holandês; estresse de calor; aclimatação; embrião; superovulação
The results of a multiple ovulation and embryo transfer (MOET) program, obtained in its first two years, were evaluated. The donor heifers were imported already pregnant from Hungary to the North-East area of Brazil. Comparisons of the results obtained in 1996 and 1997 with superovulation programs with AUSA-Superov (FSH) and Folligon (eCG) were made. The number of CL found on both ovaries was 5.3 (FSH) and 4.2 (eCG) in 1996, and 8.2 (FSH) and 9.6 (eCG) in 1997, while the number of degenerated eggs and embryos was 2.8 (FSH) and 3.8 (eCG) in 1996, and 6.5 (FSH) and 7.4 (eCG) in 1997. The number of good quality embryos was 1.9 (FSH) and 2.3 (eCG) in 1996, and 5.5 (FSH) and 4.0 (eCG) in 1997. The embryo collection efficiency was 40% (FSH) and 35% (eCG) in 1996, and 85% (FSH) and 83% (eCG) in 1997. Significant differences were found for all results obtained in 1996 and 1997 (P<0.05). Two new commercial products were used in 1997, Pluset and Folltropin-V, both with FSHp. With Pluset therapy the number of unfertilized eggs and degenerated embryos harvested was 11.7 per retrieval, while the number of good quality embryos was 5.1 per retrieval. With Folltropin therapy the number was 4.5 and 1.9, respectively, showing that better results were obtained with Pluset (P<0.05).
Cattle; Holstein-Friesian; heat stress; acclimatization; embryo; superovulation
Efeito da superovulação sobre o desempenho de bovinos doadores de embrião importados de clima temperado para clima tropical nos dois primeiros anos de adaptação
[Effect of superovulation on performance of bovine embryo donors imported from temperate zone to tropical climate during the first two years of adaptation]
B. Bényei1, C.C.W. Barros2
1Instituto Experimental da Faculdade de Ciências Veterinárias, Universidade São Estevão,
2225, Üllõ-Budapeste, Hungria
Consultor do convênio entre CODEVASF e AGROINVEST (Hungria)
2Médico Veterinário - Distrito de Irrigação do Projeto Senador Nilo Coelho, Petrolina-PE
Recebido para publicação em 5 de fevereiro de 1999.
E-mail: benyeib@matavnet.hu
RESUMO
Avaliou-se o desempenho de fêmeas bovinas doadoras de embrião nos primeiros dois anos de trabalho em um centro do programa multiple ovulation and embryo transfer (MOET). Utilizaram-se novilhas prenhes de recente importação de clima moderado para o Nordeste Brasileiro. Compararam-se os resultados de superovulação em 1996, primeiro ano de adaptação dos animais, e 1997 com AUSA-Superov (FSH) e Folligon (eCG). O número de CL contados no ovário foi 3,3 (FSH) e 4,2 (eCG) em 1996 e 8,2 (FSH) e 9,6 (eCG) em 1997, o de óvulos e embriões degenerados coletados de 2,8 (FSH) e 3,8 (eCG) em 1996 e 6,5 (FSH) e 7,4 (eCG) em 1997, o de embriões de boa qualidade de 1,9 (FSH) e 2,3 (eCG) em 1996 e 5,5 (FSH) e 4,0 (eCG) em 1997. A eficiência da coleta foi de 40% (FSH) e 35% (eCG) em 1996 e 85% (FSH) e 83% (eCG) em 1997. Todos os valores citados para 1996 e 1997 foram significativamente diferentes (P<0,05). Em 1997 foram utilizados dois produtos novos, Pluset e Folltropin-V, ambos à base de pFSH. Com Pluset o número de óvulos e embriões degenerados lavados foi de 11,7 unidades/doadora e o número de embriões de boa qualidade foi de 5,1 unidades/lavagem. Com Folltropin-V, o número foi de 4,5 unidades/lavagem e o de embriões viáveis de 1,9 unidades/doadora. Melhores resultados foram obtidos com Pluset (P<0,05).
PALAVRA-CHAVES: Bovino, Holandês, estresse de calor, aclimatação, embrião, superovulação
ABSTRACT
The results of a multiple ovulation and embryo transfer (MOET) program, obtained in its first two years, were evaluated. The donor heifers were imported already pregnant from Hungary to the North-East area of Brazil. Comparisons of the results obtained in 1996 and 1997 with superovulation programs with AUSA-Superov (FSH) and Folligon (eCG) were made. The number of CL found on both ovaries was 5.3 (FSH) and 4.2 (eCG) in 1996, and 8.2 (FSH) and 9.6 (eCG) in 1997, while the number of degenerated eggs and embryos was 2.8 (FSH) and 3.8 (eCG) in 1996, and 6.5 (FSH) and 7.4 (eCG) in 1997. The number of good quality embryos was 1.9 (FSH) and 2.3 (eCG) in 1996, and 5.5 (FSH) and 4.0 (eCG) in 1997. The embryo collection efficiency was 40% (FSH) and 35% (eCG) in 1996, and 85% (FSH) and 83% (eCG) in 1997. Significant differences were found for all results obtained in 1996 and 1997 (P<0.05). Two new commercial products were used in 1997, Pluset and Folltropin-V, both with FSHp. With Pluset therapy the number of unfertilized eggs and degenerated embryos harvested was 11.7 per retrieval, while the number of good quality embryos was 5.1 per retrieval. With Folltropin therapy the number was 4.5 and 1.9, respectively, showing that better results were obtained with Pluset (P<0.05).
Keywords: Cattle, Holstein-Friesian, heat stress, acclimatization, embryo, superovulation
INTRODUÇÃO
O hormônio foliculoestimulante (FSH) tem função essencial no desenvolvimento dos folículos (Monneaux et al., 1983). O uso de FSH exógeno para induzir superovulação é baseado nessa função fisiológica. Folículos em vários estágios de desenvolvimento estão normalmente presentes nos ovários, em qualquer tempo. Grupos consecutivos de folículos pequenos crescem, maturam e se degeneram ou ovulam. O FSH estimula o crescimento de folículos pequenos. FSH exógeno reverte a atresia de folículos acima de 1,7mm (Moor et al., 1984). O hormônio luteinizante (LH) estimula a produção de andrógenos na teca interna. Andrógenos tecais são usados como precursores para a produção de estrógenos pelas células granulosas que foram estimuladas pelo FSH (Richards, 1980). Essa é uma sucessão essencial para recomeçar a meiose nos oocistos imediatamente antes da ovulação. O LH excessivo durante o tratamento de superovulação causa a ativação prematura dos oocistos (Moor et al., 1984). Em vacas (Donaldson & Ward, 1986a,b) superovuladas com FSH, a alta concentração de LH resultou em baixa fecundação. Entre os profissionais, as preparações com FSH são as mais preferidas para superovular doadoras por causa da baixa razão FSH:LH (Seidel & Seidel, 1991). O produto AUSA-Superov contém baixa concentração de LH, e a razão FSH:LH é maior que 500 (Donaldson & Ward, 1986a). O procedimento de purificação e controle de qualidade de Folltropin-V assegura uma constância de baixa proporção de FSH:LH (Armstrong & Opavsky, 1986; Lindsell et al., 1986). No Pluset a razão FSH/LH é elevada (1:1), mas pelos vários autores, o resultado de superovulação é melhor se comparado com os produtos de FSH com baixa concentração de LH (Lauria et al., 1983; Coulthard & Burton, 1991; Cremonensi et al., 1992; Boland, 1995).
A mudança das condições meteorológicas para as novilhas importadas de clima temperado para o clima tropical semi-árido provoca acentuado estresse. Elas devem adaptar-se à relativa continuidade das horas claras do dia, a não caracterização de quatro estações do ano e à temperatura elevada (Bényei et al., 1999)
Sobre as condições de ambiente mencionadas e seus efeitos de estresse de calor para a reprodução há vários trabalhos da literatura. Em vacas leiteiras em ambiente subtropical no verão (temperatura acima de 30ºC), o resultado da inseminação diminui significativamente, caindo até 20% na eficiência, em média. Essa tendência nas novilhas aparece quando a temperatura está acima de 35ºC (Badinga et al., 1985). Embora pareça que a habilidade da superovulação das vacas estressadas pela temperatura elevada não mude (Putney et al., 1988a), temperatura acima de 32ºC aumenta a quantidade de embriões das categorias 2 ou 3 e de embriões retardados (Putney et al., 1989). Outros autores observaram efeito do calor sobre o desenvolvimento dos embriões (Massey et al., 1984; Shea et al., 1984; Gordon et al., 1987) ou mesmo sobre a superovulação nas vacas puras.
Há poucas referências na literatura internacional sobre a aclimatação das vacas da raça Holandesa nascidas no clima temperado e transferidas para o ambiente semi-árido.
Este trabalho tem por objetivo estudar o efeito da aplicação de vários hormônios encontrados no mercado internacional sobre o desempenho de vacas doadoras de embriões, importadas de clima temperado para o semi-árido do nordeste brasileiro, em sua fase de adaptação às novas condições climáticas.
MATERIAL E MÉTODOS
As novilhas importadas foram colocadas em currais, em grupos de 10, utilizando-se 15 m2 /cabeça de área coberta com telhas de cerâmica e piso de asfalto recoberto por uma camada de bagaço de cana. A habilidade de absorção de água do bagaço, menor que a da palha de trigo utilizada na Hungria, foi a solução mais prática e mais próxima da tecnologia húngara. Com freqüência eram colocadas novas camadas até atingir saturação, quando então era trocada. Os animais tinham acesso a uma área contígua a céu aberto, com sombreamento para diminuir os efeitos dos raios solares, e piso de terra também coberto por bagaço de cana. Devido ao clima quente da região, as vacas eram refrescadas por um sistema de microaspersão que funcionava durante 10 minutos em cada hora. O conjunto de ordenha, com capacidade para cinco animais simultaneamente, era totalmente informatizado.
O volumoso, produzido na própria Fazenda, constava de feno de alfafa, capim-elefante, o mais utilizado na região, e milho, planta inteira desintegrada. Como concentrado utilizavam-se os farelos de milho, de trigo e de soja, e torta de algodão. A ração balanceada, distribuída antes da ordenha matinal e às 15h, obedecia os critérios de produção de leite, adotando-se três grupos de produção. O último grupo de alimentação era formado pelas vacas prenhes e vacas secas, que certamente foram doadoras lavadas uma ou mais vezes, e novilhas prenhes.
A Fazenda está localizada a 9º de latitude de Sul. A vegetação original, caatinga, sugere um ambiente seco e quente. Pelos registros de 30 anos, a quantidade média de chuva é de 430,4 mm/ano, com média das mínimas de 151,7 mm/ano e das máximas de 1055,6 mm/ano, concentrada (83%) entre dezembro e abril. No resto do ano praticamente não chove. A temperatura é alta. Na época de seca atinge o máximo de 42ºC continuamente e à noite não baixa de 27 a 28ºC; na época chuvosa, o máximo situa-se entre 33ºC e 38ºC, e à noite 18ºC a 22ºC. A umidade do ar é baixa, 35%, assim, as temperaturas elevadas são suportáveis. Baseado no estudo de Armstrong (1994), a região deve provocar estresse de leve a sério. As horas claras são diferentes das do local de origem dos animais. No semi-árido do Nordeste, a diferença entre o dia mais longo e o mais curto é apenas de 30 minutos. Na Hungria, o clima é continental, com quatro estações bem definidas no ano. A temperatura no verão chega a 35ºC e no inverno pode baixar até 20ºC. No verão são 16 horas claras e no inverno apenas 8 horas.
O trabalho genético com as vacas importadas constitui um programa MOET (Multiply Ovulation and Embryo Transfer). O rebanho de doadoras está ao lado da cidade de PetrolinaPE e os postos de transferência localizados nos estados da Bahia, Alagoas, Pernambuco, Sergipe e Minas Gerais (Vale do São Francisco). Para a escolha das receptoras, usaram-se critérios sanitários e reprodutivos, preferindo-se novilhas mestiças. Fez-se a indução do cio com prostaglandina (Veteglan, Serono Itália), e depois de 8 a 12 dias com base na presença de corpos lúteos florescentes, fez-se a superovulação, testando eCG, Folligon (Intervet Holanda) e três tipos de FSH, AUSA-Superov (Ausa International Inc. - Canadá), Folltropin-V (Vetrepharm Canadá) e Pluset (Serono Itália). Depois de 48 horas do início do tratamento aplicou-se prostaglandina para induzir o cio, e após 60 e 72 horas, fez-se a inseminação. A lavagem foi feita no sétimo dia após a primeira inseminação, usando DPBS (IMV França) (Seidel & Seidel, 1991). Os embriões de primeira qualidade foram tratados com 0,25% tripsina (Nutricell Brasil) por 60 a 90 segundos, e depois lavados três vezes em meio de manutenção de Emcare (Immuno-Chemical Products LTD - Nova Zelândia) (Thompson, 1996), preparados para congelamento com 1,5M etileno glicol em Emcare durante cinco minutos, e colocados no palhete de 0,25 (francês), pronto para transferência direta. Os palhetes foram colocados no congelador (SY-LAB, CRYOCELL - 1200 Áustria), à temperatura da câmara de 6ºC. Depois foi induzida a formação dos cristais (seeding) em 7ºC, e com a velocidade de 0.5ºC/minuto foram congelados até 30ºC e colocados no nitrogênio líquido. Dos embriões de segunda qualidade foram separados os melhores para congelar e os de qualidade mais baixa foram inovulados para receptoras sincronizadas.
Utilizou-se a análise de variância para estudar o efeito dos hormônios sobre o número de ovulações/doadora, número de estruturas coletadas, número de embriões viáveis coletados e número de ovos e embriões degenerados coletados. A diferença entre médias obtidas em 1996 e 1997 foram determinadas pelo teste t (P<0,05).
RESULTADOS E CONCLUSÃO
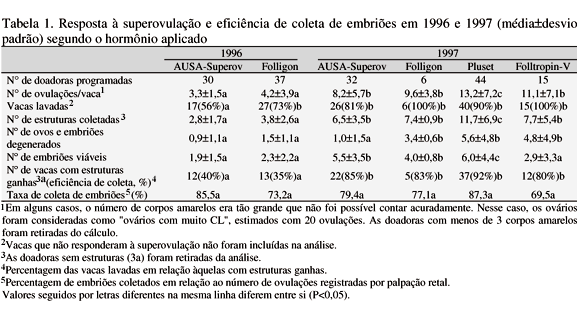
Os resultados de produção de embriões em 1996 e 1997 são apresentados na Tab. 1.
Em 1996, embora o número de vacas superovuladas tenha sido razoável, todos os dados mostraram resultados ruins O número de ovulações 3,3± 1,5 (FSH) e 4,2± 3,9 (eCG), o número de estruturas coletadas 2,8± 1,7 (FSH) e 3,8± 2,6 (eCG) e o número de embriões viáveis 1,9± 1,5 (FSH) e 2,3± 2,2 (eCG) foram baixos. Em 1997, esses valores foram significativamente melhores (P<0,05), isto é, número de ovulações igual a 8,2± 5,7 (FSH) e 9,6± 3,8 (eCG), número total de estruturas 6,5± 3,5 (FSH) e 7,4± 0,9 (eCG), e de embriões viáveis 5,5± 3,5 (FSH) e 4,0± 0,8 (eCG). O número de doadoras lavadas foi significativamente menor em 1996 (56% e 73% vs 81% e 100%; P<0,05) e a eficiência de coleta também foi menor (P<0,05), 40% (FSH) e 35% (eCG) vs 85% (FSH) e 83% (eCG). Os resultados indicam que em 1997 os hormônios utilizados foram efetivos, o que não ocorreu no ano anterior de modo satisfatório. A eficiência desses hormônios já foi citada na literatura (Holstein..., 1994). A razão de sua não eficiência no primeiro ano provavelmente está ligada com o novo ambiente das matrizes e as respostas à superovulação foram baixas naquele ano.
Segundo Dutt (1961) e Omtvedt (1989), o grande número de ovos infecundos é resultado do estresse materno em estudo feito em ovelhas e porcas, respectivamente, em câmara térmica isolada. Putney (1989) realizou o mesmo trabalho em temperatura ambiente em vacas e obteve resultados semelhantes. Todos esses autores acreditam que a temperatura elevada influi na maturação dos oocistos. Ealy et al. (1993) relataram uma pesquisa de laboratório, segundo a qual o embrião bovino é sensível, a partir da fecundação até o estádio de oito células, à temperatura de 43ºC, fato não observado nos estádios posteriores de desenvolvimento. O genoma bovino começa a funcionar no estádio de 8 a 16 células, e a partir desse, a célula produz uma proteína chamada proteína de estresse de temperatura (heat shock protein), que diminui o efeito letal da temperatura elevada (Barnes, 1990). Malayer (1988) explica que o aumento do número de óvulos e o grande número de embriões degenerados são provenientes da mudança de ambiente do útero.
A razão do insucesso durante o período de adaptação está provavelmente ligada ao estresse para a produção de embriões, devido à não caracterização das quatro estações do ano, às temperaturas elevadas e à continuidade das horas claras. Essas circunstâncias causaram a "dormência" do sistema reprodutivo do rebanho, com baixos resultados na produção de embriões no primeiro ano.
Como os trabalhos mencionados na literatura consultada mostram que o estresse de calor não interfere nos resultados de superovulação, mas altera a qualidade das estruturas obtidas, supõe-se, então, que os piores resultados de 1996 (número de corpos amarelos no ovário, percentagem de vacas lavadas, número de estruturas lavadas e embriões viáveis) são devidos à menor habilidade de superovulação. Esse período durou um ano e meio, atrasando o trabalho do programa de MOET.
O hormônio Pluset, usado em 1997, é um produto obtido da hipófise suína, e embora a razão FSH:LH seja 1:1, ele apresentou bons resultados. O número de ovulações por vaca foi significativamente maior que o dos outros hormônios utilizados (P<0,05) e o número total de embriões obtidos foi bom (11,7± 6,9). O número de vacas lavadas (90%) e a eficiência de superovulação (92%) foram melhores nesse tratamento do que nos outros, o que evidência a boa habilidade para superovulação. A desvantagem desse hormônio foi o aumento do número de embriões descartados. O AUSA-Superov, produto da hipófise suína, e cuja proporção FSH:LH é maior que 500, resultou em superovulação leve no segundo ano. Segundo Halve... (1994), é possível repetir a superovulação logo após o cio depois da lavagem caso os ovários se regenerem. O número de estruturas obtidas com AUSA-Superior foi significativamente menor que com Pluset (P<0,05), mas o número de embriões descartados foi baixo, desse modo, o número de embriões obtidos foi razoável. O Folltropin-V é preparado com extrato da hipófise de suíno, mas o efeito dele não trouxe o resultado desejado. O número de CL mostra a habilidade da superovulação, resultando em boa quantidade de estruturas lavadas, mas o número de embriões viáveis foi pequeno. O Folligon é um produto de eCG, com peso molecular grande, quase o peso de proteína, logo se produzem anticorpos contra ele, por isso foi usado com restrição (seis animais superovulados). O resultado da superovulação foi razoável mas o número de embriões viáveis foi baixo, sendo uma desvantagem, o que conduziu a um resultado total fraco, por isso foi pouco usado.
Em 1996 foram superovuladas 45 doadoras e efetuadas 68 lavagens. Em 1997 foram superovulados 46 animais e feitas 80 lavagens. Neste último ano pôde-se incluir mais 17 lavagens de novilhas, mas logo após a lavagem elas foram inseminadas, assim não se pôde contar com elas mais tarde. Isso resultou em 1,51 e 2,00 lavagens por doadora (P>0,05) em 1996 e 1997, respectivamente.
O limite de superovulação de uma doadora em um ano depende de vários fatores. O resultado do primeiro ano não indicou a possibilidade de um Centro de Doadoras (Bényei, 1992) porque no início do trabalho não havia informações sobre a habilidade de superovulação das doadoras, por isso foram programadas todas as vacas disponíveis, sendo esta a razão do grande número de animais superovulados. Depois da superovulação sem sucesso, elas foram inseminadas, o que explica o menor número de lavagens e resultados de superovulação abaixo das expectativas. A situação se normalizou em 1997, assim o rodízio dos processamentos foram equilibrados (inseminações para prenhez, parições e lavagens), resultando em duas lavagens por ano. Estas foram influenciadas pelo intervalo entre o parto e a superovulação, de 60 dias, que em vários casos foi mais longo, mas nunca inferior a 60 dias, pelo período ótimo entre as lavagens, de 50 a 60 dias, porque poucas vacas ciclaram normalmente depois da lavagem (dados não apresentados), pelas vacas que produziram poucos embriões, mas que foram inseminadas depois da segunda lavagem, e pelas doadoras que não puderam ser superovuladas no período de 90 dias, mas que foram inseminadas. Assim, o resultado de duas lavagens por ano ocorre por três motivos: superovulação que começou no ano posterior à parição, doadoras consideradas boas só usadas para produzir embriões, e vacas só inseminadas por não serem boas doadoras. Essa foi a rotina da Unidade Central de Doadoras em 1997, isto é, parições, lavagens e inseminações aconteceram continuamente.
A conclusão final é que ao se projetar um programa de MOET da raça Holandesa importada de clima temperado para região tropical deve-se contar com um período de um ano e meio de aclimatação hormonal. Nesse período, os animais não são capazes de produzir embriões em grande quantidade, o que só ocorre depois da fase de adaptação. Deve-se pensar também que uma vaca será lavada duas vezes por ano. A escolha do hormônio disponível também reflete nos resultados.
- ARMSTRONG, D.T., OPAVSKY, M.A. Biological characterization of a pituitary FSH preparation with reduced LH activity. Theriogenology, v.25, p.135, 1986. (Abstract).
- ARMSTRONG, D.V. Heat stress interactions with shade and cooling. J. Dairy Sci., v.77, p.2044-2050, 1994.
- BADINGA, L., COLLIER, R.J., THATCHER, W.W. et al. Effects of climatic and management factors on conception rate of dairy cattle in subtropical environment. J. Dairy Sci, v.68, p.78-85, 1985.
- BARNES, F.L., EYESTONE, W.H. Early cleavage and the maternal zygotic transition in bovine embryos. Theriogenology, v.33, p.141-149, 1990.
- BÉNYEI, B. Embryo transfer as a tool to develop the genetic pool of dairy cattle. In: INTERNATIONAL CONGRESS ABOUT THE APPLICATION AND UTILIZATION OF AGRICULTURAL SCIENTIFIC RESULTS IN DEVELOPING COUNTRIES. Gödöllő, Hungary: Tropical and Subtropical Department, University of Agriculture, 1992. p.32. (Abstract)
- BÉNYEI, B., FÁRI, M., BARROS, C.W.C. et al. Superovulatory response of continuously heat stressed heifers and cows in Brazil. Theriogenology, v.51, p.260, 1999. (Abstract)
- BOLAND, M.P. Progress report on superovulation and embryo production in cattle and sheep using Pluset. Serono Rep., p.25-30, 1995.
- COULTHARD, H., BURTON, M.J. The efficacy of Pluset (Serono) as a superovulation agent in cattle and the effect of timing of GnRH injection prior to insemination. REUNION AETE, 7, Cambridge, 1991. p.35. (Abstract).
- CREMONENSI, F., STACCHEZZINI, S., GEIGER, R. et al. Superovulation in Maremmana cows. REUNION AETE, 8, Lyon, 1992. p.13. (Abstract).
- DONALDSON, L.E., WARD, D.N. Effects of luteinising hormone on embryo production in superovulated cows. Vet. Rec., v.119, p.625-626, 1986b.
- DONALDSON, L.E., WARD, D.N. Use of porcine follicle stimulating hormone after chromatographic purification in superovular of cattle. Theriogenology, v.25, p.625-626, 1986a.
- DUTT, R.H. Critical period for early embryo mortality in ewes exposed to high ambient temperature. J. Anim. Sci., v.22, p.713-720, 1961.
- EALY, A.D., DROST, M. HANSEN, P.J.J. Developmental changes in embryonic resistance to adverse effects of maternal stress in cows. J. Dairy Sci., v.76, p.2899-2905, 1993.
- GORDON, I., BOLAND, M.P., McGOVERN, H. et al. Effect of season on superovulatory responses and embryo quality in Holstein cattle in Saudi Arabia. Theriogenology, v.29, p.231, 1981.
- HALVE the time between superovulation. Embryo Count, v.7, p.5, 1994.
- HOLSTEIN produce well. Embryo Count, v.7, p.4, 1994.
- LAURIA, A., OLIVA, O., GENAZZANI, A.R. et al. Superovulation of dairy and beef cows using porcine FSH with defined LH content. Theriogenology, v.20, p.675-682, 1983.
- LINDSELL, C.E., RAJKUMAR, A.W., MANNING, S.K. et al. Variability in FSH ratios among batches of commercially available gonadotropins. Theriogenology, v.25, p.167, 1986.
- MALAYER, J.R., HANSEN, P.J., BUHI, W.C. Effect of day of the estrus cycle, side of the reproductive tract and heat shock on in-vivo protein secretion by bovine endometrium. J. Reprod. Fert., v.84, p.567-577, 1988.
- MASSEY, J.M., ODEN, A.J. No seasonal effect on embryo donor performance in the Southwest region of the USA. Theriogenology, v.21, p.196-217, 1984.
- MONNEAUX, D., CHUPIN, D., SAUMONDE, J. Superovulatory responses of cattle. Theriogenology, v.19, p.55-61, 1983.
- MOOR, R.M., KRUIP, T.A.M., GREEN, D. Intraovarian control of folliculogenesis: limits to superovulation? Theriogenology, v.21, p.103-116, 1984.
- OMTVEDT, I.T., NELSON, R.E., EDMONDS, R.L. Influence of heat stress during early, mid and late pregnancy of gilts. J. Anim. Sci., v.32, p.312-321, 1967.
- PUTNEY, D.J., DROST, M., THATCHER, W.W. Influence of summer heat stress on pregnancy rates of lactating dairy cattle following embryo transfer or artificial insemination. Theriogenology, v.31, p.765-778, 1989a.
- PUTNEY, D.J., MULLINS, S., THATCHER, W.W. et al. Embryonic development in superovulated dairy cattle exposed to elevated ambient temperatures between the onset of estrus and insemination. Anim. Reprod. Sci., v.19, p.37-51, 1989b.
- PUTNEY, D.J., THATCHER, W.W., DROST, M. et al. Influence of environmental temperature on reproductive performance of bovine embryo donors and recipients in the Southwest region of the United States. Theriogenology, v.30, p.905-922, 1988.
- RICHARDS, J.S. Maturation of ovarian follicles: actions and interactions of pituitary and ovarian hormones on follicular cell differentiation. Physiol. Rev., v.60, p.51-59, 1980.
- SEIDEL, G.E., SEIDEL, S.M. Training manual for embryo transfer in cattle. Rome: FAO Animal Production and Health Paper, 1991.
- SHEA, B.F., JANZEN, R.E., McDERMAND, D.P. Seasonal variation in response to stimulation and related embryo transfer procedures in Alberta over a nine year period. Theriogenology, v.21, p.186-195, 1984.
- THOMPSON, J. Defining the requirements for bovine embryo culture. Theriogenology, v.45, p.27-40, 1996.
Datas de Publicação
-
Publicação nesta coleção
21 Set 2000 -
Data do Fascículo
Ago 2000
Histórico
-
Recebido
05 Fev 1999