Resumos
Em delineamento inteiramente ao acaso, estudou-se o efeito da defaunação em ovinos que receberam cana-de-açúcar e uréia sobre o consumo e digestibilidade aparente de nutrientes, dinâmica da fase sólida, parâmetros de fermentação ruminal e de degradação in situ. As taxas de passagem no rúmen foram, respectivamente, de 3,05 e 1,97%/h, para ovinos faunados e defaunados. Valores mais elevados para taxa (5,4%/h) e extensão (46,7%) da digestão ruminal e de degradabilidade efetiva da FDN da cana (31,5%) foram obtidos nos ovinos faunados. Seus consumos diários (57,7 e 32,3 g/kg0,75, respectivamente para MS e FDN) foram superiores (P<0,06) aos dos ovinos defaunados. Em função do tempo de amostragem observaram-se maiores (P<0,05) concentrações de AGV total, acetato e propionato para os cordeiros faunados. Nos defaunados ocorreu queda pós-prandial mais acentuada (P<0,04) no pH. Para digestibilidade aparente diferença (P<0,05) entre tratamentos foi verificada apenas para matéria seca.
Rúmen; protozoários ciliados; defaunação; degradabilidade in situ; digestibilidade aparente; taxa de passagem
The defaunation effect on feed intake, nutrient apparent digestibilities, solid phase dynamics, ruminal fermentation parameters and in situ degradation in sheep fed on sugar cane plus urea was studied in a completely randomized design with three replicates. Ruminal passage rates of 3.05 and 1.97%/h were observed for the faunated and defaunated groups, respectively. Higher values for sugar cane NDF ruminal digestion rate (5.41%/h), extension (46.7%) and effective degradability (31.53%) were observed for the faunated group. The faunated group daily intake (57.7 and 32.3 g/kg0.75 for dry matter and NDF residue, respectively) was higher (P<0.06) than that for the defaunated group. Total VFA, acetate and propionate concentrations were higher (P<0.05) in the faunated group and were dependent on sampling time. Defaunated sheep showed higher (P<0.04) post prandial decrease in pH. Relative to apparent digestibility difference (P<0.05) between treatments was observed only for dry matter.
Rumen; ciliate protozoa; defaunation; digestibility, in situ degradability; rate of passage
Efeitos da defaunação em ovinos alimentados com cana-de-açúcar (Saccharum officinarum, L.) adicionada de uréia
[Effects of defaunation in sheep fed sugar cane (Saccharum officinarum, L.) plus urea]
F.C.F. Lopes1, L.J.M. Aroeira1, P.B. Arcuri1, M.S. Dayrell2, A. Vittori3
1Embrapa Gado de Leite
Rua Eugênio do Nascimento, 610 - Bairro Dom Bosco
36038-330 - Juiz de Fora, MG
2Nutriplan Prod. Agrop. Ltda - Juiz de Fora, MG
3Doutoranda em Zootecnia na Universidade Federal de Viçosa
4Zootecnista
Recebido para publicação em 1 de junho de 2001
Recebido para publicação após modificações em 15 de março de 2002.
E-mail: fcfl@cnpgl.embrapa.br
RESUMO
Em delineamento inteiramente ao acaso, estudou-se o efeito da defaunação em ovinos que receberam cana-de-açúcar e uréia sobre o consumo e digestibilidade aparente de nutrientes, dinâmica da fase sólida, parâmetros de fermentação ruminal e de degradação in situ. As taxas de passagem no rúmen foram, respectivamente, de 3,05 e 1,97%/h, para ovinos faunados e defaunados. Valores mais elevados para taxa (5,4%/h) e extensão (46,7%) da digestão ruminal e de degradabilidade efetiva da FDN da cana (31,5%) foram obtidos nos ovinos faunados. Seus consumos diários (57,7 e 32,3 g/kg0,75, respectivamente para MS e FDN) foram superiores (P<0,06) aos dos ovinos defaunados. Em função do tempo de amostragem observaram-se maiores (P<0,05) concentrações de AGV total, acetato e propionato para os cordeiros faunados. Nos defaunados ocorreu queda pós-prandial mais acentuada (P<0,04) no pH. Para digestibilidade aparente diferença (P<0,05) entre tratamentos foi verificada apenas para matéria seca.
Palavras-chave: Rúmen, protozoários ciliados, defaunação, degradabilidade in situ, digestibilidade aparente, taxa de passagem
ABSTRACT
The defaunation effect on feed intake, nutrient apparent digestibilities, solid phase dynamics, ruminal fermentation parameters and in situ degradation in sheep fed on sugar cane plus urea was studied in a completely randomized design with three replicates. Ruminal passage rates of 3.05 and 1.97%/h were observed for the faunated and defaunated groups, respectively. Higher values for sugar cane NDF ruminal digestion rate (5.41%/h), extension (46.7%) and effective degradability (31.53%) were observed for the faunated group. The faunated group daily intake (57.7 and 32.3 g/kg0.75 for dry matter and NDF residue, respectively) was higher (P<0.06) than that for the defaunated group. Total VFA, acetate and propionate concentrations were higher (P<0.05) in the faunated group and were dependent on sampling time. Defaunated sheep showed higher (P<0.04) post prandial decrease in pH. Relative to apparent digestibility difference (P<0.05) between treatments was observed only for dry matter.
Keywords: Rumen, ciliate protozoa, defaunation, digestibility, in situ degradability, rate of passage
INTRODUÇÃO
Ao constituir um ecossistema aberto e contínuo, o rúmen proporciona ambiente ideal para a manutenção de uma população microbiana estável, que tem evoluído no transcurso de milhões de anos de seleção natural (Yokoyama & Johnson, 1988). Sua microflora é bastante diversa, consistindo de cerca de 109 a 1010 bactérias, 105 a 106 protozoários e 103 a 104 fungos por ml de fluído ruminal (Santra & Jakhmola, 1998).
Calcula-se que os ciliados, que constituem a maioria dos protozoários presentes no rúmen, respondam por cerca de 2% do peso do conteúdo ruminal, 40% do nitrogênio microbiano e proporcionem 60% dos produtos da fermentação microbiana nesse órgão (Yokoyama & Johnson, 1988). A despeito desses números, o benefício de sua presença para o ruminante é controverso. Pressões evolucionárias não os têm eliminado e acredita-se que possam desempenhar papel importante, mas não ainda devidamente elucidado, na ecologia do rúmen (Ryle & Ørskov, 1987).
Alguns autores (Jouany et al., 1988; Santra & Jakhmola, 1998) têm estudado os efeitos da defaunação sobre o ecossistema ruminal. Além de aspectos inerentes à dieta, ao animal, manejo etc., têm-se observado decréscimos nas digestibilidades ruminal e total da matéria orgânica, da matéria seca e, principalmente, das frações fibrosas das dietas fornecidas aos animais defaunados, mostrando a importância dos protozoários na degradação ruminal da parede celular (Veira, 1986; Santra & Jakhmola, 1998).
Os objetivos do presente estudo foram avaliar o efeito da defaunação em ovinos com dieta exclusiva de cana-de-açúcar e uréia sobre o consumo voluntário e a digestibilidade aparente de nutrientes, a dinâmica da fase sólida e os parâmetros de fermentação ruminal e de degradação in situ.
MATERIAL E MÉTODOS
O experimento foi conduzido na Embrapa Gado de Leite (Campo Experimental de Coronel Pacheco), no município de Coronel Pacheco, Minas Gerais.
Foram usados seis ovinos machos castrados e caudectomizados, com idade entre 21 e 28 meses, peso vivo médio de 45,6kg, sem raça definida e previamente fistulados no rúmen. O delineamento estatístico adotado foi inteiramente ao acaso, comparando-se dois tratamentos: ovinos com fauna ruminal normal (faunados) ou com fauna ruminal desprovida de protozoários (defaunados).
Para defaunação artificial do rúmen, os cordeiros foram separados de suas mães 48 horas após o nascimento e prevenidos, por isolamento geográfico, do contato com ruminantes (Jouany et al., 1998). O status da defaunação foi acompanhado semanalmente.
Nos primeiros 60 dias, leite bovino integral previamente congelado foi fornecido aquecido a 40oC. A partir da primeira semana, tiveram à disposição mistura 1:1 (base matéria seca) de feno de coast-cross (Cynodon dactylon cv. Coast-cross) e de concentrado rico em lipídios (fubá de milho, 73,0%; farelo de soja, 22,0%; calcário, 1,4%; fosfato bicálcico, 1,3% e mistura mineral, 1,0%).
Para evitar toxidez por cobre (Dayrell et al., 1993) a mistura mineral dos animais defaunados foi desprovida desse elemento. Todos os alimentos foram mantidos em estufa de ventilação forçada (65oC, 48h) antes de estarem disponíveis aos cordeiros. Forneceu-se água deionizada e tratada com solução de hipoclorito de sódio aos animais defaunados. Após o oitavo mês de vida, os animais defaunados passaram a receber sacarina, alimento obtido da fermentação aeróbica da cana-de-açúcar picada.
Os cordeiros faunados foram criados segundo o manejo normalmente dispensado ao rebanho de ovinos da Embrapa Gado de Leite.
Durante o período experimental, os cordeiros foram mantidos em gaiolas metabólicas individuais e os tratamentos geograficamente isolados para evitar infestação por protozoários no rúmen dos animais defaunados.
A dieta experimental foi fornecida em uma refeição diária pela manhã, e constituiu-se exclusivamente de cana-de-açúcar picada (cultivar CB 47-355, brix médio de 21o) adicionada (base natural) de 1% da mistura 9:1 de uréia:sulfato de amônia. Todo o alimento foi previamente preparado e estocado (10oC) em sacos plásticos (6kg). O fornecimento da dieta experimental foi feito por substituição gradativa e respeitou-se o período de adaptação de 10 dias, findo o qual iniciou-se a fase de coleta de amostras.
Para obtenção dos coeficientes de digestibilidade da matéria seca (MS), da proteína bruta (PB), da fibra em detergente neutro (FDN) e da fibra em detergente ácido (FDA) foram feitas, por sete dias consecutivos, pesagens e amostragens da dieta oferecida e respectivas sobras individuais. A coleta total individual diária de fezes foi feita com auxílio de bolsas adaptadas aos animais. As amostras armazenadas a 10ºC foram descongeladas, pré-secas (65oC, 72h), moídas (1mm) e analisadas quanto aos teores de MS (105ºC), de PB, pelo método Kjeldahl, e de FDN e FDA, pelo método de Van Soest, segundo os procedimentos de Silva (1998). O cromo nas fezes foi dosado por espectofotometria de absorção atômica, após digestão com ácido nitro-perclórico (Kimura & Miller, 1952). Amostras diárias individuais de urina foram coletadas mas perdidas por problemas relacionados ao seu armazenamento.
Amostras de fluído ruminal foram coletadas imediatamente antes (tempo zero) e 2, 4, 6, 8, 12 e 24 horas após o fornecimento da refeição diária. Após coadura e homogeneização, determinou-se o pH (potenciômetro digital Micronal B-375). Imediatamente após, foram pipetadas duas alíquotas de 5ml de líquido ruminal, adicionadas a tubos contendo 1ml de ácido metafosfórico 25% para análises das concentrações dos principais ácidos graxos voláteis (AGV; acético, propiônico e butírico) pela cromatografia gasosa por ionização de chama. As amostras foram rigorosamente homogeneizadas e congeladas (10oC). Amostras de líquido ruminal para determinação de nitrogênio amoniacal também foram coletadas, sendo no entanto perdidas por problemas relacionados ao seu armazenamento.
Os dados de pH e de concentrações (mg/ml) e percentagens molares de AGV foram analisados pelo procedimento GLM (SAS..., 1985), segundo delineamento em parcelas subdivididas com 20 graus de liberdade (GL) para o erro, nos quais os tratamentos constituíram as parcelas principais e os tempos de amostragem, as subparcelas.
Para estimativa dos parâmetros de degradabilidade in situ (MS, FDN e FDA) sacos confeccionados em náilon (11x 5cm) contendo 2,5g de amostra seca (65oC) e moída (5mm) da dieta experimental foram incubados no rúmen (Nocek, 1988). Trabalhou-se com abertura de malhas de 46m, valor próximo ao recomendado por Veira (1986) como suficiente para permitir o fluxo e a atividade normal dos grandes protozoários no interior dos sacos de náilon. Antes da incubação todos os sacos foram mergulhados em água (30min). Os referentes ao tempo zero (estimativa da fração solúvel mais partículas com tamanho reduzido que atravessam os poros do náilon, S) foram retirados e congelados. Os demais foram colocados no rúmen e retirados 2, 6, 12, 24, 48 e 72h após a incubação, sendo também congelados. Todos os sacos foram lavados simultaneamente, secos (65°C, 72 h) e pesados, e nos resíduos, analisados quanto aos teores de MS (105°C), FDN e FDA.
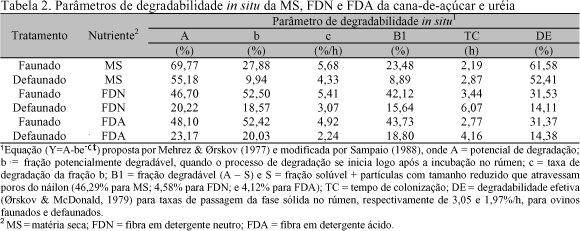
Os parâmetros de degradabilidade in situ foram estimados pelo processo iterativo do algoritmo Marquardt com auxílio do procedimento para modelos não lineares (PROC NLIN) do SAS... (1985), segundo a equação proposta por Mehrez & Ørskov (1977) e modificada por Sampaio (1988). Os parâmetros da equação foram estimados por tratamento e os tempos de colonização (TC) foram calculados à semelhança de Figueira (1991). Foram feitas novas estimativas dos parâmetros da equação sem os dados referentes aos tempos de incubação inferiores ao TC calculado (GL do erro do modelo = 11). As determinações da degradabilidade efetiva (DE) foram feitas segundo Ørskov & McDonald (1979).
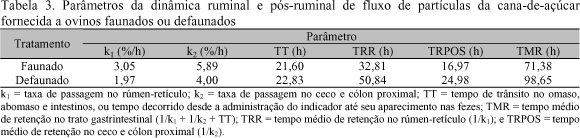
Para estimativa da taxa de passagem da fase sólida no rúmen (k1), a cana-de-açúcar picada foi lavada com detergente neutro comercial. O material seco obtido foi tratado com dicromato de sódio (Figueira, 1991), e 20g de fibra mordentada (5,24% de cromo v/v) foram colocadas no rúmen de cada cordeiro. Coletaram-se amostras de fezes com auxílio de bolsas em intervalos de 4h até 72h e, após esse tempo, em intervalos de 6h até as 120h da administração do material mordentado. Após as coletas as amostras de fezes foram congeladas (10oC) e, posteriormente, pré-secas (65oC, 72h), moídas (1mm) e analisadas quanto aos teores de cromo.
As estimativas das taxas de passagem da fase sólida foram feitas pelo processo iterativo do algoritmo Marquardt com auxílio do procedimento PROC NLIN (SAS..., 1985), segundo a equação usada por Grovum & Williams (1973). Os parâmetros do modelo foram obtidos por tratamento (GL do erro do modelo = 73).
RESULTADOS E DISCUSSÃO
A composição química média da cana-de-açúcar adicionada de 1% da mistura 9:1 de uréia:sulfato de amônia foi de 24,8, 12,6, 56,4 e 34,9%, respectivamente, para MS, PB, FDN, e FDA expressos com base na MS. Todos os valores situam-se nos limites dos relatados por Oliveira (1990) e Aroeira et al. (1993a, 1995) e podem ser considerados bastante representativos da dieta.
Na Tab. 1 têm-se os valores médios observados para consumo diário (g/kg0,75) e para os coeficientes de digestibilidade aparente dos nutrientes (MS, PB, FDN e FDA) da dieta estudada.
O baixo consumo de MS observado pode ser considerado característico para animais alimentados exclusivamente com cana-de-açúcar e uréia. Em trabalho com novilhos que receberam a mesma dieta, Oliveira (1990) relatou ingestão de 41,72 g/kg0,75/dia. Quando suplementada com concentrados protéicos, obtiveram-se valores mais elevados (70,04 a 86,6 g/kg0,75/dia; Figueira, 1991; Aroeira et al., 1993b), destacando-se a importância dessa estratégia nutricional para a dieta em questão.
O baixo nível de consumo foi atribuído, principalmente, ao elevado conteúdo de fibra indigestível (100A; Tab. 2) na cana-de-açúcar (Figueira, 1991). O acúmulo dessa fração e o prolongamento de seu tempo de permanência no rúmen provocou reduzida taxa de passagem nesse compartimento, impondo restrições à ingestão da dieta, particularmente severas nos ovinos defaunados.
Quanto à digestibilidade da MS, a despeito da análise de variância para esse dado ter sido feita com apenas quatro GL para o erro, diferenças (P<0,05) foram verificadas entre tratamentos (Tab. 1). Para PB os valores foram semelhantes (P>0,05), embora mensurações dessa característica sejam de limitado valor em estudos de nutrição de ruminantes (Veira, 1986). Outras considerações sobre o metabolismo do nitrogênio e da nutrição protéica dos ovinos não puderam ser feitas face às relatadas perdas de amostras. Os coeficientes de digestibilidade aparente da MS e PB foram próximos aos valores relatados para novilhos por Figueira (1991) e Aroeira et al. (1993b), e os referentes à FDN e FDA superiores aos observados pelos mesmos autores.
Os parâmetros de degradabilidade in situ da MS, FDN e FDA da dieta experimental encontram-se na Tab. 2 e os parâmetros da dinâmica ruminal e pós-ruminal de fluxo de partículas da cana-de-açúcar, de acordo com os tratamentos, são apresentados na Tab. 3.
Os parâmetros da dinâmica ruminal e pós-ruminal da cana-de-açúcar nos ovinos faunados foram próximos aos obtidos em novilhos por Figueira (1991) e Aroeira et al. (1993a, 1995) e podem ser considerados típicos para essa forrageira. Os parâmetros de degradação in situ da MS e FDN observados para esse tratamento (Tab. 2) situaram-se nos limites dos relatados para bovinos e bubalinos que receberam dietas à base de cana-de-açúcar, exceto quanto às taxas de degradação que foram sempre mais elevadas do que aquelas consultadas na literatura (Figueira, 1991; Aroeira et al. 1993a; 1995; Franzolin & Franzolin, 2000), e semelhantes apenas ao valor de 6%/h verificado por Pereira et al. (1996) para taxa de degradação da MS em bovinos em dieta exclusiva de cana e uréia.
Com base nos parâmetros das Tab. 2 e 3 percebe-se que os valores são sempre mais favoráveis aos ovinos faunados. Isso parcialmente justifica as diferenças (P<0,06) detectadas no consumo de nutrientes (Tab. 1), a despeito da análise de variância para estes dados ter sido feita com apenas quatro GL para o erro.
A literatura é controversa quanto ao efeito da defaunação sobre a taxa de passagem da fase sólida no rúmen (Jouany et al., 1988; Santra & Jakhmola, 1998). No experimento a taxa de passagem observada para os ovinos defaunados foi bem menor do que a dos animais faunados. A despeito do sensível aumento no tempo de retenção ruminal da digesta, os valores de degradabilidade efetiva das frações fibrosas permaneceram extremamente baixos para os cordeiros defaunados, indicando limitada atividade celulolítica ruminal.
Entretanto, em função do maior tempo de retenção no ceco e cólon proximal, devido à menor taxa de passagem nesses compartimentos, consistente digestão pós-ruminal das frações fibrosas potencialmente degradáveis não fermentadas no rúmen seria esperada nos animais defaunados face à semelhança (P>0,05) entre tratamentos nos coeficientes de digestibilidade aparente de FDN e FDA. Veira (1986) sugeriu que mudanças podem ocorrer no sítio de digestão, com aumento na digestão pós-ruminal de carboidratos em animais defaunados. Contudo, não houve diferenças (P>0,05) entre tratamentos quanto às concentrações e quantidades (g/kg0,75/dia) fecais de PB (dados não mostrados), que poderiam ser indício da existência de maior atividade celulolítica pós-ruminal nos animais defaunados.
Trabalhos com ruminantes que receberam dietas distintas evidenciaram o importante papel dos protozoários na digestão ruminal da MS, matéria orgânica e dos componentes da parede celular dos alimentos (De Smet et al., 1992; Santra & Jakhmola, 1998). Segundo Orpin (1984), a digestão da lamela média por enzimas pectinolíticas extracelulares dos holotríquios facilitaria o acesso de outros microrganismos celulolíticos, como bactérias e ciliados do gênero Epidinium, que engolfam fibras e digerem celulose, hemicelulose e pectina, acelerando o processo de fermentação das frações fibrosas.
Quanto aos tempos de colonização (TC), a defaunação provocou inflação em seus valores, principalmente nos referentes às frações fibrosas (Tab. 2). Figueira (1991) discutiu que o elevado TC observado para FDN da cana-de-açúcar pode estar relacionado ao alto conteúdo de carboidratos rapidamente solúveis na forrageira, os quais são preferencialmente fermentados pelos microrganismos ruminais. Neste experimento a ausência de protozoários ciliados parece ter agravado tal efeito e pode estar ainda associada à alterações qualitativas na população de bactérias (Santra & Jakhmola, 1998), atribuídas às variações no ambiente ruminal, provocadas pela maior disponibilidade de carboidratos solúveis nos tempos logo após as refeições.
Face à correlação entre TC e taxa de degradação (Ferri et al., 1998), valores menores para essa última característica seriam esperados para os animais defaunados. No entanto, a magnitude das diferenças foi tal que uma parcial responsabilidade pelo menor consumo de nutrientes dos ovinos defaunados pode ser creditada ao baixo valor da taxa de degradação da FDN observado neste tratamento. Segundo Ørskov & DeB Hovell (1978), uma das principais limitações da cana-de-açúcar refere-se às reduzidas taxas de degradação no rúmen, o que se explica pela alta correlação dessa característica com o consumo de MS (Ferri et al., 1998). Outros autores têm relatado diferenças nos valores das taxas de degradação em favor de animais faunados (De Smet et al., 1992), comprovando que a digestão das fibras vegetais é mais rápida na presença dos protozoários ciliados (Orpin, 1984).
Segundo Ryle & Ørskov (1987), o consumo de dietas à base de cana-de-açúcar exige prolongado tempo dedicado à mastigação e ruminação, promovendo abundante salivação, que resulta em valores de pH normalmente altos e razoavelmente constantes, favorecendo a sobrevivência de muitas espécies de protozoários. Assim, o ecossistema ruminal de bovinos que receberam dietas à base de cana-de-açúcar apresenta variada população de ciliados, com nítida predominância de pequenos protozoários do gênero Entodinium, além de restrito número de Epidinium e variável população de grandes protozoários da subclasse Holotrichia, especificamente dos gêneros Isotricha e Dasytricha (D'Agosto et al., 1996; Franzolin & Franzolin, 2000). Estes últimos, a despeito de seu reduzido número mas em função de seu tamanho, respondem pela maior parcela da biomassa de ciliados do rúmen de bovinos sob tal dieta (Minor et al., 1977), contribuindo indiretamente para a digestão da celulose (Ryle & Ørskov, 1987).
A partir da ingestão do alimento, partículas danificadas dos tecidos das plantas liberam conteúdo celular (açúcares solúveis) que atrai protozoários holotríquios por quimiotaxia. Em 5min, 90% dos ciliados holotríquios tornam-se associados a essas partículas danificadas (Orpin, 1984), metabolizando carboidratos solúveis, particularmente sacarose (Bonhomme, 1990), até sua exaustão do meio. Esses açúcares são armazenados intracelularmente como polímeros de reserva e, posteriormente, fermentados de um modo mais lento para produzir AGV. Por esse mecanismo tamponante, a maior parte dos carboidratos rapidamente fermentáveis é excluída da ação bacteriana, modulando e prolongando o período de fermentação ruminal (Bonhomme, 1990), evitando quedas pós-prandiais bruscas no pH, e favorecendo o contínuo desenvolvimento de microrganismos celulolíticos (Santra & Jakhmola, 1998).
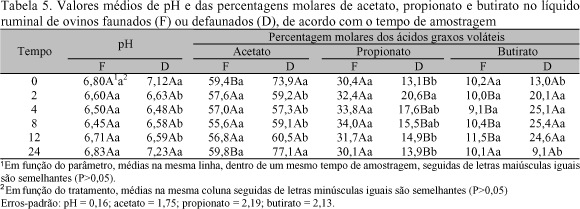
Nas Tab. 4 e 5 têm-se, de acordo com o tempo de amostragem, os valores de pH e das concentrações (mg/ml) e percentagens molares dos principais AGV no rúmen dos ovinos faunados e defaunados.
Não houve diferenças (P>0,05) entre tratamentos quanto aos valores de pH, os quais mantiveram-se próximos à neutralidade, independente do tempo de amostragem, o que pode ser considerado normal para dieta à base de cana-de-açúcar (Minor et al., 1977, Ryle & Ørskov, 1987). Entretanto, com relação ao tempo zero, queda pós-prandial (2h) ligeiramente mais acentuada (P<0,04) foi observada no pH ruminal dos ovinos defaunados, possivelmente pela ausência da atividade tamponante dos ciliados holotríquios (Santra & Jakhmola, 1998). Isso pode ser melhor observado com as concentrações totais de AGV (P<0,07), propionato (P<0,07) e acetato imediatamente antes (tempo zero) e após duas horas do fornecimento da ração diária. Nesses tempos ocorreram os picos de produção, evidenciando intensa atividade fermentativa no rúmen dos animais defaunados. Com a exaustão dos carboidratos solúveis (sacarose) nos tempos anteriores à refeição principal (0 e 24h), verificaram-se aumento (P<0,0001) na proporção de acetato e quedas nas de butirato (P<0,03) e propionato (Tab. 5). Dessas últimas observações percebe-se instabilidade da função ruminal dos cordeiros defaunados.
Nos ovinos faunados verificou-se, independente do tempo de amostragem, considerável estabilidade nos padrões de fermentação ruminal, com níveis praticamente constantes (P>0,05) nas concentrações e proporções de AGV, com destaque para a alta fermentação de propionato e baixo nível de butirato (Tab. 4 e 5), conforme relatado por Minor et al. (1977). As percentagens molares de acetato, propionato e butirato observadas nos ovinos faunados (Tab. 5) foram próximas às apresentadas por esses autores para dietas à base de cana-de-açúcar e uréia. No entanto, foram diferentes (P<0,05) das obtidas para os animais defaunados, especialmente com relação às proporções de propionato e butirato (Tab. 5), indicando mudanças nos padrões de fermentação da dieta consumida. Diferenças (P<0,05) entre tratamentos em função do tempo de amostragem também foram verificadas para as concentrações de AGV total, acetato e propionato (Tab. 4), em geral mais elevadas nos ovinos faunados.
Estes últimos resultados são parcialmente atribuídos às diferenças entre tratamentos quanto à ingestão de nutrientes (P<0,06), e por conseqüência, de consumo de carboidratos solúveis, que se constituem na principal fonte de energia para crescimento microbiano no rúmen de animais sob dietas à base de cana-de-açúcar.
Os dados observados de concentrações totais de AGV nos ovinos faunados foram, em geral, reflexo dos maiores valores de degradabilidade efetiva da MS e das frações fibrosas do alimento observados para os animais desse tratamento (Tab. 2).
CONCLUSÕES
Os ciliados desempenham importante papel ecológico no rúmen de ovinos sob dieta exclusiva de cana e uréia, orientando e controlando a intensidade da fermentação nesse órgão. A presença dos protozoários no rúmen permitiu a manutenção de ambiente estável e adequado à digestão in situ dos componentes da parede celular da cana-de-açúcar, aumentando a taxa de passagem e auxiliando na redução do efeito da FDN sobre a repleção ruminal. Isso permitiu aos ovinos faunados maior consumo de nutrientes e, por conseguinte, de carboidratos solúveis, que se constituem na principal fonte de energia para crescimento microbiano no rúmen de animais em dietas à base de cana-de-açúcar.
- AROEIRA, L.J.M.; LOPES, F.C.F.; DAYRELL, M.S. Digestibilidade, degradabilidade e taxa de passagem da cana-de-açúcar mais uréia e do farelo de algodăo em vacas mestiças Holandęs x Zebu em lactaçăo. Rev. Soc. Bras. Zootec., v.24, p.1016-1026, 1995.
- AROEIRA, L.J.M.; SILVEIRA, M.I.; LIZIEIRE, R.S. et al. Degradabilidade no rúmen e taxa de passagem da cana-de-açúcar mais uréia, do farelo de algodăo e do farelo de arroz em novilhos mestiços Europeu x Zebu. Rev. Soc. Bras. Zootec., v.22, p.552-564, 1993a.
- AROEIRA, L.J.M.; SILVEIRA, M.I.; LIZIEIRE, R.S. et al. Digestibilidade, balanço de nitrogęnio e concentraçăo de amônia no rúmen de novilhos mestiços alimentados com cana-de-açúcar de uréia mais farelos de arroz ou de algodăo. Rev. Soc. Bras. Zootec., v.22, p.893-901, 1993b.
- BONHOMME, A. Rumen ciliates: their metabolism and relationships with bacteria and their hosts. Anim. Feed Sci. Technol., v.30, p.203-266, 1990.
- D'AGOSTO, M.; CARNEIRO, M.E.; NETTO, C.M.M. et al. Avaliaçăo dos ciliados do rúmen de bovinos mantidos com duas dietas. Arq. Bras. Med. Vet. Zootec., v.48, p.353-361, 1996.
- DAYRELL, M.S.; ARCURI, P.B.; AROEIRA, L.J.M. et al. Toxidez de cobre em carneiros sem protozoários no rúmen. Arq. Bras. Med. Vet. Zootec., v.45, p.505-509, 1993.
- DE SMET, S.; DEMEYER, D.I.; VAN NEVEL, C.J. Effect of defaunation and hay:concentrate ratio on fermentation, fibre digestion and passage in the rumen of sheep. Anim. Feed Sci. Technol., v.37, p.333-344, 1992.
- FERRI, C.M.; JOUVE, V.V.; STRITZLER, N.P. et al. Estimation of intake and apparent digestibility of kleingrass from in situ parameters measured in sheep. Anim. Sci., v.67, p.535-540, 1998.
- FIGUEIRA, D.G. Efeito do nível de uréia sobre as digestibilidades aparente e "in situ" e a dinâmica da fase sólida em bovinos alimentados com cana-de-açúcar e farelo de algodăo. 1991. 123f. Dissertaçăo (Mestrado). Universidade Federal de Minas Gerais, Belo Horizonte, MG.
- FRANZOLIN, R.; FRANZOLIN, M.H.T. Populaçăo protozoários ciliados e degradabilidade ruminal em búfalos e bovinos zebuínos sob dieta ŕ base de cana-de-açúcar. Rev. Bras. Zootec., v.29, p.1853-1861, 2000.
- GROVUM, W.L.; WILLIAMS, V.J. Rate of passage of digesta in sheep. 4. Passage of marker through the alimentary tract and the biological relevance of rate-constants derived from the changes in concentration of marker in faeces. Br. J. Nut., v.30, p.313-329, 1973.
- JOUANY, J.P.; DEMEYER, D.I.; GRAIN, J. Effect of defaunating the rumen. Anim. Feed Sci. Technol., v.21, p.229-265, 1988.
- KIMURA, F.; MILLER, V.L. Chromic oxide measurement. Improved determination of chromic oxide in cow feed and feces. Agric. Food Chem., v.111, p.633-635, 1952.
- MEHREZ, A.Z.; ORSKOV, E.R. A study of the artificial fibre bag technique for determining the digestibility of feeds in the rumen. J. Agric. Sci., v.88, p.645-650, 1977.
- MINOR, S.; MacLEOD, N.A.; PRESTON, T.R. et al. Studies on digestion in different sections of the intestinal tract of bulls fed sugar cane/urea with different supplements. Trop. Anim. Prod., v.2, p.163-174, 1977.
- NOCEK, J.E. In situ and other methods to estimate ruminal protein and energy digestibility. A review. J. Dairy Sci., v.71, p.2051-2069, 1988.
- OLIVEIRA, W.H. Digestibilidade aparente e partiçăo da digestăo da cana-de-açúcar adicionada de níveis crescentes de uréia. 1990. 77f. Dissertaçăo (Mestrado). Universidade Federal de Minas Gerais, Belo Horizonte, MG.
- ORPIN, C.G. The role of ciliate protozoa and fungi in the rumen digestion of plant cell walls. Anim. Feed Sci. Technol., v.10, p.121-143, 1984.
- ŘRSKOV, E.R.; DeB HOVELL, F.D. Digestion ruminal del heno (medida atraves de bolsas de dacron) en el ganado alimentado con cańa de azucar o heno de pangola. Prod. Anim. Trop., v.3, p.9-11, 1978.
- ŘRSKOV, E.R.; McDONALD, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci., v.92, p.499-503, 1979.
- PEREIRA, O.G.; VALADARES FILHO, S.C.; GARCIA, R. et al. Degradabilidade in vivo e in situ de nutrientes e eficięncia de síntese de proteína microbiana, em bovinos, alimentados com cana-de-açúcar sob diferentes formas. Rev. Soc. Bras. Zootec., v.25, p.763-777, 1996.
- RYLE, M.; ŘRSKOV, E.R. Rumen ciliates and tropical feeds. World Anim. Rev., v.64, p.21-30, 1987.
- SAMPAIO, I.B.M. Experimental designs and modelling techniques in the study of roughage degradation in rumen and growth of ruminant. 1988. 228f. Thesis, (Phylosophy Doctor) Reading, Reading, UK.
- SANTRA, A.; JAKHMOLA, R.C. Effect of defaunation on animal productivity. J. Appl. Anim. Res., v.14, p.103-116, 1998.
- SAS Institute Inc. SASâ User's Guide: Statistics, Version 5 Edition. Cary, NC: SAS Institute Inc., 1985. 956p.
- SILVA, D.J. Análise de alimentos. Viçosa: Imprensa Universitária da UFV, 1998. 166p.
- VEIRA, D.M. The role of ciliate protozoa in nutrition of the ruminant. J. Anim. Sci., v.63, p.1547-1560, 1986.
- YOKOYAMA, M.T.; JOHSON, K.A. Microbiologia del rumen e intéstino In: CHURCH, C.D. El Rumiante: fisiología digestiva e nutrición. Zaragoza: Editorial Acribia S.A., 1988. p.137-157.
Datas de Publicação
-
Publicação nesta coleção
17 Jul 2002 -
Data do Fascículo
Abr 2002
Histórico
-
Recebido
01 Jun 2001 -
Aceito
15 Mar 2002