Resumos
Descreveu-se a estrutura populacional da raça Nelore Mocho utilizando dados do registro genealógico de animais nascidos entre 1969 e 1998. O banco de dados foi separado em quatro períodos de 1979-1983, 1984-1988, 1989-1993 e 1994-1998. Calcularam-se as estatísticas descritivas do número de criadores, de reprodutores e de reprodutrizes. A endogamia total aumentou de 0,55% para 0,98%, a esperada sob acasalamento ao acaso aumentou de 0,11% para 0,56% e a endogamia devido à subdivisão populacional permaneceu constante, ao redor de 0,45%, indicando que a subdivisão genética da raça Nelore Mocho é próxima a zero e praticamente inexistente. O tamanho efetivo populacional foi estimado pelo coeficiente total de endogamia e permaneceu ao redor de 120. A inclusão de animais no livro genealógico aumentou de 4.630 para 13.907, entretanto, esta ação não foi suficiente para incrementar o tamanho efetivo. Baseado na probabilidade de origem do gene foram calculados o número efetivo de fundadores, de ancestrais e de genomas remanescentes. Esses parâmetros decresceram ao longo do período, atingindo valores de 144, 98 e 64. Todas as estimativas foram análogas na descrição da estrutura genética populacional e devem ser consideradas no futuro para o desenvolvimento de programas de seleção.
Bovino; Nelore Mocho; estrutura populacional; tamanho efetivo; intervalo de gerações; probabilidade de origem do gene
The population structure of the Polled Nelore breed using pedigree records from animals registered from 1969 to 1998 was described. The data file was divided in the following periods: 1979-1983, 1984-1988, 1989-1993 and 1994-1998. Descriptive statistics of the number of breeders, sires and dams were calculated. The total inbreeding increased from 0.55% to 0.98%, the inbreeding expected under random mating increased from 0.11% to 0.56%, and the inbreeding due to population subdivision was constantly around 0.45%, indicating that genetic subdivisions of the Polled Nelore breed is near to zero and quite irrelevant. The effective population size estimated from the total inbreeding was around 120. The upgrading of non-registered animals increased from 4,630 to 13,907 animals, however, this action was not enough to increase the effective population size. Based on probabilities of gene origin the effective numbers of founders, ancestors and remaining genomes were calculated. These parameters decreased over the period reaching values of 144, 98 and 64. All the estimates were analogous in the description of the population genetic structure and should be considered in developing future selection programs.
Cattle; Polled Nelore; population structure; effective population size; generation interval; gene origin probability
Estrutura populacional da raça Nelore Mocho
[Population structure of the Polled Nelore breed]
F.J.C. Faria1, A.E. Vercesi Filho1, F.E. Madalena2, L.A. Josahkian3
1Aluno de Doutorado em Ciência Animal, Escola de Veterinária da UFMG
Caixa Postal 567
30123-970 - Belo Horizonte, MG
2Escola de Veterinária da UFMG
3Associação Brasileira de Criadores de Zebu e Faculdade de Agronomia e Zootecnia de Uberaba
Recebido para publicação em 17 de julho de 2001
Recebido para publicação, após modificações, em 27 de agosto de 2002
E-mail: fariafjc@dedalus.lcc.ufmg.br
RESUMO
Descreveu-se a estrutura populacional da raça Nelore Mocho utilizando dados do registro genealógico de animais nascidos entre 1969 e 1998. O banco de dados foi separado em quatro períodos de 1979-1983, 1984-1988, 1989-1993 e 1994-1998. Calcularam-se as estatísticas descritivas do número de criadores, de reprodutores e de reprodutrizes. A endogamia total aumentou de 0,55% para 0,98%, a esperada sob acasalamento ao acaso aumentou de 0,11% para 0,56% e a endogamia devido à subdivisão populacional permaneceu constante, ao redor de 0,45%, indicando que a subdivisão genética da raça Nelore Mocho é próxima a zero e praticamente inexistente. O tamanho efetivo populacional foi estimado pelo coeficiente total de endogamia e permaneceu ao redor de 120. A inclusão de animais no livro genealógico aumentou de 4.630 para 13.907, entretanto, esta ação não foi suficiente para incrementar o tamanho efetivo. Baseado na probabilidade de origem do gene foram calculados o número efetivo de fundadores, de ancestrais e de genomas remanescentes. Esses parâmetros decresceram ao longo do período, atingindo valores de 144, 98 e 64. Todas as estimativas foram análogas na descrição da estrutura genética populacional e devem ser consideradas no futuro para o desenvolvimento de programas de seleção.
Palavras-chave: Bovino, Nelore Mocho, estrutura populacional, tamanho efetivo, intervalo de gerações, probabilidade de origem do gene
ABSTRACT
The population structure of the Polled Nelore breed using pedigree records from animals registered from 1969 to 1998 was described. The data file was divided in the following periods: 1979-1983, 1984-1988, 1989-1993 and 1994-1998. Descriptive statistics of the number of breeders, sires and dams were calculated. The total inbreeding increased from 0.55% to 0.98%, the inbreeding expected under random mating increased from 0.11% to 0.56%, and the inbreeding due to population subdivision was constantly around 0.45%, indicating that genetic subdivisions of the Polled Nelore breed is near to zero and quite irrelevant. The effective population size estimated from the total inbreeding was around 120. The upgrading of non-registered animals increased from 4,630 to 13,907 animals, however, this action was not enough to increase the effective population size. Based on probabilities of gene origin the effective numbers of founders, ancestors and remaining genomes were calculated. These parameters decreased over the period reaching values of 144, 98 and 64. All the estimates were analogous in the description of the population genetic structure and should be considered in developing future selection programs.
Keywords: Cattle, Polled Nelore, population structure, effective population size, generation interval, gene origin probability
INTRODUÇÃO
A raça Nelore é conhecida na Índia como Ongole, e no seu país de origem é utilizada para trabalho e produção de leite. No Brasil, o rebanho Nelore é quase que exclusivamente utilizado para produção de carne (Santiago, 1986). Das raças indianas, a Nelore contou durante muitos anos com número reduzido de animais, porém, nos últimos decênios ela se expandiu, surgindo variedades entre elas a Mocha. Seu registro genealógico iniciou-se em 1969. De acordo com a Evolução... (2001), em 1993 foram vendidas 180.892 doses de sêmen da raça Nelore Mocho, o que representou 5,4% do total de doses vendidas das raças de corte, e 209.646 doses em 2000, equivalentes a 6,1% do total comercializado para corte. O Nelore Mocho é a segunda raça em número de registros junto à Estatísticas (2001).
No modelo de população ideal descrito por Wright (1931) as alterações de freqüência alélica seriam inexistentes, entretanto, as populações reais não seguem esse preceito, e vários são os fatores que comprometem esse modelo. As populações reais, selvagens ou domésticas, detêm certa estrutura populacional. Em programas de melhoramento animal, com o passar dos anos a estrutura inicial apresenta alterações na freqüência gênica. A existência de uma estrutura piramidal, com fluxo gênico preponderantemente unidirecional, favorece a diminuição das diferenças entre estratos. Em seguida às alterações estruturais ocorrem reduções da variabilidade genética (que pode ser quantificada pelo coeficiente de endogamia) e do número de linhagens e de genomas fundadores (quantificados por meio do número efetivo de fundadores e de genomas remanescentes).
Atenção tem sido dada para a exploração racional da variabilidade genética intra e inter populacional. Dessa forma, este trabalho objetivou descrever a estrutura populacional e quantificar a variabilidade genética da raça Nelore Mocho, por meio de informações obtidas de seu registro genealógico.
MATERIAL E MÉTODOS
Foram utilizados dados do registro genealógico da raça Nelore Mocho, disponíveis sob processamento eletrônico, da Associação Brasileira de Criadores de Zebu (ABCZ). O arquivo de dados consistiu de 360.035 observações entre os anos de 1969 a 1998, referentes a amostragem de 380.849 registros (Estatísticas, 2001). As informações referentes a cada animal consistiram de: pai, mãe, sexo, categoria de registro (puro de origem ou livro aberto), dia, mês e ano de nascimento, criador e proprietário. O arquivo de pedigree foi gerado por meio do banco de dados iniciais e totalizou 410.853 observações. As análises foram geradas por meio de procedimentos contidos no pacote estatístico SAS®.
Inicialmente foram geradas estatísticas descritivas sobre esta amostra para, em seguida, proceder-se à separação do banco de dados em períodos, compreendendo os anos de 1979-1983, 1984-1988, 1989-1993 e 1994-1998. Em cada período foram calculados o número de machos e fêmeas em reprodução, as médias, variâncias e covariâncias do número de progênies (filhos e filhas por reprodutor/matriz), e o intervalo de gerações médio. O intervalo de gerações ( ) em cada período foi calculado de acordo com a fórmula proposta por Rendel & Robertson (1951). Estimou-se também o intervalo médio de gerações de pais a filhos em todos os períodos como: pais de filhos, pais de filhas, mães de filhas e mães de filhos. O tamanho efetivo da população(Ne) foi estimado por meio da taxa de variação dos coeficientes médios de endogamia entre gerações sucessivas conforme Falconer & Mackay (1996).
) em cada período foi calculado de acordo com a fórmula proposta por Rendel & Robertson (1951). Estimou-se também o intervalo médio de gerações de pais a filhos em todos os períodos como: pais de filhos, pais de filhas, mães de filhas e mães de filhos. O tamanho efetivo da população(Ne) foi estimado por meio da taxa de variação dos coeficientes médios de endogamia entre gerações sucessivas conforme Falconer & Mackay (1996).
A estimação dos parâmetros baseados na probabilidade de origem do gene, juntamente com o coeficiente de endogamia, foram obtidos por meio da utilização do pacote PEDIG®, o qual apresenta vários programas escritos em linguagem Fortran, gentilmente cedido pelo Dr. Didier Boichard (INRA Station de Génétique Quantitative et Appliquée). O número efetivo de fundadores representa o número de animais com igual contribuição que produziria a mesma variabilidade genética encontrada na população estudada (Lacy, 1989). A aproximação clássica na determinação do número efetivo de fundadores foi obtida por:
Nfun =  , em que:
, em que:
Nfun = número efetivo de fundadores,
 = somatório da contribuição esperada (qk) do k fundador na população.
= somatório da contribuição esperada (qk) do k fundador na população.
O número efetivo de ancestrais representa o número mínimo de animais (fundadores ou não) necessário para se explicar a total diversidade genética da população estudada (Boichard et al., 1997). A determinação do número efetivo de ancestrais foi então determinada computando-se a contribuição marginal de cada ancestral por:
Nanc =  , em que:
, em que:
Nanc = número efetivo de ancestrais,
 = contribuição marginal do k ancestral na população.
= contribuição marginal do k ancestral na população.
O número efetivo de genomas remanescentes representa o número de fundadores com igual contribuição que não teria perda de alelos fundadores por deriva na progênie e que produziria a mesma diversidade genética da encontrada na população estudada (Boichard et al., 1997). O programa simulou segregações para cada progênie, gerando genótipos em função da amostragem aleatória de um alelo em cada progenitor. As freqüências fk foram determinadas por contagem na população referência:
 , em que:
, em que:
Ngen = número efetivo de genomas remanescentes,
Na = número efetivo de alelos (Crow & Kimura, 1970).
O Ngen foi determinado em função da média de 1.000 simulações de segregações gênicas. As estatísticas de F de Wright (1938), Fit, Fst e Fis, também foram calculadas. FIT é o coeficiente médio de endogamia da população, Fst designa o coeficiente médio de endogamia esperado se os reprodutores de cada período fossem acasalados aleatoriamente, e foi estimado em populações hipotéticas produzidas pelo acasalamento ao acaso dos pais dentro de cada período por meio do programa escrito em linguagem Fortran gentilmente cedido pelo Dr. Tetsuro Nomura (Kyoto Sangyo University), e Fis expressa o desvio da casualidade obtida nos acasalamentos atuais, obtido como:
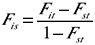
Se Fis>0, então a endogamia real ultrapassa o nível esperado sob acasalamentos ao acaso, implicando que os acasalamentos estão acontecendo entre animais mais aparentados do que a média, consequentemente a população estará dividida em sub-populações. Quando Fis<0, evita-se a endogamia ou predomina o acasalamento entre sub-populações.
RESULTADOS E DISCUSSÃO
Na avaliação de populações destaca-se a identificação do número de criadores e a distribuição do número de animais por rebanho. A Tab. 1 apresenta o número de criadores distribuídos em classes conforme o número de animais registrados, separados por sexo, no período de 1994 a 1998. A determinação desse período teve o intuito de retratar com maior fidelidade o número efetivo de criadores da raça. Constam da amostra registros de 132.267 animais, 66.566 (50,3%) machos e 65.701 (49,7%) fêmeas. Fato interessante foi o equilíbrio no número de machos e fêmeas registrados, com pequena superioridade numérica para o primeiro, contrariamente ao descrito por Faria et al. (2000) num período maior, no qual o número de fêmeas registradas superou o de machos. O maior número de registros de machos pode evidenciar aumento na procura por reprodutores, o que favoreceria o maior registro desse sexo, contrapondo às descrições para outras raças zebuínas estudadas, nas quais o registro de fêmeas foi superior ao de machos (Faria et al., 2002). O número médio de animais registrados por criador nesse período foi de 147, sendo 83 e 80 a média de machos e fêmeas registrados, respectivamente. Nesse qüinqüênio, 23,5% dos animais registrados originaram-se de 10 criadores, evidenciando concentração de animais, fato notadamente verificado em populações estruturadas e já descrito na literatura (Faria et al., 2001).
Do total de 157.637 machos, 6.663 animais foram identificados como pais do rebanho registrado, representando 4,2% do total de machos. As médias do número de filhos e filhas por progenitor foram 29,76 e 29,28, respectivamente, e 49,47 de crias de ambos os sexos. Dez animais (0,15% dos pais) contribuíram com 13,7% dos registros de filhos e um reprodutor foi progenitor de 4,0% dos animais registrados. Ao se estimar por período a média e a variância do número de progênies (Tab.2) observa-se aumento do número de reprodutores e da variância de tamanho de família. Isto certamente influenciou as estimativas populacionais posteriormente calculadas. A razão macho:fêmea passou de 1:13,2 para 1:20,5. A média de idade do pai quando do registro da primeira cria foi de 4,15 anos. Na Tab. 3 são expostos o número de reprodutores separados por classe de acordo com o sexo da progênie. Verifica-se que 5.181 reprodutores foram pais de machos e 5.990 pais de fêmeas, evidenciando maior seleção para pais de machos.
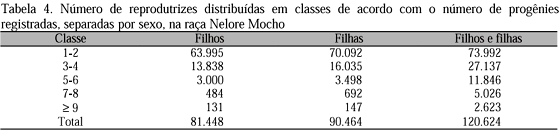
De 202.398 fêmeas, 120.630 foram identificadas como mães do rebanho registrado, representando 59% do total de fêmeas. Os números médios de filhos e filhas por mãe foram de 1,84 e 1,87, respectivamente, e o de ambos os sexos, 2,65. A distribuição do número de filhos por mãe, separados por sexo, pode ser visualizada na Tab. 4. Das mães, 67,5% contribuíram para o registro de machos e 75% para o de fêmeas, sugerindo que o maior rigor no registro de machos acrescentou maior intensidade de seleção para mães de machos do que para mães de fêmeas, já observado no caso dos progenitores.
As médias de idade das mães quando dos registros da primeira e da última crias foram de 4,6 e 7,9 anos, respectivamente, o que resultou em uma "vida útil" para fins de registro de 3,3 anos. A média do intervalo de registros de suas crias foi de 1,4 anos (tempo entre registros sucessivos). Salienta-se que a média de idade ao primeiro registro pode não corresponder à idade ao primeiro parto, assim como a média do intervalo de registros das crias pode não corresponder à média do intervalo de partos pelo fato de algumas crias não terem sido registradas.
Dada a importância e a necessidade de descrição acurada das populações estruturadas, determinou-se o número de fêmeas dessa população levando-se em conta o intervalo médio de registro das crias, isto é, média de 0,71 bezerro registrado/ano/vaca. Com 148.098 animais registrados entre 1995 e 2000 Estatísticas (2001), pode-se determinar a média de 24.683 registros/ano, correspondendo a 34.764 vacas registradas em reprodução no Brasil (24.683/0,71). Segundo critérios da FAO (1992), esse número de vacas coloca a raça na categoria de "monitoração" com relação ao perigo de extinção. Conforme estimativas da FAO (Domestic..., 1998), a população não apresenta risco de extinção, e 100% das fêmeas eram acasaladas com touros da mesma raça o que contribuiu para a evolução do número de animais registrados, confirmado pela estatística apresentada pela ABCZ (Estatísticas (2001)).
Na Tab. 5 são apresentadas as estimativas do intervalo de gerações para as quatro passagens gaméticas, de pais para filhos registrados e de pais para filhos que se tornaram pais, nos qüinqüênios estudados. O menor intervalo obtido no primeiro período pode ser atribuído à forma de truncamento dos dados, visto que foram incluídos animais com nascimento a partir do início do registro em 1969, e as estimativas não apresentaram tendência à estabilização. O último período evidenciou alteração na ordem das passagens, pois tanto pais de filhos como pais de pais permaneceram mais tempo em reprodução, alongando o intervalo nesses passos, diferindo de trabalhos descritos na literatura em que as passagens de pai para filho são menores do que as de mãe para filho (Herron & Pattie, 1977; Toll & Barker, 1979; Vassalo et al., 1986; Razook et al., 1993). Razook et al. (1993) descreveram intervalos médios de pais para filhos de 3,66 e 3,74 anos para Nelore e Guzerá, respectivamente, nos rebanhos de seleção da Estação Experimental de Zootecnia de Sertãozinho - SP, onde os machos iniciam a vida reprodutiva com dois anos de idade e permanecem em serviço por duas estações de monta.
Ao considerar as passagens dos 35 reprodutores com mais de 1.000 filhos, possivelmente utilizados em inseminação artificial, o intervalo nas passagens pai-filho e pai-filha foi de 9,03 e 8,87 anos, respectivamente. Esses touros representaram 0,5% do total de reprodutores e foram pais de 91.994 animais (25,5% dos registrados), com média de 2.628 filhos por reprodutor, ou seja, cerca de 53 vezes maior do que a média geral dos pais. A intensa utilização desses poucos reprodutores como pais contribuiu para o incremento da variância progenitor - progênie que é uma das principais causas da diminuição do tamanho efetivo (Tab.2). Apesar disso, o número de animais sem ascendência conhecida inscritos no livro genealógico aumentou nos períodos (4.630, 10.362, 10.252 e 13.907, respectivamente para 1979-1983, 1984-1988, 1989-1993 e 1994-1998 para animais com ao menos um ancestral desconhecido), mas tal fato não propiciou aumento do Ne.
Determinaram-se o coeficiente de endogamia com sua respectiva taxa de variação e o Ne, representados pela Fig.1. Os valores calculados do primeiro ao último período foram de 0,55%, 0,67%, 0,85% e 0,98% para o FIT, que expressa o coeficiente de endogamia, de 0,41%, 0,42%, 0,43% e 0,40% para o DF, que expressa a taxa de variação, e de 122, 119, 116 e 124 para o Ne. A endogamia mostrou-se crescente do primeiro ao último período, e DF e Ne foram praticamente constantes, exceção encontrada no último período, em que se observou ligeiro acréscimo no Ne, coincidindo com a redução momentânea do DF. Dos 410.853 animais do arquivo de genealogia, 70.949 apresentaram coeficiente de endogamia positivo, média de 4,12% e máximo de 42,18%. Na Tab. 6 são apresentados o número de animais distribuídos por classes de endogamia.
O coeficiente total de endogamia apresentou-se em níveis aceitáveis, atingindo no último período 0,98%, com crescimento de 81,0%. A razão encontrada entre Ne/N no último período foi de 9,37x10-4, evidenciando o acentuado desvio desses dois parâmetros. Goddard & Smith (1990) sugerem Ne mínimo de 40 por geração para maximizar o ganho genético em população selecionada para produção de leite, e Meuwissen & Woolliams (1994) recomendam valores entre 30 e 250 para prevenir decréscimo no valor adaptativo.
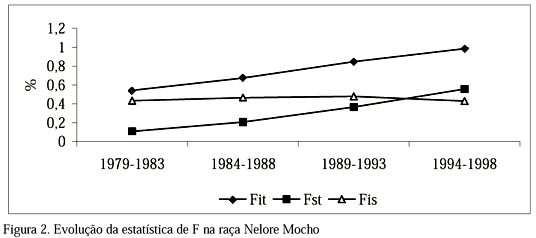
A evolução da estatística de F pode ser vista na Fig. 2. Fst passou de 0,11% no primeiro período para 0,56% no último, FIS manteve-se em torno de 0,45%, ou seja, valores muito próximos a zero ao longo dos períodos, indicando que a subdivisão é praticamente inexistente nesta população. Resultado semelhante foi obtido por Nomura et al. (2001) para a raça Japanese Black. Esses autores citaram que após o início do programa de avaliação genética para a raça, devido à utilização de poucos animais por meio da inseminação artificial, a subdivisão que existia na população praticamente desapareceu. Alguns fatores são sugestivos para explicação da ausência de subdivisão no Nelore Mocho, como a ampla comercialização de sêmen da raça que favorece a migração e disseminação de material genético entre rebanhos, e a utilização de poucos reprodutores com grande contribuição.
Na Tab. 7 encontram-se o número efetivo de fundadores, de ancestrais e de genomas remanescentes calculados para essa população. Segundo Boichard et al. (1997), as análises obtidas dos parâmetros baseados na probabilidade de origem do gene são mais eficientes na predição da variabilidade genética porque são menos sensíveis às falhas ou perdas de informações dos pedigrees. Houve queda na magnitude dos parâmetros estimados entre os períodos, exprimindo redução na variabilidade, os quais seguem ordem decrescente entre si Nfun > Nanc > Ngen. Nfun avalia como se mantém o balanço esperado da contribuição dos fundadores entre as gerações, levando em consideração a taxa de seleção e a variação de tamanho de família. Uma das causas de perda de genes na população é o efeito gargalo, que resulta da diminuição do número de reprodutores entre períodos, e que é tomado em consideração quando da estimação do Nanc, enquanto que Ngen avalia o número de genes fundadores que são mantidos na população para um dado locus. Desse modo, Nfun apresentou os maiores valores por expressar somente a contribuição de fundadores, e Ngen as menores estimativas por considerar todas as possibilidades de perdas de alelos fundadores.
O Nfun estimado no presente trabalho está dentro da faixa de variação descrita por Faria et al. (2002). Esses autores relataram maiores valores do Nfun para a raça Gir seguida da Guzerá, possivelmente em função da predominância de monta natural e existência de subdivisão nas populações, e menores valores para as raças Gir Mocho e Sindi em ordem decrescente. Contudo, apesar de existir subdivisão nesses grupamentos, a razão desse achado pode ser atribuída ao pequeno número de fundadores e machos em uso nas duas últimas raças. O número efetivo de ancestrais no Nelore Mocho revela a predominância na utilização de alguns reprodutores, explicada por sua contribuição marginal junto à população referência (Tab. 8). A razão Nfun/Nanc expressa o efeito "gargalo" e resulta da diminuição do número de reprodutores entre períodos. O valor médio encontrado para o Nelore Mocho foi de 1,44 e para a raça Tabapuã entre 1,02 e 1,44, respectivamente, no primeiro e último períodos conforme Vercesi Filho et al. (2002). Boichard et al. (1997) relataram uma razão igual a 3,0 para a raça Normanda, relacionado à intensa utilização de poucos animais por meio de inseminação artificial. O Nanc do último período no Nelore Mocho foi maior do que os valores relatados na literatura por Vercesi Filho et al. (2002) e Boichard et al. (1997), enquanto que Solkner et al. (1998) citaram valor de 113,7 para a raça Simental na Áustria.
Na Tab. 8 evidencia-se o aumento da contribuição de certos animais. No último qüinqüênio 50 animais respondem por quase metade dos alelos presentes na população. Solkner et al. (1998) encontraram resultado semelhante para a raça Simental, em que 55 animais contribuíram com metade dos alelos, ao passo que na raça Grauvieh esse número reduziu-se a 16. A proporção Ngen/Nfun indica a extensão do processo de deriva genética, e os valores encontrados para o Nelore Mocho oscilam em torno de 0,5. Vercesi Filho et al. (2002) relataram para a raça Tabapuã a razão de 0,63 de 1994 a 1998.
CONCLUSÕES
A utilização mais prolongada de certos reprodutores aumentou o intervalo de gerações das passagens gaméticas pai-filho e pai-filha. O aumento da razão fêmea:macho e da variância da progênie de reprodutores contribuíram para decréscimos na variabilidade genética. Apesar do aumento do número de animais sem genealogia conhecida incluídos no livro genealógico, esse fato não foi suficiente para incrementar a variabilidade genética. O coeficiente de subdivisão populacional (Fis) é praticamente inexistente, indicando ausência de subdivisão na raça. Todas as funções utilizadas para estimar a variabilidade genética no Nelore Mocho, mostraram-se concordantes. Em todas as estimativas a raça apresentou variabilidade. Entretanto, as estimativas de variabilidade merecem atenção e devem ser constantemente monitoradas, evitando complicações que possam ocorrer no futuro.
AGRADECIMENTOS
Os autores agradecem a ABCZ pela concessão dos dados para este trabalho, realizado com apoio do CNPq, CAPES e FAPEMIG. Externam também os agradecimentos aos Drs. D. Boichard, T. Nomura, R.F. Euclydes e T.H.E. Meuwissen pela concessão dos programas, pelo auxílio em sua implementação e pelos comentários que muito contribuíram para a execução deste trabalho.
- BOICHARD, D.; MAIGNEL, L.; VERRIER, É. The value of using probabilities of gene origin to measure genetic variability in a population. Genet. Sel. Evol., v.29, p.5-23, 1997.
- CROW, J.F.; KIMURA, M. An introduction to population genetics theory New York: Harper and Row, 1970.
- DOMESTIC animal diversity information system [s.l.]: FAO, 1998. (CD-Rom).
- ESTATÍSTICAS. Uberaba: ABCZ. <http://www.abcz.org.br> Acesso em 10/04/2001.
- EVOLUÇÃO da Inseminação Artificial no Brasil de 1979-1999", [s.l.]: ASBIA. <http://www.asbia.org.br> Acesso em 25/05/2001.
- FALCONER, D.S.; MACKAY, T. Introduction to quantitative genetics London : Longman Group, 1996.
- FAO. The management of global animal genetic resources. Animal production and health. 104, Rome: FAO, 1992.
- FARIA, F.J.C.; VERCESI FILHO, A.E.; JOSAHKIAN, L.A. et al. Estrutura da população de bovinos Nelore e Nelore Mocho: Resultados preliminares. In: SIMPÓSIO NACIONAL DE MELHORAMENTO ANIMAL, 3, 2000, Belo Horizonte. Anais.. Belo Horizonte, 2000, p.367-370.
- FARIA, F.J.C.; VERCESI FILHO, A.E.; MADALENA, F.E. et al. Parâmetros populacionais do rebanho Sindi registrado no Brasil. Rev. Bras. Zootec., v.30(6S), p.1989-1994, 2001.
- FARIA, F.J.C.; VERCESI FILHO, A.E.; MADALENA, F.E. et al. Pedigree analysis in the Brazilian zebu breeds. In: WORLD CONGRESS ON GENETICS APPLIED TO LIVESTOCK PRODUCTION, 7, 2002, Montpellier. Proceedings.. Montpellier, 2002. (No prelo).
- GODDARD, M.G.; SMITH, C. Optimum number of bull sires in dairy cattle breeding. J. Dairy Sci., v.73, p.1113-1122, 1990.
- HERRON, N.D.; PATTIE, W.A. Studies of the Australian Ilawarra Shorthorn breed of dairy cattle. II. Aust. J. Agric. Res., v.28, p.1119-1132, 1977.
- LACY, R.C. Analysis of founder representation in pedigrees: founder equivalents and founder genome equivalents. Zoo Biol., v.8, p.111-123, 1989.
- MEUWISSEN, T.H.E.; WOOLLIAMS, J. Effective sizes of livestock populations to prevent a decline in fitness. Theor. Appl. Genet, v.89, p.1019-1026 ,1994.
- NOMURA, T.; HONDA, T.; MUKAI, F. Inbreeding and effective population size of Japanese Black cattle. J. Anim. Sci., v.79, p.366-370, 2001.
- RAZOOK, A.G.; FIGUEIREDO, L.A.; BONILHA NETO, L.M. et al. Intensidades de seleção e repostas diretas e correlacionadas em 10 anos de progênies de bovinos das raças Nelore e Guzerá selecionadas para peso pós desmame. Bol. Ind. Anim., v.2, p.147-163, 1993.
- RENDEL, J.M.; ROBERTSON, A. Estimation of genetic gain in milk yield by selection in a closed herd of dairy cattle. J. Genet., v.50, p.1-8, 1951.
- SANTIAGO, A.A. O Zebu na Índia, no Brasil e no mundo Campinas: Instituto Campineiro de Ensino Agrícola, 1986.
- SÖLKNER, J.; FILIPCIC, L.; HAMPSHIRE, N. Genetic variability of populations and similarity of subpopulations in Austrian cattle breeds determined by analysis of pedigrees. Anim. Sci., v.67, p.249-256, 1998.
- TOLL, G.L.; BARKER, J.S.F. Genetic history of the pedigree Poll Hereford breed in Australia: Effect of the importation ban. Aust. J. Agric. Res., v.30, p.767-777, 1979.
- VASSALO, J.M.; DIAZ, C.; GARCIA-MEDINA, J. R. A note on the population structure of the Avileña breed of cattle in Spain. Livest. Prod. Sci., v.15, p.285-288, 1986.
- VERCESI FILHOA.E.; FARIA, F.J.C.; MADALENA, F.E. et al. Estrutura populacional do rebanho Tabapuã registrado no Brasil. Arq. Bras. Med. Vet. Zootec., 2002, (no prelo).
- WRIGHT, S. Evolution in Mendelian populations. Genetics, v.16, p.97-159. 1931.
- WRIGHT, S. Size of population and breeding structure in relation to evolution. Science, v.87, p.430-431, 1938.
Datas de Publicação
-
Publicação nesta coleção
25 Mar 2003 -
Data do Fascículo
Out 2002
Histórico
-
Revisado
27 Ago 2002 -
Recebido
17 Jul 2001