Resumos
Foram estudadas as alterações cutâneas de ratas Wistar adultas castradas e não castradas, mantidas em estado hipotireóideo por 120 dias. Dois grupos eutireóideos, um castrado e outro não castrado, serviram de controle. Secções da pele abdominal ventral e torácica dorsal foram coradas pelas técnicas de HE, PAS, azul de alcian (alcian blue), picro-sirius red-luz polarizada e Verhoeff. Adelgaçamento da epiderme, atrofia de glândulas sebáceas, redução parcial ou total de fibras elásticas e do colágeno do tipo III da derme caracterizaram tanto o hipotireoidismo quanto a castração. Retardo no crescimento dos folículos pilosos e hiperceratose foram vistos apenas na deficiência dos hormônios tireoidianos, independente do estado funcional das gônadas. A associação hipotireoidismo e castração caracterizou-se por adelgaçamento da epiderme da região abdominal ventral e espessamento e aumento da celularidade na região torácica dorsal, com derme adelgaçada e desprovida de fibras elásticas e de colágeno do tipo III. Conclui-se que a deficiência dos hormônios sexuais ou tireoidianos altera todos os componentes da pele e que as lesões são agravadas na associação hipotireoidismo-castração.
rata; pele; hipotireoidismo; castração
The cutaneous alterations of castrated and intact female Wistar rats kept under a hypothyroidism state for 120 days were studied. Two euthyroid groups, one castrated and other intact, served as controls. Ventral abdominal and dorsal thoracic skin sections were stained by HE, PAS, alcian blue, picro-sirius red-polarized light and Verhoeff. Epidermis and sebaceous glands atrophy, partial or total reduction of elastic fibrils and type III dermal collagen were observed in both hypothyroidism and castration. Delay in hair follicle growth and hyperkeratosis were only observed when deficiency of thyroid hormones was present, independent of gonadal function. The association of hypothyroidism and castration was characterized by ventral abdominal epidermis atrophy, thickening and hyperplasia of dorsal thoracic epidermis, dermis atrophy with loss of elastic fibrils and type III collagen. In conclusion, ovarian steroids or thyroid hormones deficiency modifies all the components of the skin and that the injuries are aggravated in the association hypothyroidism-castration.
rat; skin; hypothyroidism; castration
Morfologia e histoquímica da pele de ratas hipotireóideas castradas e não castradas
[Morfology and histochemistry of the skin in hypothyroid castrated and intact rats]
E. Ferreira; R. Serakides; V.A Nunes; M.G. Gomes; C.M. Silva; N.M. Ocarino; A.F.C. Ribeiro
Escola de Veterinária UFMG, Caixa Postal 567 30123-970, Belo Horizonte, MG
Endereço para correspondência Endereço para correspondência E. Ferreira, R. Serakides E-mail: serakide@dedalus.lcc.ufmg.br
RESUMO
Foram estudadas as alterações cutâneas de ratas Wistar adultas castradas e não castradas, mantidas em estado hipotireóideo por 120 dias. Dois grupos eutireóideos, um castrado e outro não castrado, serviram de controle. Secções da pele abdominal ventral e torácica dorsal foram coradas pelas técnicas de HE, PAS, azul de alcian (alcian blue), picro-sirius red-luz polarizada e Verhoeff. Adelgaçamento da epiderme, atrofia de glândulas sebáceas, redução parcial ou total de fibras elásticas e do colágeno do tipo III da derme caracterizaram tanto o hipotireoidismo quanto a castração. Retardo no crescimento dos folículos pilosos e hiperceratose foram vistos apenas na deficiência dos hormônios tireoidianos, independente do estado funcional das gônadas. A associação hipotireoidismo e castração caracterizou-se por adelgaçamento da epiderme da região abdominal ventral e espessamento e aumento da celularidade na região torácica dorsal, com derme adelgaçada e desprovida de fibras elásticas e de colágeno do tipo III. Conclui-se que a deficiência dos hormônios sexuais ou tireoidianos altera todos os componentes da pele e que as lesões são agravadas na associação hipotireoidismo-castração.
Palavras-chave: rata, pele, hipotireoidismo, castração
ABSTRACT
The cutaneous alterations of castrated and intact female Wistar rats kept under a hypothyroidism state for 120 days were studied. Two euthyroid groups, one castrated and other intact, served as controls. Ventral abdominal and dorsal thoracic skin sections were stained by HE, PAS, alcian blue, picro-sirius red-polarized light and Verhoeff. Epidermis and sebaceous glands atrophy, partial or total reduction of elastic fibrils and type III dermal collagen were observed in both hypothyroidism and castration. Delay in hair follicle growth and hyperkeratosis were only observed when deficiency of thyroid hormones was present, independent of gonadal function. The association of hypothyroidism and castration was characterized by ventral abdominal epidermis atrophy, thickening and hyperplasia of dorsal thoracic epidermis, dermis atrophy with loss of elastic fibrils and type III collagen. In conclusion, ovarian steroids or thyroid hormones deficiency modifies all the components of the skin and that the injuries are aggravated in the association hypothyroidism-castration.
Keywords: rat, skin, hypothyroidism, castration
INTRODUÇÃO
O hipotireoidismo é a causa mais comum de dermatose de origem endócrina no cão (Muller et al., 1985) e também no homem. As alterações teciduais já são bem conhecidas e se caracterizam por mixedema, atrofia epidérmica, hiperceratose, redução e mudança da composição das fibras colágenas e elásticas e alopecia (Muller et al., 1985; Feingold, Elias, 1987; Grando, 1993; Doliger et al., 1995).
Também os esteróides sexuais alteram o metabolismo da pele, havendo, na ausência ou inatividade das gônadas, o adelgaçamento da epiderme (Brincat, 2000), a desorganização das fibras colágenas (Kafantari et al., 2000) e a retenção de gordura na hipoderme (Dukes, 1988). Isso porque os hormônios sexuais estimulam o desenvolvimento dos ceratinócitos, prolongam o ciclo de crescimento do folículo piloso, diminuem a secreção sebácea e estimulam a síntese e a maturação do colágeno (Grando, 1993; Brincat, 2000; Kafantari et al., 2000).
O estudo da inter-relação entre os hormônios tireoidianos e sexuais femininos em outros tecidos (Serakides et al., 2000a,b; Gomes et al., 2001) tem demonstrado que a deficiência conjunta pode agravar ou minimizar os efeitos da deficiência isolada de cada um deles. Na pele não se conhece essa inter-relação, mas postula-se que haja homologia entre os receptores da triiodotironina (T3) e dos esteróides sexuais, razão pela qual o efeito desses hormônios é similar na proliferação e diferenciação dos ceratinócitos e na síntese de proteoglicanos e fatores de crescimento (Grando, 1993).
Dessa forma, o estudo da morfologia e das alterações histoquímicas da pele de ratas hipotireóideas castradas é pertinente e se constituiu no objetivo deste trabalho.
MATERIAL E MÉTODOS
Foram utilizadas 40 ratas Wistar, com três meses de idade, alojadas em caixas plásticas, na densidade de cinco ratas/caixa, as quais receberam ração comercial e água destilada ad libitum e foram mantidas em regime de 12 horas de luz e 12 horas de escuro.
A metade das ratas foi castrada e, uma semana após a castração, dois grupos de 10 ratas, um castrado e outro não castrado, foram induzidos ao hipotireoidismo pela administração diária de propiltiouracil (Sigma, St Louis, USA). A droga, diluída em 5ml de água destilada, foi administrada por sonda gástrica, na dose de 1mg/animal/dia, durante 120 dias. Os dois grupos restantes (10 ratas castradas e 10 não castradas) foram mantidos em estado eutireóideo, pela administração de 5 ml de água destilada, como placebo, seguindo-se o mesmo esquema e procedimentos usados na administração da droga antitireoidiana. Com isso, foram formados quatro grupos experimentais ou tratamentos, a saber: (1) hipotireóideo castrado, (2) hipotireóideo não castrado, (3) eutireóideo castrado e (4) eutireóideo não castrado.
Ao final do período experimental, todas as ratas foram sacrificadas após sedação com pentobarbital sódico e colhidos fragmentos da pele abdominal ventral e torácica dorsal que foram fixados em formalina a 10% neutra e tamponada e submetidos ao processamento rotineiro de inclusão em parafina. Secções de 5m m foram coradas pelas técnicas da hematoxilina-eosina (HE) para análise histomorfométrica; PAS ou azul de alcian pH 2,5 (alcian blue) para análise de glicoproteínas; Verhoeff para análise das fibras elásticas (Prophet et al., 1992) e picro-sirius red-luz polarizada para caracterização do tipo de colágeno (Junqueira et al., 1979). Amostras de sangue foram colhidas para determinação da concentração plasmática de T4 livre pela técnica da quimioluminescência (Access Immunoassay System, Sanofi Diagnostics Pasteur Inc., Chaska, MN, USA.), usando-se kit comercial, segundo o protocolo do fabricante.
Em uma secção corada por HE de cada região, foi determinada com auxílio de uma ocular micrométrica e objetiva de 40x a altura da epiderme, em 15 pontos selecionados aleatoriamente. Às médias dos 15 pontos foi aplicado um fator de correção, obtido pela escala de lâmina micrométrica.
O delineamento experimental foi o inteiramente ao acaso, com quatro tratamentos e 10 repetições, sendo cada animal uma repetição. Para cada variável estudada foram determinadas a média e o desvio-padrão. A significância da diferença entre as médias foi verificada pela análise de variância e a comparação entre médias, pelo teste SNK (Sampaio, 1998).
RESULTADOS
Os valores da tiroxina livre estavam significativamente mais baixos nas ratas tratadas com propiltiouracil (Tab. 1), o que confirma a indução e manutenção do estado hipotireóideo pela administração da droga ininterruptamente por 120 dias.
Somente nas ratas hipotireóideas, independente de serem castradas ou não, foram evidenciadas alterações cutâneas macroscópicas. Os pêlos e a pele apresentavam-se amarelados e recobertos por escamas acastanhadas e os pêlos, ressecados e opacos, estavam reduzidos em número.
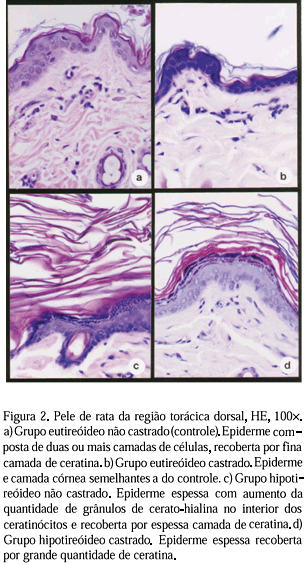
Histologicamente, a pele de ambas as regiões nas ratas eutireóideas não castradas apresentava epiderme composta de duas a quatro camadas de células, recoberta por fina camada de ceratina (Fig. 1a e 2a). Na derme, o tecido conjuntivo era denso e constituído principalmente por fibras colágenas do tipo I e III (Fig. 3a) e pequena quantidade de fibras elásticas (Fig.4a). Os folículos pilosos eram abundantes e em diferentes estádios de diferenciação e as glândulas sebáceas apresentavam células com citoplasma claro e vacuolizado. A castração neste grupo não alterou a camada córnea, mas reduziu significativamente a espessura da epiderme da região abdominal ventral (Tab. 1), constituída na maioria das vezes de somente uma camada de ceratinócitos (Fig. 1b). Nenhuma alteração foi evidenciada na região torácica dorsal (Tab.1 e Fig.2b). A derme, independente da localização, era delgada, apresentava aumento do colágeno do tipo I, em detrimento do colágeno III, geralmente ausente (Fig. 3b) e apresentava-se desprovida de fibras elásticas (Fig. 4b). As glândulas sebáceas apresentavam células com citoplasma pouco volumoso e núcleo com cromatina densa e, às vezes, picnóticos. O número e as características dos folículos pilosos não se alteraram. A hipoderme estava mais espessa e com maior acúmulo de tecido adiposo.
À semelhança das ratas eutireóideas castradas, as hipotireóideas com gônadas funcionais tiveram a altura e a celularidade da epiderme também reduzidas significativamente na região abdominal ventral (Tab.1 e Fig. 1c), mas não na torácica dorsal (Tab. 1 e Fig. 2c). Os ceratinócitos apresentavam, na sua maioria, núcleos achatados e havia grande quantidade de grânulos de cerato-hialina na camada superior do extrato espinhoso, além de espessamento da camada córnea (Fig. 1c e 2c). Essa hiperceratose era mais marcante na região torácica dorsal. Em 70% das ratas, a derme apresentava-se delgada e desprovida de fibras elásticas. Nos 30% restantes, havia apenas redução do número e da espessura das fibras elásticas (Fig. 4c) e, em cerca de 50%, menor número de fibras colágenas do tipo III (Fig. 3c). Os folículos pilosos, presentes em número reduzido, apresentavam-se em telógeno (fase de repouso). As glândulas sebáceas apresentavam células com citoplasma pouco volumoso e núcleo com cromatina densa. A hipoderme apresentava-se espessa com deposição de grande quantidade de tecido adiposo.
Na associação hipotireoidismo-castração, as alterações da epiderme e derme intensificaram-se. Havia hiperceratose intensa (Fig. 1d e 2d) e aumento da quantidade de grânulos de cerato-hialina no interior dos ceratinócitos. Em um dos animais, foi observado um cisto epidermóide. A altura da epiderme da região abdominal ventral era idêntica à dos grupos eutireóideo castrado e hipotireóideo não castrado, mas menor em relação ao controle (Tab. 1 e Fig. 1d). No entanto, na região torácica dorsal a epiderme foi a mais alta (Tab. 1 e Fig. 2d)), dado o aumento da celularidade. A derme, independente da localização anatômica, era intensamente delgada, desprovida de colágeno do tipo III (Fig. 3d) e de fibras elásticas (Fig. 4d), mas com maior quantidade de colágeno do tipo I (Fig. 3d). As glândulas sebáceas pareciam mais ativas e os folículos pilosos, embora ainda reduzidos em número, já se apresentavam em outras fases de seu ciclo, demonstrando discreta retomada do crescimento.
Nem o hipotireoidismo nem a castração, mesmo associados, não alteraram o conteúdo e a composição dos glicosaminoglicanos da derme.
DISCUSSÃO
Sabe-se que no hipotireoidismo humano pode haver menor conversão hepática do b -caroteno, acarretando hipercarotenemia e coloração amarelada da pele (Feingold, Elias,1987). Pela cor amarelada da pele e pêlos observada nas ratas hipotireóideas pode-se suspeitar de dificuldade de metabolização dos carotenóides da dieta também em ratos. A falta de hidratação de pêlos e pele e a descamação crostosa parece ter muito a ver com a interferência com o processo normal de ceratinização da epiderme, como bem demonstrado pelos achados histológicos. Aliás, uma das ações bem conhecidas dos hormônios tireoidianos é a diferenciação epidérmica, agindo em associação com outras moléculas tais como os retinóides e a vitamina D3 (Törmä et al., 1993).
Também os hormônios sexuais agem sobre a pele, mas como os hormônios tireoididanos, induzem a respostas variadas conforme sua localização e características anatômicas. Isso foi devidamente comprovado pela diferença de resposta entre a pele da região abdominal ventral e a da torácica dorsal.
A resposta às deficiências hormonais, associadas ou isoladas, envolveram todos os elementos da pele, destacando-se a dos ceratinócitos. Tanto a proliferação quanto a diferenciação epidérmicas estavam comprometidas.
Os ceratinócitos apresentam receptores para T3 e são capazes de converter T4 em T3, a forma ativa dos hormônios tireoidianos. Entretanto acredita-se que a ação de T3 na diferenciação dos ceratinócitos possa ser indireta e mediada pelo fator de crescimento epidermal (EGF) ou pela expressão dos genes responsáveis pela renovação da epiderme (Hoath et al. 1983; Grando, 1993). Além disso, suspeita-se que parte dos efeitos da deficiência dos hormônios tireoidianos sobre a epiderme possa ser mediada pela deficiência secundária de vitamina A, sabidamente essencial à diferenciação epitelial e à ligação do EGF com a célula. O que já foi determinado é que todas as moléculas que afetam a diferenciação epidérmica (retinóides, vitamina D, T3) ligam-se a receptores intracelulares pertencentes à superfamíla esteróides/hormônio tireoidiano e que a diferenciação dos ceratinócitos é regulada por elas pela transcrição genética (Törmä et al., 1993). Os hormônios sexuais também deveriam ser arrolados entre as moléculas que afetam a diferenciação epidérmica, pois sua deficiência provocou efeitos idênticos aos do hipotireoidismo, o que coaduna com a assertiva de que eles têm modos de ação comuns sobre a pele e determinados pela homologia de seus receptores (Grando, 1993).
Interessante foi o efeito da associação hipotireoidismo-castração na epiderme. Enquanto a ação isolada do hipotireoidismo ou da castração não alterou a epiderme da região torácica dorsal (apenas a da região abdominal ventral), houve hipotrofia da epiderme na região abdominal ventral e hiperplasia na torácica dorsal. É provável que na associação hipotireoidismo-castração um terceiro fator, difícil de ser apontado, tenha determinado ora a indiferenciação, ora a proliferação de ceratinócitos, dependendo das características topográficas e anatômicas da pele. Sabe-se que tanto a castração quanto o hipotireoidismo diminuem a síntese de 1,25 (OH)2 D3, importante para o controle da proliferação e diferenciação dos ceratinócitos (Matsumoto, 1991; Grando, 1993), sendo sua principal ação a de inibir a proliferação e estimular a diferenciação dos ceratinócitos. Assim, seria importante investigar se o aumento da celularidade (hiperplasia) da epiderme das ratas hipotireóideas castradas pode ter sido mediado pela deficiência secundária da vitamina D.
A gênese da hiperceratose no hipotireoidismo ainda não foi totalmente elucidada. Sabe-se que T3 regula diretamente a expressão dos genes da ceratina (Ohtsuki et al., 1992) e, ao regular a diferenciação do ceratinócito, também estaria regulando a maturação epidérmica, o ciclo epitelial e a síntese normal das ceratinas. No entanto, dada a inter-relação entre hormônios e outras moléculas na pele, não se pode descartar a possibilidade da deficiência secundária de vitamina A e de zinco mediar as alterações de conversão da camada córnea (Feingold, Elias,1987; Prasad et al., 1999).
A técnica de coloração pelo picro-sirius e análise sob luz polarizada (Junqueira et al., 1979) mostrou claramente as alterações do colágeno decorrentes do hipotireoidismo e da castração. A diminuição ou mesmo o desaparecimento de determinados tipos de colágeno, comprovam a inibição da síntese, maturação e renovação do colágeno, funções comprovadas desses hormônios (Grando, 1993).
O mixedema é considerado uma das alterações clínicas mais freqüentes em pacientes hipotireóideos. Acredita-se que sua gênese esteja relacionada à diminuição da degradação e conseqüente acúmulo de ácido hialurônico na derme, seguida de retenção de água (Feingold, Elias,1987). Pelas colorações de PAS e azul de alcian, eficientemente utilizadas no diagnóstico do mixedema (Doliger et al., 1995), não foi observada nenhuma alteração do conteúdo ou composição dos glicosaminoglicanos da derme das ratas hipotireóideas. Resultados semelhantes foram obtidos por Wiig et al. (2000) em ratos submetidos à tireoidectomia, que sugeriram a influência do conteúdo e da composição do colágeno para explicar a diferença com os achados no homem.
O ciclo dos folículos pilosos foi afetado somente no hipotireoidismo. O ciclo é dividido em fases: fase de crescimento (anagenia); fase de regressão e remodelação (catagenia) e fase de repouso (telogenia). Acredita-se que as células da papila dérmica presentes na base da porção epitelial do folículo piloso sejam responsáveis pela anagenia, induzindo as células tronco do bulbo folicular a se multiplicarem (Chanda et al., 2000). Apesar do hipotireoidismo ter provocado a parada do crescimento com atrofia folicular e alopecia, o mecanismo de ação dos hormônios tireoidianos sobre o folículo piloso ainda não foi elucidado. Interessante é que, apesar da castração não ter alterado os folículos pilosos das ratas eutireóideas, as ratas hipotireóideas castradas tiveram discreta retomada do crescimento folicular. Isso pode ter sido uma resposta adicional à resposta típica da castração que diminui o intervalo entre a telogenia e anagenia, estimulando o crescimento folicular (Chanda et al., 2000). Além disso, deve também ser considerado que a elevação das concentrações plasmáticas de T4 livre, induzida pela castração nas ratas hipotireóideas, reduziu o efeito inibidor do hipotireoidismo sobre o crescimento folicular.
CONCLUSÃO
A deficiência dos hormônios sexuais e tireoidianos altera todos os componentes da pele, independente da região e característica anatômica, havendo na associação hipotireoidismo-castração o agravamento das alterações cutâneas.
Recebido para publicação em 8 de julho de 2002
Recebido para publicação, após modificações, em 11 de outubro de 2002
- BRINCAT, M.P. Hormone replacement therapy and the skin. Maturitas, v.35, p.107-117, 2000.
- CHANDA, S.; ROBINETTE, C.L.; COUSE, J.F. et al. 17b -Estradiol and ICI-182780 regulate the hair follicle cycle in mice through an estrogen receptor-a pathway. Am. Physiol. Endocrinol. Metab., v.278, p.E202-E210, 2000.
- DOLIGER, S.; DELVERDIER, M.; MORÉ, J. et al. Histochemical study of cutaneous mucins in hypothyroid dogs. Vet. Pathol., v.32, p.628-634, 1995.
- DUKES, H.H. Fisiologia dos animais domésticos. 10.ed. Rio de Janeiro:Guanabara Koogan, 1988. p.477-479.
- FEINGOLD, K.R.; ELIAS, P.M. Endocrine-skin interactions. J. Am. Acad. Dermatol., v.17, p.921-940, 1987.
- GOMES, M.G.; SERAKIDES, R.; SILVA, C.M. et al. Relação tireóide-gônadas e o perfil hematológico de ratas hipotireóideas e eutireóideas. Arq. Bras. Endocrinol. Metabol., v.45, p.s79, 2001.
- GRANDO, S.A. Physiology of endocrine skin interrelations. J. Am. Acad. Dermatol., v28, p.281-292, 1993.
- HOATH, S.B.; LAKSHMANAN J.; SCOTT, S.M. et al. Effect of thyroid hormones on epidermal growth factor concentration in neonatal mouse skin. Endocrinology, v.112, p.308-314, 1983.
- JUNQUEIRA, L.C.U.; BIGNOLAS, G.; BRENTANI, R.R. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histoch. J., v.11, p.447-455, 1979.
- KAFANTARI, H.; KOUNADI, E.; FATOUROS, M. et al. Estructural alterations in rat skin and bone collagen fibrils induced by ovariectomy. Bone, v.26, p.349-353, 2000.
- MATSUMOTO, K. Involvement of endogenously produced 1,25-dihydroxyvitamin D-3 in the growth and differentiation of human keratinocytes. Biochim. Biophys. Acta, v.1092, p.311-318, 1991.
- MULLER, G.H.; KIRK, R.W.; SCOTT, D.W. Dermatologia dos pequenos animais. 3.ed. São Paulo:Manole, 1985. p.517-588.
- OHTSUKI, M.; TOMIC-CANIC, M.; FREEDBERG, I.M. et al. Regulation of epidermal keratin expression by retinoic acid and thyroid hormone. J. Dermatol, v.19, p.774-780,1992.
- PRASAD, R.; KUMAR, V.; KUMAR, R. et al. Thyroid hormones modulate zinc transport activity of rat intestinal and renal brush border membrane. Am. J. Physiol, v.276, p.E774-E778, 1999.
- PROPHET, E.B., MILLS. B., ARRINGTON, J.B. et al. Afip laboratory methods in histotechnology Washington: American Registry of Pathology, 1992. 278p.
- SAMPAIO, I.B.M. Estatística aplicada à experimentação animal Belo Horizonte: FEP/MVZ, 1998. 211p.
- SERAKIDES, R.; NUNES, V.A.; NASCIMENTO, E.F. et al. Hipogonadismo e função tireoidiana em ratas hipertireóideas e eutireóideas. Arq. Bras. Med. Vet. Zootec., v.52, p.571-578, 2000Ş.
- SERAKIDES, R.; NUNES, V.A.; NASCIMENTO, E.F. et al. Relação tireóide-gônadas e níveis plasmáticos de fósforo, cálcio e fosfatase alcalina em ratas. Arq. Bras. Med. Vet. Zootec., v.52, p.579-585, 2000b.
- TÖRMÄ, H.; ROLLMAN, O.; VAHLQUIST, A. Detection of mRNA transcripts for retinoic acid, vitamin D3, and thyroid hormone (c-erb-A) nuclear receptors in human skin using reverse trancription and polymerase chain reaction. Acta Dermatol. Venereol.,v.73, p.102-107, 1993.
- WIIG, H.; REED, R.K.; TENSTAD, O. Interstitial fluid pressure, composition of interstitium and interstitial exclusion of albumin in hypothyroid rats. Am. J. Physiol. Heart Circ. Physiol., v.278, p.H1627-h1639, 2000.
Endereço para correspondência
Datas de Publicação
-
Publicação nesta coleção
23 Abr 2003 -
Data do Fascículo
Fev 2003
Histórico
-
Recebido
08 Jul 2002 -
Aceito
11 Out 2002