Resumos
O objetivo deste trabalho foi determinar a degradação ruminal, pela técnica in situ, da matéria seca (MS) e da proteína bruta (PB) de 10 concentrados protéicos. As degradações potenciais da MS e da PB das farinhas de origem vegetal, soja, algodão, mamona e palmiste, mostraram-se elevadas (próximas a 100%), porém em função de taxas de degradação mais altas para o farelo de soja (10%) e menores para o algodão (4%), mamona (MS:3%; PB:1,2%) e palmiste (1,7%), as degradabilidades efetivas (DE) foram bem superiores para o farelo de soja, independentemente da taxa de passagem, o que a torna a fonte protéica de maior disponibilidade ruminal. O glúten de milho mostrou ser uma fonte protéica de baixa degradabilidade ruminal (DE da PB: 16% para 0,05 de taxa de passagem). Dentre os alimentos de origem animal, a maior degradabilidade potencial da proteína bruta foi verificada para a farinha de carne e ossos ( 75,5%), seguida das farinhas de peixe I (58,5%), de penas e vísceras (52,3%) e de sangue (36,7%). A maior degradabilidade efetiva para a taxa de passagem de 5% foi a da farinha de carne e ossos (51%), seguida da farinha de peixe I (41%), de penas e vísceras (40,0%) e de sangue (33%). A farinha de peixe II apresentou valores muito baixos de degradabilidade, apenas 22% com 48h de incubação ruminal.
bovino; rúmen; digestibilidade in situ; concentrado protéico
This work aimed to determine the in situ rumen digestibility of dry matter (DM) and crude protein (CP) using 10 protein sources. Potential degradabilities of DM and CP of soybean meal, cottonseed meal, castorbean meal and palmist coconut meal were high (around 100%), however due to higher fractional degradation rates of soybean meal (10%) and lower for cottonseed meal (4%), castorbean meal (DM:3%, CP:1.2%) and palmist meal (1.7%), the effective degradability (ED) of soybean was significantly higher despite the rates of passage, showing this protein source has the highest ruminal nitrogen availability (from 86% to 63%, as rate of passage increased from 2 to 8%). Gluten meal is a low rumen degradable protein source (ED of CP: 16% for 0.05 passage rate). Among protein sources of animal origin, the highest potential CP degradability was observed for meat and bone meal (75.5%), followed by fish meal I (58.5%), feather and viscera meal (52.3%), and blood meal (36.7%). The highest ED for a passage rate of 5% was observed for meat and bone meal (51%), followed by fish meal I (41%), feather and viscera meal (40%) and blood meal (33%). Fish meal II showed very low rumen degradability with only 22% of CP disappearance at 48 hours of incubation.
bovine; rumen; in situ; digestibility; protein source
Concentrados protéicos para bovinos. 1. Digestibilidade in situ da matéria seca e da proteína bruta
Protein concentrates for bovines.1. In situ digestibility of dry matter and crude protein
J.F.C. MoreiraI; N.M. RodríguezII, * * Autor para correspondência: E-mail: norberto@vet.ufmg.br ; P.C.C. FernandesIII; C.M. VelosoIV; E.O.S. SalibaII; L.C. GonçalvesII; I. BorgesII; A.L.C.C. BorgesII
IMinistério da Agricultura
IIEscola de Veterinária da UFMG Caixa Postal 567 30123-970 Belo Horizonte, MG
IIIAlltech-Brasil
IVUniversidade Estadual do Sudoeste da Bahia (UESB), Campus de Itapetinga
RESUMO
O objetivo deste trabalho foi determinar a degradação ruminal, pela técnica in situ, da matéria seca (MS) e da proteína bruta (PB) de 10 concentrados protéicos. As degradações potenciais da MS e da PB das farinhas de origem vegetal, soja, algodão, mamona e palmiste, mostraram-se elevadas (próximas a 100%), porém em função de taxas de degradação mais altas para o farelo de soja (10%) e menores para o algodão (4%), mamona (MS:3%; PB:1,2%) e palmiste (1,7%), as degradabilidades efetivas (DE) foram bem superiores para o farelo de soja, independentemente da taxa de passagem, o que a torna a fonte protéica de maior disponibilidade ruminal. O glúten de milho mostrou ser uma fonte protéica de baixa degradabilidade ruminal (DE da PB: 16% para 0,05 de taxa de passagem). Dentre os alimentos de origem animal, a maior degradabilidade potencial da proteína bruta foi verificada para a farinha de carne e ossos ( 75,5%), seguida das farinhas de peixe I (58,5%), de penas e vísceras (52,3%) e de sangue (36,7%). A maior degradabilidade efetiva para a taxa de passagem de 5% foi a da farinha de carne e ossos (51%), seguida da farinha de peixe I (41%), de penas e vísceras (40,0%) e de sangue (33%). A farinha de peixe II apresentou valores muito baixos de degradabilidade, apenas 22% com 48h de incubação ruminal.
Palavras-chave: bovino, rúmen, digestibilidade in situ, concentrado protéico
ABSTRACT
This work aimed to determine the in situ rumen digestibility of dry matter (DM) and crude protein (CP) using 10 protein sources. Potential degradabilities of DM and CP of soybean meal, cottonseed meal, castorbean meal and palmist coconut meal were high (around 100%), however due to higher fractional degradation rates of soybean meal (10%) and lower for cottonseed meal (4%), castorbean meal (DM:3%, CP:1.2%) and palmist meal (1.7%), the effective degradability (ED) of soybean was significantly higher despite the rates of passage, showing this protein source has the highest ruminal nitrogen availability (from 86% to 63%, as rate of passage increased from 2 to 8%). Gluten meal is a low rumen degradable protein source (ED of CP: 16% for 0.05 passage rate). Among protein sources of animal origin, the highest potential CP degradability was observed for meat and bone meal (75.5%), followed by fish meal I (58.5%), feather and viscera meal (52.3%), and blood meal (36.7%). The highest ED for a passage rate of 5% was observed for meat and bone meal (51%), followed by fish meal I (41%), feather and viscera meal (40%) and blood meal (33%). Fish meal II showed very low rumen degradability with only 22% of CP disappearance at 48 hours of incubation.
Keywords: bovine, rumen, in situ, digestibility, protein source
INTRODUÇÃO
Estudos visando conseguir melhor aproveitamento do nitrogênio pelo ruminante são direcionados principalmente na busca do equilíbrio entre a utilização do nitrogênio não protéico, da proteína degradável e da não degradável. Para tal fim, deve-se considerar separadamente a demanda de fontes nitrogenadas dos microrganismos do rúmen e a do animal. O conhecimento da degradação ruminal da proteína desempenha papel fundamental nesse contexto, já que determina não apenas sua contribuição para atender às necessidades dos microrganismos como também à quantidade potencial de proteína que escapa da fermentação ruminal e, conseqüentemente, capaz de fornecer aminoácidos disponíveis para digestão a partir do abomaso.
O metabolismo da proteína no rúmen é particularmente interessante em virtude de resultar de dois processos interativos: da degradação da proteína da dieta e da síntese de proteína microbiana, esta última realizada tanto à custa da proteína degradada como de fontes de nitrogênio não protéico.
Para estar disponível para os microrganismos, a proteína deve passar pelo processo de proteólise e ser hidrolisada a peptídeos e aminoácidos, muitos dos quais são subseqüentemente desaminados. A extensão da degradação de fontes protéicas no rúmen é variável, e as diferenças estão relacionadas principalmente a fatores intrínsecos do alimento, espécie animal e influências da natureza da dieta sobre o ambiente ruminal (Siddons e Paradine, 1983; Min et al., 2002; Volden et al., 2002).
Para animais em crescimento e vacas em lactação, o suprimento de proteína microbiana pode ser insuficiente para atender à demanda de produção. Nestes, as necessidades elevadas de aminoácidos devem ser supridas pelo incremento da proteína de origem dietética que escapa da degradação no rúmen e passa para o trato digestivo posterior, onde será submetida aos processos de digestão e absorção (Orskov et al., 1981; Rodriguez, 1986; Volden et al., 2002).
Este trabalho objetivou avaliar a degradação da matéria seca (MS) e da proteína bruta (PB) de concentrados protéicos no rúmen de bovinos.
MATERIAL E MÉTODOS
Foram utilizados quatro novilhos mestiços europeu-zebu, com idade média de 20 meses e peso médio de 320 kg no início da fase experimental. Os animais possuíam fístulas no rúmen e no duodeno, equipadas com cânulas de borracha flexível com 4,0cm de diâmetro no rúmen e cânulas de material sintético rígido com 3,0cm de diâmetro no duodeno.
A dieta fornecida foi constituída por 60% de feno de capim-braquiária picado e 40% de concentrado composto por 45% de grão de milho moído e 55% de farelo de algodão (Tab. 1). As refeições foram divididas em duas partes iguais e fornecidas às 7 e 19 horas, ocasião em que se adicionava mistura mineral completa correspondente a aproximadamente 1,0% do peso da refeição. O alimento fornecido era totalmente consumido até três horas após colocado à disposição dos animais. Adotou-se o período de 20 dias para adaptação à nova dieta antes de se iniciar a fase experimental propriamente dita.
A degradação in situ foi determinada em sacos confeccionados com "dacron" nas dimensões de 8,0 ´ 16,0 cm e malha de 50 um. As bolsas continham cinco gramas dos alimentos moídos em peneiras de 2,0mm. Os sacos foram introduzidos no rúmen às 7 horas, antes do fornecimento da refeição matutina. A retirada foi em ordem seqüencial, extraindo-se o conjunto de três sacos nos tempos de 6, 12, 24 e 48 horas após serem introduzidos no rúmen.
Os sacos foram lavados em água corrente e secos em estufa de circulação forçada a 65° C. Os resíduos foram moídos em peneira de 1mm, reunidos por animal e por horários e acondicionados em frascos de vidro.
Com três sacos preenchidos e lacrados pelo mesmo processo realizou-se a lavagem em água corrente de forma similar ao procedido com os sacos incubados, com o objetivo de se determinar a fração solúvel.
Amostras do líquido ruminal foram retiradas para análise do pH e concentração do nitrogênio amoniacal nos tempos de 3, 6 e 12 horas após o fornecimento da refeição. O pH no líquido ruminal foi determinado em potenciômetro modelo Beckman Expandomatic. O nitrogênio amoniacal foi determinado por destilação de 50ml de líquido ruminal, logo após sua coleta, com 2,5g de óxido de magnésio, utilizando-se ácido bórico como solução receptora e titulando com ácido clorídrico 0,01 N.
As equações não lineares para o desaparecimento da MS e da PB foram estimadas com o uso do programa Statistic Analysis System (SAS, 1985). A equação de degradação e a degradação efetiva (DE) da MS e da PB dos alimentos foram estimadas utilizando-se os modelos de Orskov e McDonald (1979).
No caso de impossibilidade de utilização dessa última equação, devido a não definição dos parâmetros de degradação, a DE foi estimada pelo modelo de Kristensen et al. (1982). Para determinação da DE após o último tempo de incubação foi adotada a taxa de degradação entre o último intervalo estudado como constante até o tempo presumível para o desaparecimento total do alimento no rúmen, aplicando-se então a fórmula para o cálculo da fração efetivamente degradada após 48 horas:
DE=[(100-DEG48).(exp-kp*t48+exp-kp*t100)/2], em que:
DEG48 é a degradação do nutriente até 48 horas de incubação, kp é a taxa de passagem, t48 é o último horário de incubação (48 horas), e t100 é o tempo de incubação presumível para o desaparecimento total do nutriente no rúmen, adotando-se a taxa de digestão no último intervalo de incubação como constante.
Os alimentos estudados foram: farelos de soja (Glycine max), de algodão (Gossypium sp.), de mamona (Ricinus communis), de palmiste (Elaecis guineensis) e de glúten de milho (Zea mays) e farinhas de carne e ossos, de penas e vísceras, de sangue e de peixe I e II (Tab. 2).
RESULTADOS E DISCUSSÃO
As concentrações médias de N-NH3 e o pH do líquido ruminal nos tempos amostrados foram de 26,1 mg/100ml e 6,8, respectivamente, sem alterações significativas entre tempos, sugerindo a manutenção do ambiente ruminal estável e conseqüente atividade normal das bactérias celulolíticas e proteolíticas (Satter, Slyter, 1974; Van Soest, 1994).
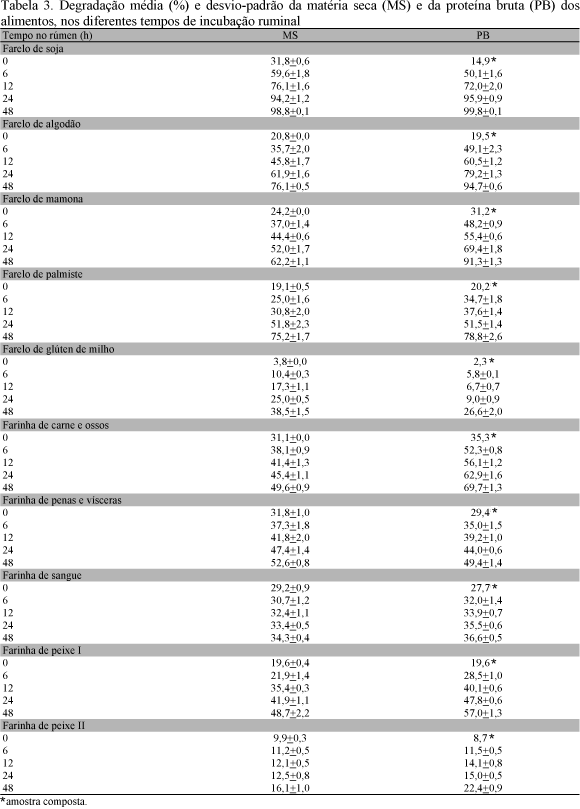
Os valores percentuais médios e desvios-padrão do desaparecimento da MS e da PB dos alimentos testados, nos diferentes tempos de incubação, estão na Tab. 3.
As taxas fracionais de degradação (c) para a MS e a PB do farelo de soja foram de 9,62 e 10,45, respectivamente (Tab. 4). Valores de c dessa amplitude para a MS e a PB do farelo de soja foram observados por Saad (1992), enquanto Valadares Filho (1994) registrou média de 6,62 para quatro ensaios. Fica evidente que o farelo de soja não apenas apresenta alta degradação potencial (DP) ("a" + "b"), mas também rápido desaparecimento no rúmen.
A elevada degradação potencial da PB do farelo de algodão também credencia este concentrado como fonte de alta disponibilidade de nitrogênio para a microbiota ruminal. Entretanto, os valores de desaparecimento da PB nos diversos tempos (Tab. 3) foram um tanto inferiores ao do farelo de soja, devendo o farelo de algodão ter relativamente maior potencial de suprir proteína pós-ruminalmente em comparação ao farelo de soja, caso fossem consideradas as mesmas taxas de passagem. As taxas de degradação foram menores no farelo de algodão (4,0%) se comparadas às do farelo de soja (10%), fato que conduz a valores de degradação efetiva (DE) (Tab. 5) marcadamente inferiores aos do farelo de soja (72% vs 59% para taxa de passagem de 5%). Resultados bem próximos foram encontrados por Saad (1992) para o valor de c da PB (4,3) com a dieta de 25% de concentrado. Os valores encontrados neste trabalho são semelhantes aos observados por Figueira (1991) para a MS (4,2) e para a PB (4,6) aos relatados por Valadares Filho (1994).
A curva de degradação da MS do farelo de mamona procedente de indústria de extração do óleo, submetido à temperatura elevada (não especificada) para fins da eliminação da ricina, apresentou boa correspondência com a equação não linear com alto coeficiente de determinação (98%).
Para a PB, embora os dados observados tenham convergido com a equação não linear (R2=99%), seu desaparecimento não atingiu valor assintótico no último intervalo estudado, o que resultou em estimativa do valor da degradação potencial muito elevado (158,4%). Em vista disso, foi aplicada a equação do segundo grau. Esse comportamento da PB do farelo de mamona pode ser atribuído à desnaturação da proteína durante a aplicação de calor no processo para eliminação dos princípios tóxicos, o que deve ter limitado sua taxa de degradação, tornado-a praticamente constante nos intervalos estudados. O valor do desaparecimento da PB até o tempo de 48 horas (91,3%) e a tendência da curva permitem deduzir que a degradação potencial alcançou valores em torno de 100%, a exemplo do verificado para os farelos de soja e algodão.
A degradação potencial para a MS alcançou valor mais baixo, o que provavelmente está relacionado com a presença de cascas e do envoltório de natureza rígida da semente da mamona.
O valor de c para a MS do farelo de mamona (3,64) foi muito inferior ao obtido para o farelo de soja e semelhante ao do farelo de algodão. Devido à utilização da equação do segundo grau para representar o desaparecimento da PB, o parâmetro c não pode ser calculado para esse nutriente no farelo de mamona, contudo, considerando o comportamento do farelo de mamona nos diversos tempos de incubação e observando os valores da DE determinados conforme Kristensen et al. (1982), pode-se classificá-lo como próximo ao farelo de algodão quanto à capacidade relativa de fornecer nitrogênio para o ambiente ruminal.
O farelo de palmiste apresentou 15,7% de PB (Tab. 2), valor baixo para ser considerado um suplemento protéico, e elevado conteúdo de extrato etéreo (19,1%), indicando que a extração do óleo pela indústria não foi completa. Em amostras desse tipo de alimento Hindle et al. (1995) observaram percentuais de proteína entre 15 e 22%, elevado conteúdo de constituintes da parede celular e proporção variável de extrato etéreo. O baixo conteúdo de PB deve estar relacionado com a presença do tegumento de natureza rígida que envolve o endocarpo e entra na composição do farelo e, também, ao processo de extração do óleo que permitiu proporção elevada de extrato etéreo no produto residual. Devido ao baixo teor de proteína, o fornecimento de nitrogênio para o rúmen pelo farelo de palmiste não pode ser considerado relevante, cerca de 5,9% de PB com o tempo de incubação de 12 horas.
Na composição do farelo entram a fração do endocarpo e o envoltório rígido da amêndoa. Durante o processo de moagem essa última fração é parcialmente pulverizada pelo moinho e mistura-se à gordura do alimento, dificultando a passagem de partículas pela malha da peneira. Embora a moagem do alimento vise uniformizar a amostra e simular a mastigação, características próprias desse e de outros alimentos de natureza similar devem concorrer para a excessiva pulverização e aumentar a quantidade de partículas liberadas pela malha dos sacos de náilon, interferindo nos resultados do desaparecimento da MS. Hindle et al. (1995) afirmam que o tamanho da partícula pode influenciar o desaparecimento de componentes de subprodutos da família das palmáceas. Em virtude da incubação no último intervalo estudado não ter revelado valor assintótico para a degradação do farelo de palmiste, fica inviabilizado o cálculo da degradação potencial da MS e da PB pela equação não linear. Contudo, os valores de desaparecimento da MS e da PB às 48h de incubação foram elevados (75,2% da MS e 79% da PB).
Os valores da DE calculados segundo Kristensen et al. (1982) foram de 63,5%, 39,9% e 32,8% para a MS do farelo de palmiste e de 66,7%, 47,5% e 39,7% para a PB, considerando as taxas de passagem de 0,02, 0,05 e 0,08, respectivamente, valores esses inferiores aos observados para as fontes vegetais já discutidas, porém superiores aos do glúten de milho (Tab. 5).
O farelo de glúten de milho mostrou baixa degradabilidade ruminal. Às 48 horas de incubação apenas 26,6% da PB e 38,5% da MS tinham desaparecido. Embora os dados do desaparecimento da MS tenham convergido com a equação não linear (R2=98), a DP resultante foi muito superior (59,3%) ao valor observado após 48h de incubação. Isso significa que o desaparecimento da MS determinado no último intervalo não foi suficiente para que a curva apresentasse valor assintótico, evidenciando a necessidade de incubação por período mais longo para se atingir o platô. Em vista disso, optou-se pela utilização da equação do segundo grau para representar o desaparecimento da MS (R²=0,99) (Tab. 4).
Para a PB do farelo de glúten de milho os dados observados não convergiram com a equação não linear, possivelmente em virtude da súbita inflexão da curva no intervalo entre 24 e 48 horas de incubação e a reduzida taxa de desaparecimento até 24 horas. Como o efeito quadrático foi significativo, a equação de segundo grau também foi utilizada para representar o desaparecimento da PB (Y = 6,9 0,2 t + 0,0123 t * t ). O incremento da taxa de digestão da PB do farelo de glúten de milho no último intervalo pode estar vinculado aos tipos de proteínas presentes nesse concentrado. A zeína constitui cerca de 60% da proteína do endosperma do milho e se classifica no grupo das prolaminas, que são ricas em aminoácidos hidrófobos e favorecidas pela formação de pontes internas de sulfeto, tornando-as insolúveis e resistentes à degradação. Outro grupo de proteínas do endosperma do milho são as glutelinas, naturalmente insolúveis em agentes fortemente dissociativos e formadas de unidades ligadas por pontes de disulfeto intermoleculares (Larkis, 1981; Payne, Rhodes, 1982). É possível que após o tempo de incubação de 24 horas tenha ocorrido quebra dessas ligações e, como conseqüência, modificações na conformação tridimensional das proteínas, tornando-as mais susceptíveis à ação enzimática.
Pode-se também especular que as proteínas do glúten de milho estariam complexadas com o amido residual do farelo, gelatinizado durante a elaboração do produto, tornando-se mais refratárias à ação das proteases (Nocek e Tamminga, 1991; Orias et al., 2002).
McAllister et al. (1994) afirmaram que a matriz protéica do endosperma do milho e do sorgo é extremamente resistente à adesão e penetração por microrganismos, e apenas os fungos teriam capacidade de colonizar essa estrutura nestes alimentos.
As curvas de desaparecimento e as degradabilidades potenciais da MS nas farinhas de carne e ossos e de penas e vísceras apresentaram valores bem próximos um do outro. Entretanto, o mesmo não aconteceu com as DP da PB, cujos valores foram de 75,5 e 52,3%, respectivamente (Tab.3 e 4). Chama a atenção o alto valor da fração a, que possivelmente deve ser conseqüência do escape de partículas sólidas pelos poros dos saquinhos, já que proteínas de origem animal não se caracterizam pela alta solubilidade. Isso deixa pequenas parcelas da fração lentamente degradada (b). As baixas taxas de degradação da PB das farinhas de carne e ossos e de penas e vísceras (3,3% e 4,2%) levaram a baixos valores de DE (51,4% e 39,8% para 0,05 de taxa de passagem, respectivamente).
Frações relativamente elevadas da MS (29,2%) e da PB (27,7%) na farinha de sangue também desapareceram no tempo zero (Tab. 3). O desaparecimento de material nos tempos subseqüentes foi bastante reduzido, em torno de 3,6% para a MS e de 4,6% para a PB no intervalo entre 6 e 48 horas de incubação, o que faz desse produto fonte nitrogenada pouco disponível para a microbiota ruminal. O valor da DP da proteína foi semelhante ao obtido com 48 horas de incubação e a taxa c foi alta para a pequena fração lentamente degradável (b). A degradação da farinha de sangue foi pouco influenciada pela taxa de passagem e o valor médio de DE da MS foi de 32,6% (Tab. 4). Os baixos percentuais de degradabilidade da farinha de sangue estão de acordo com dados de outros trabalhos (Loerch et al., 1983; Broderick et al., 1988).
A farinha de peixe II apresentou aspecto físico e perfil de degradação muito diferente da farinha de peixe I. A equação de primeiro grau foi a que melhor se ajustou aos dados observados para o desaparecimento da MS e da PB da farinha de peixe II, (MS=10,4+0,1139t e PB=10,2+0,2468t), o que inviabiliza o cálculo da DP. Entretanto, devido aos baixos percentuais de desaparecimento até o tempo de incubação de 48 horas (16,1 e 22,4%, respectivamente), esse alimento apresenta degradação muito reduzida (Tab. 3). Variações na degradação de farinhas de peixe podem ser atribuídas a peculiaridades na composição da matéria prima utilizada, técnicas de processamento e condições de estocagem, o que torna heterogênea a natureza de partidas desse ingrediente. O ARC (1980) e o Ruminant... (1985) atribuem degradações entre 30 e 70% para a PB, desse modo os valores da farinha de peixe II estão fora desse padrão.
CONCLUSÕES
O farelo de soja mostrou ser a fonte protéica de maior degradabilidade efetiva ruminal (77% para 0,05 de taxa de passagem). Embora de degradabilidade potencial similar à soja, os farelos de algodão, mamona e palmiste, devido à menor taxa de degradação, apresentam menor degradabilidade efetiva (59%; 62% e 48%). O glúten de milho é fonte protéica vegetal de baixa degradabilidade ruminal (DE da PB para taxa de 0,05: 16%). A farinha de carne e ossos é a fonte protéica de origem animal de maior degradabilidade efetiva (51% a 0,05), e as de peixe I, penas e vísceras e de sangue as menos degradadas (41%, 40% e 33%, respectivamente).
Trabalho realizado com auxílio financeiro do CNPq, FAPEMIG e FINEP
Recebido para publicação em 3 de julho de 2002
Recebido para publicação, após modificações, em 9 de janeiro de 2003
- ARC. The nutrient requirements of ruminant livestock Agricultural Research Council, England, 1980. 351p.
- BRODERICK, G.A.; WALLACE, R.J.; 0RSKOV, E.R. et al. Comparison of estimates of ruminal protein degradation by in vitro and in situ methods. J. Anim. Sci., v.66, p.1739-1745, 1988.
- FIGUEIRA, D.G. Efeito do nível de uréia sobre as digestibilidades aparente e "in situ", e a dinâmica da fase sólida em bovinos alimentados com cana-de-açúcar e farelo de algodão. 1991. 123f. Tese (Mestrado). Escola de Veterinária, Universidade Federal de Minas Gerais, Belo Horizonte.
- HINDLE, V.A.; STEG, A.; VAN VUUREN, A.M. et al. Rumen degradation and pos-ruminal digestion of palm kernel by-products in dairy cows. Anim. Feed Sci. Technol., v.51, p.103-121, 1995.
- KRISTENSEN, E.S.; M0LLER, P.D.; HVELPLUND, T. Estimation of the effective protein degradability in the rumen of cows using the nylon bag technique combined with the outflow rate. Acta Agric. Scand., v.32, p.123-127, 1982.
- LARKIS, B.A. Seed storage proteins: Characterization and biosynthesis. In: STUMPF, P.K.; CONN, E.E. (Eds.). The biochemistry of plants: A compreensive treatise. v.6. Proteins and nucleic acids. New York: Academic, 1981, p.449-489.
- LOERCH, S.C.; BERGER, L.L.; PLEGGE, S.D. et al. Digestibility and rumen escape of soybean meal, blood meal, meat and bone meal and dehydrated alfafa nitrogen. J. Anim. Sci., v.57, p.1037-1047, 1983.
- McALLISTER, T.A.; BAE, H.D.; JONES, G.A. et al. Microbial attachment and feed digestion in the rumen. J. Anim. Sci., v.72, p.3004-3018, 1994.
- MIN, B.R.; ATTWOOD, G.T.; REILLY, K. at al. Lotus corniculatus condensed tannins decrease in vivo populations of proteolytic bacteria and affect nitrogen metabolism in the rumen of sheep. Can. J. Microbiol., v.48,p.911-921, 2002.
- NOCEK, J.E.; TAMMINGA, S. Site of digestion of starch in the gastrointestinal tract of dairy cows and its effects on milk production and composition. J. Dairy Sci., v.74, p.3598-3629, 1991.
- ORIAS, F.; ALDRICH, C.G.; ELIZALDE, J.C. et al. The effects of dry extrusion temperature of whole soybeans on digestion of protein and amino acids by steers. J. Anim. Sci., v.80, p.2493-2501, 2002.
- ORSKOV, E.R.; HUGHES-JONES, M.; McDONALD, I. Degradability of protein supplements and utilization of undegraded protein by high-producing dairy cows. In: HARESIGN, W.; COLE, D.J.A. (Eds.) Recent developments in ruminant nutrition. London: Butterworths, 1981. p.17-30.
- ORSKOV, E.R.; McDONALD, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci., v.92, p.499-503, 1979.
- PAYNE, P.I.; RHODES, A.P. Cereal storage proteins: Structure and role in agricultural and food technology. In: BOULTHER, D.; PARTHIER, B. (Eds.). Nucleic acids and proteins in plants. I. Structure, biochemistry and physiology of proteins. Berlin: Springer:Verlag, 1982. p.346-369. (Encyclopedia of plant physiology. v.14 A)
- RODRIGUEZ, N.M. Importância da degradabilidade da proteína no rúmen para a formulação de rações para ruminantes. Cad. Téc. Esc. Vet. UFMG, n.1, p.27-45, 1986.
- RUMINANT nitrogen usage. Washington, D.C.: National Academy Press, 1985. 138p.
- SAAD, C.E.P. Degradabilidade in situ dos farelos de soja e de algodão e das farinha de carne e osso e de peixe sob duas relações concentrado:volumoso em bovinos. 1992. 88f. Tese (Mestrado). Escola de Veterinária, Universidade Federal de Minas Gerais, Belo Horizonte.
- SAS user's guide: statistics. 5.ed. Cary: SAS Institute Inc., v.1, 1985. 956p.
- SATTER, L.D.; SLYTER, L.L. Effect of ammonia concentration on rumen microbial protein production in vitro. Br. J. Nutr., v.32, p.199-205, 1974.
- SIDDONS, R.C.; PARADINE, J. Protein degradation in the rumen of sheep and cattle. J. Sci. Food Agric., v.34, p.701-708, 1983.
- VALADARES FILHO, S.C. Utilização da técnica in situ para avaliação dos alimentos. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 31, 1994, Maringá. Anais.. Maringá:SBZ, 1994. p.95-118.
- VAN SOEST, P.J. Nutritional ecology of the ruminant. 2.ed. Ithaca: Cornell University. 1994. 476p.
- VOLDEN, H.; MYDLAND, L.T.; OLAISEN, V. Apparent ruminal degradation and rumen escape of soluble nitrogen fractions in grass and grass silage administered intraruminally to lactating dairy cows. J. Anim. Sci., v.80, p.2704-2716, 2002.
Datas de Publicação
-
Publicação nesta coleção
08 Out 2003 -
Data do Fascículo
Jun 2003
Histórico
-
Aceito
09 Jan 2003 -
Recebido
03 Jul 2002