Abstracts
The association between single-strand conformation polymorphism (SSCP) in the gene of insulin-like growth factor-I receptor (IGF-IR) and age at first calving (AFC), calving interval (CI), lactation length (LL), and milk yield (MY) was studied using 106 graded Holstein females. The polimerase chain reaction (PCR) with specific initiating oligonucleotides, resulted an amplified fragment of 335pb. The population genotypes frequencies were 82.1% and 17.9%, for AA and AB genotypes, respectively. The frequency of A allele was 0.91 and 0.09 of B allele. No association between the identified polymorphism and AFC, CI, and MY was observed. The LL was positively associated (P<0.05) with the absence of B allele. Animals carrying the AA genotype presented a longer lactation period.
beef cattle; reproductive efficiency; post-calving; candidate gene
Estudou-se a associação entre polimorfismos de conformação de fita simples (SSCP) no gene do receptor do fator-I de crescimento semelhante à insulina (IGF-IR) e idade ao primeiro parto (IPP), intervalo entre partos (IEP), duração da lactação (DL) e produção de leite (PL), em 106 fêmeas puras por cruza da raça Holandesa. A reação em cadeia da polimerase (PCR) com oligonucleotídeos iniciadores específicos gerou um fragmento de 335pb. A freqüência genotípica da população para o polimorfismo foi 82,1% de indivíduos homozigotos para o alelo A e 17,9% de heterozigotos (AB). A freqüência do alelo A foi 0,91 e a do alelo B, 0,09. Não foi encontrada associação entre o polimorfismo estudado e as características IPP, IEP e PL. A característica DL foi positivamente associada (P<0,05) à ausência do alelo B. A lactação dos animais portadores do genótipo AA foi mais longa.
IGF-IR; eficiência produtiva; eficiência reprodutiva; gado de leite; gene candidato
ZOOTECNIA E TECNOLOGIA E INSPEÇÃO DE PRODUTOS DE ORIGEM ANIMAL
Associação entre polimorfismos do gene IGF-IR e características produtivas e reprodutivas em fêmeas bovinas da raça Holandesa
Association between IGF-IR gene polymorphisms and productive and reproductive traits in Holstein cows
W. SchoenauI; P.M. PorciunculaII; G. ZamberlanII; F.S. MesquitaIII; V. VieiraIV; J.F.C. OliveiraV; P.B.D. GonçalvesV, * * Autor para correspondência ( corresponding author) E-mail: bayard@biorep.ufsm.br
IDepartamento de Fisiologia UFSM - Santa Maria, RS
IIAluno de graduação UFSM Santa Maria, RS
IIIMédico veterinário autônomo
IVMédico veterinário COASUL São João, PR
VDepartamento de Clínica de Grandes Animais UFSM Faixa de Camobi, km 9 Campus Universitário 97105-900 - Santa Maria, RS
RESUMO
Estudou-se a associação entre polimorfismos de conformação de fita simples (SSCP) no gene do receptor do fator-I de crescimento semelhante à insulina (IGF-IR) e idade ao primeiro parto (IPP), intervalo entre partos (IEP), duração da lactação (DL) e produção de leite (PL), em 106 fêmeas puras por cruza da raça Holandesa. A reação em cadeia da polimerase (PCR) com oligonucleotídeos iniciadores específicos gerou um fragmento de 335pb. A freqüência genotípica da população para o polimorfismo foi 82,1% de indivíduos homozigotos para o alelo A e 17,9% de heterozigotos (AB). A freqüência do alelo A foi 0,91 e a do alelo B, 0,09. Não foi encontrada associação entre o polimorfismo estudado e as características IPP, IEP e PL. A característica DL foi positivamente associada (P<0,05) à ausência do alelo B. A lactação dos animais portadores do genótipo AA foi mais longa.
Palavras-chave: IGF-IR, eficiência produtiva, eficiência reprodutiva, gado de leite, gene candidato
ABSTRACT
The association between single-strand conformation polymorphism (SSCP) in the gene of insulin-like growth factor-I receptor (IGF-IR) and age at first calving (AFC), calving interval (CI), lactation length (LL), and milk yield (MY) was studied using 106 graded Holstein females. The polimerase chain reaction (PCR) with specific initiating oligonucleotides, resulted an amplified fragment of 335pb. The population genotypes frequencies were 82.1% and 17.9%, for AA and AB genotypes, respectively. The frequency of A allele was 0.91 and 0.09 of B allele. No association between the identified polymorphism and AFC, CI, and MY was observed. The LL was positively associated (P<0.05) with the absence of B allele. Animals carrying the AA genotype presented a longer lactation period.
Keywords: beef cattle, reproductive efficiency, post-calving, candidate gene
INTRODUÇÃO
O sistema de fatores de crescimento semelhantes à insulina (IGFs) afeta o desenvolvimento, a lactação e a reprodução animal por meio do estímulo do crescimento, diferenciação, metabolismo e sobrevivência de vários tipos celulares, utilizando vias endócrinas, parácrinas e autócrinas (Werner e LeRoith, 2000). Este sistema é composto por dois ligantes, fatores-I e -II de crescimento semelhantes à insulina (IGF-I e IGF-II), dois receptores (IGF-IR e IGF-IIR/M6P), seis proteínas ligantes ou transportadoras com alta afinidade (IGFBPs-1 a -6) e várias proteases que regulam a disponibilidade de IGFBPs, e, conseqüentemente, a biodisponibilidade de IGFs (Grimber e Cohen, 2000).
A compreensão dos efeitos e interações do sistema IGF sobre a produção e a reprodução em bovinos de leite é de suma importância. Diversas ações sobre a atividade ovariana, incluindo o sinergismo e a amplificação dos efeitos das gonadotrofinas no crescimento e na esteroidogênese das células ovarianas (Monget et al., 2002) bem como no estabelecimento da dominância folicular (Rivera et al., 2001) são afetadas pelo sistema IGF. A relação entre a biodisponibilidade de IGF-I intrafolicular e a viabilidade folicular (Rivera et al., 2001) e a participação no eixo somatotrófico na adaptação do organismo de animais ao balanço energético negativo (Renaville et al., 2002) são fatores determinantes do sucesso reprodutivo.
O IGF-I é conhecido por regular o desenvolvimento da glândula mamária por meio de seus efeitos tanto na progressão do ciclo celular como na apoptose (Hadsell et al., 1996), com implicações na regulação da involução da glândula mamária (Harrington et al., 1994). A administração exógena de somatotrofina bovina (bST) aumentou o número de receptores de IGF-I no tecido mamário de vacas em lactação (Glimm et al., 1988) bem como aumentou a concentração circulante de IGF-I, sugerindo um papel deste fator de crescimento na estimulação da produção de leite mediada pelo bST, caracterizado pela forma da curva de lactação, indicando tanto taxas aumentadas de síntese como aumento no número ou sobrevivência de células mamárias (Bauman et al., 1985).
O receptor de IGF-I, um heterotetrâmero, consiste de duas subunidades a e duas b, membro da família de receptores dos fatores de crescimento tirosino quinase (Ulrich et al., 1986), estruturalmente e funcionalmente homólogo ao receptor da insulina (LeRoith et al., 1995). A ativação deste receptor e sua atividade intrínseca tirosino-quinase iniciam uma cascata de sinalizações envolvendo várias vias intracelulares, como a do fosfatidilinositol-3-quinase (PIK3) e a da proteíno-quinase ativada por mitógenos (MAPK; Butler et al., 1998a,b), que são responsáveis pela diversidade de ações alvo desses fatores de crescimento (Vincent e Feldman, 2002).
Em humanos, o gene que codifica para o IGF-IR está localizado no cromossomo 15, contido em 21 exons. Apresenta tamanho total de, aproximadamente, 100 quilobases (Abbott et al., 1992). Nos bovinos, o gene está localizado no cromossomo 21 (Moody et al., 1996).
O objetivo deste trabalho foi determinar associações entre um polimorfismo do gene do IGF-IR (Mesquita, 2003) e a eficiência produtiva e reprodutiva em vacas da raça Holandesa.
MATERIAL E MÉTODOS
Utilizaram-se fêmeas puras por cruza da raça Holandesa de propriedades localizadas no município de São João, no noroeste do Paraná. Todas as propriedades têm o mesmo controle de manejo implementado pelos técnicos da cooperativa. Foi realizada uma pré-seleção dos animais pela análise detalhada das fichas individuais, utilizando-se animais com, no mínimo, três partos, descartando-se os que apresentavam alterações de ordem sanitária que comprometiam sua capacidade produtiva e reprodutiva. Foram utilizados os dados da idade ao primeiro parto (IPP), do intervalo entre partos (IEP), da duração da lactação (DL) e da produção de leite (PL) para associação com o polimorfismo pesquisado.
O sangue foi obtido da veia jugular, com o uso de anticoagulante (ACD - 0,8 % ácido cítrico, 2,2% citrato de sódio e 2,4% de glicose), e o DNA genômico extraído por meio da técnica descrita por Plante et al. (1992). Foram sintetizados oligonucleotídeos iniciadores específicos a partir da seqüência do loco IGF-IR disponível em bancos de dados públicos na Internet (NCBI GenBank - número de acesso: BTU33122), com as seqüências: 5'-ACCCGCCAAGAAATTGTTTC-3' e 5'-GGCTCCTCCATACTTCCTGTA-3'. A reação para um volume final de 25µl foi de: ~50ng de DNA, 0,4µM de cada oligonucleotídeo iniciador; 50mM KCl; 1,5mM MgCl2; 10mM Tris-HCl (pH 9,0), 50µM de cada dNTP e 1,25U de Taq DNA polimerase. As condições de reação foram as seguintes: desnaturação a 94°C por 3min; 30 ciclos de 1min a 94°C; 30seg a 55°C, 1min a 72°C e ciclo final de extensão de 4min a 72°C.
O controle de amplificação foi realizado em gel de agarose 3% a 110V e caracterização do tamanho do fragmento (335pb) mediante comparação com marcador de peso molecular conhecido (escada de 50bp). Os géis foram corados com brometo de etídio, e a visualização foi realizada sobre luz ultravioleta. Após o controle da amplificação, foram utilizados 8µl do produto da reação de PCR aos quais foram adicionados 8µl de solução desnaturante (formamida 95%; 20mM EDTA, xileno-cianol 0,5%, azul de bromofenol 0,5% e água). As amostras foram submetidas a aquecimento (100°C por 10min) e rapidamente aplicadas em gel de poliacrilamida 10%, 5% de glicerol e TBE (0,09M Tris-borato, 0,002M EDTA). A eletroforese foi realizada em tampão TBE 1x, consistindo de pré-migração a 60mA por 20 minutos e posteriormente a 3W por 20 horas. As amostras foram visualizadas por meio da coloração por nitrato de prata e registradas utilizando o sistema de fotodocumentação Alpha Digidoc 1000.
A associação entre os marcadores genéticos e os desempenhos produtivo e reprodutivo foi avaliada usando-se análise de variância simples, tendo como variável independente o genótipo e, como variáveis dependentes, o IPP, o IEP, a DL e PL. Foi utilizado o pacote estatístico SAS (Statistical... 1988).
RESULTADOS E DISCUSSÃO
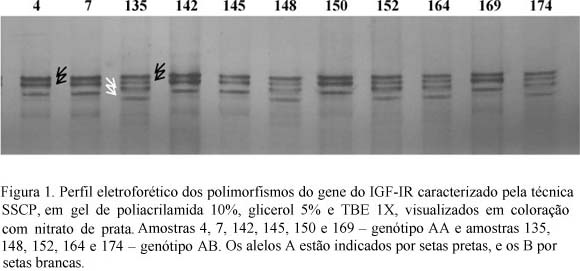
Em 106 indivíduos, a análise de PCR-SSCP evidenciou polimorfismo em que o alelo A apresentou freqüência de 0,91 e o alelo B, 0,09. A freqüência genotípica do homozigoto AA foi 82,1%, e a do genótipo heterozigoto AB, 17,9% (Fig. 1).
Mesquita (2003), ao utilizar o mesmo processo em animais da raça Brangus-Ibagé, obteve freqüências de 0,8 para o alelo A, 64% de animais homozigotos AA, 32% de heterozigotos e 4% de homozigotos para o alelo B, este último não observado nos animais deste experimento. Essas diferenças refletem, provavelmente, características intrínsecas de cada população estudada, bem como a origem da própria raça.
Pela análise da variância (Tab. 1), a DL mostrou associação significativa com os animais portadores do genótipo AA, que evidenciaram um período de lactação significativamente mais longo (P<0,05), indicando provavelmente efeito negativo do alelo B nos animais portadores. Esse resultado pode expressar a ação do IGF-I na inibição da apoptose das células da glândula mamária (Hadsell et al., 1996), retardando a involução da glândula (Harrington et al., 1994), com conseqüente aumento no período de lactação. Deve-se observar, no entanto, que o período de lactação é um critério diretamente relacionado ao manejo adotado em cada propriedade, pela influência da gestação e produção individual de leite.
Não foi observada associação entre a produção de leite e os dois genótipos encontrados. Isso pode ser justificado por modelos de cultura celular mamária de bovinos, em que o IGF-I claramente estimulou a proliferação das células mamárias (Baumrucker e Stemberger, 1989) de modo autócrino ou parácrino (Romagnolo et al., 1992), mas foi ineficaz como lactogênico (Peri et al., 1992).
Mesquita (2003) observou influência positiva de animais homozigotos para o alelo B na raça Brangus-Ibagé, que apresentaram intervalo entre partos menor do que os demais animais. A mesma característica estudada neste experimento não evidenciou associação com os alelos observados (Tab. 1). Não foram identificados animais homozigotos para o alelo B.
Considerado um dos eventos fundamentais na vida reprodutiva de uma fêmea bovina, e incluído como critério de seleção nos sistemas de produção de bovinos de leite e de corte (Patterson et al., 1992), a idade ao primeiro parto não evidenciou associação entre os diferentes genótipos caracterizados, o que exclui o marcador estudado como instrumento de seleção para essa característica.
Os resultados indicam que polimorfismos em genes candidatos com qualidades que sejam de interesse econômico devem ser procurados e pesquisados, propiciando ferramentas valiosas para a seleção de animais/populações com características produtivas e reprodutivas desejadas. Em função deles, pode-se concluir que o polimorfismo estudado em fêmeas bovinas da raça Holandesa no gene do IGF-IR não está associado com a IPP, IEP e PL. No entanto, está associado ao tempo de lactação. Animais portadores do genótipo AA têm período de lactação mais prolongado que os animais não portadores desse genótipo.
AGRADECIMENTOS
Ao médico veterinário Valmir Vieira e à Cooperativa Agropecuária Sudoeste Ltda. (COASUL), por viabilizarem a coleta das amostras e o acesso às informações sobre os animais deste experimento.
Recebido para publicação em 19 de janeiro de 2004
Recebido para publicação, após modificações, em 9 de novembro de 2004
- ABBOTT, A.M.; BUENO, R.; PEDRINI, M.T. et al. Insulin-like growth factor I receptor gene structure. J. Biol. Chem., v.267, p.10759-10761, 1992.
- BAUMAN, D.E.; EPPARD, P.J.; DeGEETER, M.J. et al. Response of high-producing dairy cows to long-term treatment with pituitary and recombinant somatotropin. J. Dairy Sci., v.68, p.1352-1362, 1985.
- BAUMRUCKER, C.R.; STEMBERGER, B.H. Insulin and insulin-like growth factor-I stimulate DNA synthesis in bovine mammary tissue in vitro. J. Anim. Sci., v.67, p.3503-3514, 1989.
- BUTLER, A.A.; BLAKESLEY, V.A.; POULAKI, V. et al. Stimulation of tumor growth by recombinant human insulin-like growth factor-I (IGF-I) is dependent on the dose and the level of IGF-I receptor expression. Cancer Res., v.58, p.3021-3027, 1998a.
- BUTLER, A.A.; YAKAR, S.; GEWOLB, I.H. et al. Insulin-like growth factor-I receptor signal transduction: at the interface between physiology and cell biology. Comp. Biochem. Physiol., v.121, p.19-26, 1998b.
- GLIMM, D.R.; BARACOS, V.E.; KENNELLY, J.J. Effect of bovine somatotropin on the distribution of immunoreactive insulin-like growth factor-I in lactating bovine mammary tissue. J. Dairy Sci., v.71, p.2923-2935, 1988.
- GRIMBER, A.; COHEN, P. Role of insulin-like growth factors and their binding proteins in growth control and carcinogenesis. J. Cell. Physiol., v.183, p.1-9, 2000.
- HADSELL, D.L.; GREENBERG, N.M.; FLIGGER, J.M. et al. Targeted expression of des(1-3) human insulin-like growth factor I in transgenic mice influences mammary gland development and IGF-binding protein expression. Endocrinology, v.137, p.321-330, 1996.
- HARRINGTON, E.A.; BENNETT, M.R.; FANIDI, A. et al. c-Myc-induced apoptosis in fibroblasts is inhibited by specific cytokines. Eur. Med. Biol. Organ. J., v.13, p.32863295, 1994.
- LeROITH, D.; WERNER, H.; BEITNER-JOHNSON, D. et al. Molecular and cellular aspects of the insulin-like growth factor I receptor. Endocrinol. Rev., v.20, p.143-163, 1995.
- MESQUITA, F.S. Associação entre polimorfismos no gene do receptor de IGF-I e características produtivas em bovinos Brangus-Ibagé. 2003. 42f. Dissertação (Mestrado) Universidade Federal de Santa Maria, Santa Maria, RS.
- MONGET P.; FABRE, S.; MULSANT, P. et al. Regulation of ovarian folliculogenesis by IGF and BMP system in domestic animals. Domest. Anim. Endocrinol., v.23, p.139-54, 2002.
- MOODY, D.E.; POMP, D.; BARENDSE, W. Linkage mapping of the bovine insulin-like growth factor-1 receptor gene. Mamm. Genome, v.7, p.168-169, 1996.
- PATTERSON, D.J.; CORAH, L.R.; BRETHOUR, J.R. et al. Evaluation of reproductive traits in Bos taurus and Bos indicus crossbred heifers: relationship of age an puberty to length of the postpartum interval to estrus. J. Anim. Sci., v.70, p.1994-1999, 1992.
- PERI I, S.; SHAMAY, A.; McGRATH, M.F. et al. Comparative mitogenic and galactopoietic effects of IGF-I, IGF-II and des-3-IGF-I in bovine mammary gland in vitro Cell Biol. Int. Rep., v.16, p.359368, 1992
- PLANTE, Y.; SCHMUTZ, S.M.; LANG, K.D.M. Restriction fragment length polymorphism in the mitochondrial DNA of cloned cattle. Theriogenology, v.38, p.897-904, 1992.
- RENAVILLE, R.; HAMMADI, M.; PORTELLE, D. Role of somatotropic axis in the mammalian metabolism. Domest. Anim. Endocrinol., v.23, p.351-360, 2002.
- RIVERA, G.M.; CHANDRASEKHER, Y.A.; EVANS, A.C.O. et al. A potential role for insulin-like growth factor protein-4 proteolysis in the establishment of ovarian follicular dominance. Biol. Reprod., v.65, p.102-111, 2001.
- ROMAGNOLO, D.; AKERS, R.M.; WONG, E.A. et al. Overexpression of ovine insulin-like growth factor-I stimulates autonomous autocrine or paracrine growth in bovine mammary-derived epithelial cells. Mol. Endocrinol., v.6, p.1774-1780, 1992.
- STATISTICAL analysis system. 6. 3.ed. Cary, NC:SAS Institute, 1988. 1028p.
- ULRICH, A.; GRAY, A.; TAM, A.W. et al. Insulin-like growth factor I receptor primary structure: comparison with insulin receptor suggests structural determinants that define functional specificity. Eur. Med. Biol. Organ. J., v.5, p.2503-2512, 1986.
- VINCENT, A.M.; FELDMAN, E.L. Control of cell survival by IGF signaling pathways. Growth Horm. IGF Res., v.12, p.193-197, 2002.
- WERNER, H.; LeROITH, D. New concepts in regulation and function of the insulin-like growth factors: implications for understanding normal growth and neoplasia. Cell. Mol. Life Sci., v.57, p.932-942, 2000.
Publication Dates
-
Publication in this collection
18 Apr 2006 -
Date of issue
Dec 2005
History
-
Accepted
09 Nov 2004 -
Received
19 Jan 2004