Resumos
Foram utilizados 12 cordeiros machos, inteiros, com peso médio de 31,6kg. Os cordeiros foram alimentados com dietas à base de milho e farelo de soja, suplementadas com diferentes fontes de cálcio: calcário calcítico (CC), feno de alfafa (FA), polpa cítrica (PC) e farinha de conchas de ostras (FCO). Os animais receberam injeção de 7,4 MBq do radiofósforo (32P) e após sete dias foram abatidos para posteriores análises. Utilizou-se o delineamento em blocos ao acaso, com quatro tratamentos e três repetições. A taxa de retenção de 32P, as atividades específicas relativas e padronizadas, e a quantidade de P inorgânico nos tecidos foram iguais para todos os tratamentos (P>0,05). A comparação entre os tecidos mostrou que a concentração de P inorgânico e a taxa de retenção de 32P (P<0,01) foram mais altas nos ossos em relação aos tecidos moles (P<0,01). A retenção de 32P apresentou a seguinte ordem decrescente: rins, fígado, coração, músculos e ossos (P<0,01). A reabsorção óssea de P foi maior que a deposição de P, levando ao balanço negativo do elemento para a maioria dos animais. Concluiu-se que as diferentes fontes de cálcio não interferiram na cinética do P e que a quantidade de P ingerida não atendeu às exigências de P dos animais.
ruminante; radiofósforo; ossos; tecidos moles; retenção
Sixteen growing rams, averaging 31.6kg were studied. They were fed diets based on corn and soya meal, supplemented with different sources of calcium: limestone (CC), alfalfa hay (FA), citrus pulp (PC) and oyster shell meal (OSM). The animals received an injection of 7.4 MBq of radioactive P (32P) and after 7 days they were slaughtered for further analyses. Experimental measurements were analyzed as a completely randomized design with four treatments and three repetitions. The rate of 32P retention, the relative specific activity, the standardized specific activity and the inorganic P in tissue were similar for all treatments (P>0.05). The comparison between tissues showed that the concentration of inorganic P and rate of 32P retention was greater in bone (P<0.01) than in soft tissue. The 32P retention presented the following decreasing order: kidney, liver, heart, muscle and bone (P<0.01). P resorption in bone was higher than P accreted in bone, leading to a negative balance of P in bone for the majority of animals. The means for P accreted in bone were 1.56, 3.29, 2.74 and 2.06g/animal and the means for P resorption in bone were 2.82, 4.51, 4.09 and 3.05g/animal for limestone (CC), alfalfa hay (FA), citrus pulp (PC) and oyster shell meal (OSM), respectively. P resorption values were higher than P accretion in bone leading to a negative balance of P for the majority of animals. It was concluded that the different sources of calcium did not interfere in the kinetics of phosphorus and the amount of P absorbed did not attend P demands of the animals.
ruminant; radiophosphorus; bones; soft tissues; retention
ZOOTECNIA E TECNOLOGIA E INSPEÇÃO DE PRODUTOS DE ORIGEM ANIMAL
Cinética do fósforo em tecidos de ovinos em crescimento alimentados com diferentes fontes de cálcio
Kinetics of phosphorus in tissues of lambs fed different sources of calcium
R.S. DiasI; A. P. RoqueI; D.M.S.S.VittiI; I.C. BuenoII; P.B. GodoyI
ICentro de Energia Nuclear na Agricultura - USP Caixa Postal 96 13400-970 Piracicaba, SP
IIUniversidade de Brasília - Brasília, DF
RESUMO
Foram utilizados 12 cordeiros machos, inteiros, com peso médio de 31,6kg. Os cordeiros foram alimentados com dietas à base de milho e farelo de soja, suplementadas com diferentes fontes de cálcio: calcário calcítico (CC), feno de alfafa (FA), polpa cítrica (PC) e farinha de conchas de ostras (FCO). Os animais receberam injeção de 7,4 MBq do radiofósforo (32P) e após sete dias foram abatidos para posteriores análises. Utilizou-se o delineamento em blocos ao acaso, com quatro tratamentos e três repetições. A taxa de retenção de 32P, as atividades específicas relativas e padronizadas, e a quantidade de P inorgânico nos tecidos foram iguais para todos os tratamentos (P>0,05). A comparação entre os tecidos mostrou que a concentração de P inorgânico e a taxa de retenção de 32P (P<0,01) foram mais altas nos ossos em relação aos tecidos moles (P<0,01). A retenção de 32P apresentou a seguinte ordem decrescente: rins, fígado, coração, músculos e ossos (P<0,01). A reabsorção óssea de P foi maior que a deposição de P, levando ao balanço negativo do elemento para a maioria dos animais. Concluiu-se que as diferentes fontes de cálcio não interferiram na cinética do P e que a quantidade de P ingerida não atendeu às exigências de P dos animais.
Palavras-chave: ruminante, radiofósforo, ossos, tecidos moles, retenção
ABSTRACT
Sixteen growing rams, averaging 31.6kg were studied. They were fed diets based on corn and soya meal, supplemented with different sources of calcium: limestone (CC), alfalfa hay (FA), citrus pulp (PC) and oyster shell meal (OSM). The animals received an injection of 7.4 MBq of radioactive P (32P) and after 7 days they were slaughtered for further analyses. Experimental measurements were analyzed as a completely randomized design with four treatments and three repetitions. The rate of 32P retention, the relative specific activity, the standardized specific activity and the inorganic P in tissue were similar for all treatments (P>0.05). The comparison between tissues showed that the concentration of inorganic P and rate of 32P retention was greater in bone (P<0.01) than in soft tissue. The 32P retention presented the following decreasing order: kidney, liver, heart, muscle and bone (P<0.01). P resorption in bone was higher than P accreted in bone, leading to a negative balance of P in bone for the majority of animals. The means for P accreted in bone were 1.56, 3.29, 2.74 and 2.06g/animal and the means for P resorption in bone were 2.82, 4.51, 4.09 and 3.05g/animal for limestone (CC), alfalfa hay (FA), citrus pulp (PC) and oyster shell meal (OSM), respectively. P resorption values were higher than P accretion in bone leading to a negative balance of P for the majority of animals. It was concluded that the different sources of calcium did not interfere in the kinetics of phosphorus and the amount of P absorbed did not attend P demands of the animals.
Keywords: ruminant, radiophosphorus, bones, soft tissues, retention
INTRODUÇÃO
Atualmente grande parte das pesquisas agropecuárias enfatiza as questões referentes às práticas que visam à sustentabilidade, ajustando viabilidade econômica, melhor desempenho animal e a preocupação com questões ambientais.
O melhor entendimento do metabolismo do P nos ruminantes, conseqüentemente, melhora a eficiência na utilização desse mineral, além de reduzir a excreção do P (Knowlton et al., 2004).
O estudo da cinética do P fornece informações importantes a respeito do metabolismo do P, como, os fluxos de entrada e saída de P dos compartimentos que representam os tecidos moles, ossos e sangue. A injeção de 32P nos animais permite rastrear a mobilização e até mesmo avaliar a velocidade de intercâmbio do P entre os tecidos. A retenção do 32P, por exemplo, representa a substituição do P estável pelo radiotraçador, esclarecendo a magnitude do intercâmbio do elemento no interior da célula com o plasma e fluidos intersticiais (Bueno e Vitti, 1999).
A relação entre o metabolismo do Ca e P ocorre porque eles são regulados por mecanismos fisicoquímicos e biológicos idênticos. Além disso, devido à natureza estática da razão Ca/P encontrada nos ossos, os efeitos do metabolismo do Ca na absorção e reabsorção óssea podem alterar as concentrações de P no sangue (Valk, et al, 2000).
A captação e a mobilização de grandes quantidades do Ca e do P ocorrem lentamente, devido à menor atividade metabólica desses elementos nos ossos.
As trocas bioquímicas de P nos tecidos moles, sangue e ossos são constantes e variam principalmente com a idade e o estado fisiológico dos animais. Nos animais jovens, a mineralização e o crescimento ósseo ocorrem independentes do P dietético, enquanto, nos animais adultos, a utilização de reservas desse mineral é menos eficiente do que a utilização do cálcio, e não parece estar sob o mesmo controle hormonal deste mineral (Underwood, 1999).
A atividade específica representa o intercâmbio do elemento com os fluidos extracelulares e compensa as diferenças em conteúdo de P dos diversos tecidos, tornando a intensidade do metabolismo independente da radioatividade. A atividade específica padronizada, por exemplo, ocorre em função da atividade injetada e da unidade de peso vivo e a atividade específica relativa remove o efeito de diferentes condições fisiológicas e dos processos metabólicos, como a formação de ossos (Smith et al., 1951).
O presente trabalho teve como objetivo avaliar a distribuição, a taxa metabólica e o intercâmbio do P nos tecidos de cordeiros alimentados com dietas contendo diferentes fontes de cálcio.
MATERIAL E MÉTODOS
O experimento foi desenvolvido no laboratório de nutrição animal do Centro de Energia Nuclear na Agricultura da Universidade de São Paulo, em Piracicaba, usando-se 12 cordeiros mestiços da raça Santa Inês, machos inteiros, com idade entre sete e oito meses, e peso médio de 31,6kg.
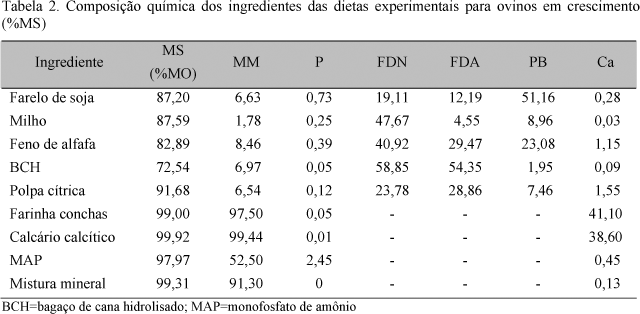
Os animais foram divididos em quatro tratamentos: calcário calcítico (CC), feno de alfafa (FA), polpa cítrica (PC), e farinha de conchas de ostras (FC) (Tab. 1). A dieta basal foi composta de milho, farelo de soja, uréia, bagaço de cana hidrolisado, mistura mineral e MAP (monofosfato de amônio) (Tab. 2).
As dietas foram formuladas para atender às exigências nutricionais de cordeiros com ganhos de peso diários equivalentes a 295g, preconizados pelo NRC (Nutrient..., 1985).
Inicialmente os animais permaneceram 20 dias nas baias para adaptação às dietas sendo, então, alocados em gaiolas metabólicas individuais, onde permaneceram durante sete dias como período de adaptação às gaiolas. Nos três dias que antecederam à injeção de P radioativo, foram coletadas amostras de fezes, urina e sobras para análise e, no dia anterior à injeção, foi feita coleta de sangue de cada animal. Após uma semana nas gaiolas, os animais receberam injeção 7,7 MBq de 32P na jugular direita. Amostras de sangue foram colhidas pela jugular esquerda aos 5 min, e à 1 e às 2, 4 e 6h após a injeção e a cada 24h, durante sete dias. Amostras de fezes, urina e sobras de cada animal foram coletadas a cada 24h após a injeção, sempre pela manhã, antes de a dieta ser oferecida, durante os sete dias.
A solução radioativa foi preparada a partir de uma solução de fosfato de sódio com 32P (Na2PO4), livre de carregador, fornecida pelo Instituto de Pesquisas Energéticas de São Paulo, utilizando-se uma solução salina estéril de NaCl a 0,85%. Para o preparo da solução padrão, foi utilizada uma amostra contendo 0,5ml da solução radioativa, colocada em um balão de um litro, completado com água destilada.
As amostras de plasma e urina foram transferidas para frascos contendo soluções cintiladoras, para posterior detecção da radioatividade através do cintilador líquido (sistema Beckman LS-5000 TA) (Laboratory..., 1979). As fezes foram maceradas, homogeneizadas e pesadas (1g) para determinação de matéria seca a 105ºC, cinzas a 500ºC, seguida da digestão com 10ml de ácido sulfúrico. Um mililitro do material digerido foi transferido para os frascos de cintilação e completado com a solução cintiladora para posterior leitura da radioatividade. Os cálculos do porcentual das atividades específicas (32P/Ptotal) foram feitos de acordo com Lofgreen (1960).
Para os tecidos foram calculadas:
retenção de 32P = (CPM da amostra do tecido/g de MS)/[dose total injetada/(mg de P/g de MS)];
atividade padronizada = [(CPM da amostra do tecido/g de MS)/(mg de P/g de MS)]/(CPM da dose injetada/kg de peso do animal);
atividade específica relativa = atividade específica do tecido/atividade específica do plasma.
A deposição e a reabsorção de P nos ossos e tecidos foram calculadas utilizando-se o modelo matemático proposto por Vitti et al. (2000).
A análise bromatológica das dietas foi realizada seguindo-se as recomendações da AOAC (Official..., 1980). A determinação do P inorgânico foi feita por colorimetria, usando o método vanadato-molibdato (Sarruge e Haag, 1974), e a do cálcio, por espectrometria de absorção atômica (Zagatto et al., 1979).
O conteúdo de P inorgânico do plasma foi determinado através do método de Fiske e Subbarow (1925), e para os tecidos o P inorgânico foi determinado pelo método vanadato-molibdato (Sarruge e Haag, 1974).
O delineamento experimental utilizado foi o inteiramente casualizado, com quatro tratamentos e três repetições. A comparação das médias foi feita utilizando-se o teste Duncan (Statistical..., 2000).
RESULTADOS E DISCUSSÃO
A retenção de 32P foi maior nos ossos (P<0,01) e igual (P>0,05) para os demais tecidos (Tab. 3) que apresentaram a seguinte ordem decrescente de retenção de 32P: músculos, fígado, coração e rins. Foi observada correlação positiva entre o teor de P dos tecidos e a retenção de 32P representada pela equação y = 0,070x 0,32, (R2 = 0,80, P<0,01). A mesma ordem decrescente de retenção de 32P foi observada por Arrington et al. (1963) em bezerros e por Bueno e Vitti (1999), que também observaram similar correlação ao fornecerem quantidade insuficiente, adequada e elevada de P para caprinos. No entanto, Lobão e Crocomo (1974), ao trabalharem com ovinos, e Teixeira et al. (2004) e Lopes et al. (1999), em suínos, encontraram diferentes ordens para as retenções de 32P, sendo que nesses trabalhos os ossos apresentaram valor de retenção menor que os dos tecidos moles. O menor intervalo de tempo entre o abate dos animais e a injeção do radionuclídeo (24h) pode explicar a menor retenção do 32P no trabalho de Lobão e Crocomo (1974), pois a taxa metabólica dos ossos é menor do que a dos demais tecidos, exigindo maior tempo para incorporação do P. No entanto, nos trabalhos com suínos, o intervalo de tempo entre o abate e a injeção do radionuclídeo foi igual ao do presente experimento, sugerindo que a taxa metabólica dos ossos nos suínos é menor que a taxa metabólica dos ossos de ovinos e caprinos, levando às diferenças na retenção de 32P. Segundo Smith et al. (1952), as taxas de incorporação de P nos tecidos dos ovinos são maiores do que nos suínos, demonstrando maiores taxas metabólicas dos tecidos na primeira espécie em relação à segunda.
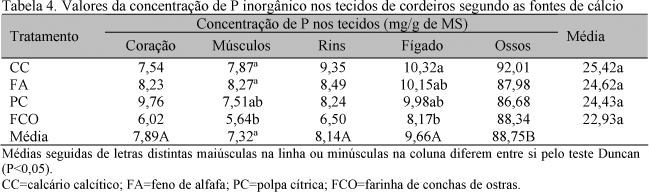
Os valores de P inorgânicos (mgP/gMS) nos tecidos foram iguais para todos os tratamentos (P>0,05) (Tab. 4). No entanto, comparando-se os teores de P nos tecidos, os ossos apresentaram os maiores valores (P<0,01), enquanto músculos, fígado, coração e rins apresentaram valores mais baixos e similares entre si. Esses resultados estão de acordo quanto à ordem com os valores encontrados por Vitti et al. (1992), que trabalharam com ovinos, por Bueno e Vitti (1999), com caprinos, e Lopes et al. (1999), com suínos. A quantidade de P inorgânico encontrada nos tecidos apresentou valor um pouco menor do que a encontrada nos trabalhos de Vitti et al. (1992) e Bueno e Vitti (1999), no entanto esses autores trabalharam com animais adultos, enquanto, no presente trabalho, foram avaliados os tecidos de animais em crescimento.
Teixeira et al. (2004) encontraram diferença significativa para os teores de P nos tecidos moles, observando os maiores valores para fígado e rins, e menores para coração e músculos.
As quantidades médias de P depositadas nos ossos foram: 1,56, 3,29, 2,74, e 2,06g/animal e as quantidades médias de P reabsorvido foram: 2,82, 4,51, 4,09 e 3,05g/animal, respectivamente, para CC, FA, PC e FCO, resultando no balanço negativo de P nos ossos. Estes resultados assemelham-se aos de Vitti et al. (2000), que encontraram balanço negativo de P nos ossos de caprinos em crescimento, alimentados com níveis baixo, médio e alto de P. A alta demanda de P por animais em crescimento poderia explicar os balanços negativos de P nos ossos.
Nos tecidos moles, as quantidades médias de P depositadas foram: 0,69, 0,80, 1,32 e 1,39g/animal, e as quantidades médias de P reabsorvidas foram: 0,36, 0,60, 0,55 e 0,31g/animal para os tratamentos CC, FA, PC e FCO, respectivamente, resultando em balanço positivo de P nos tecidos.
No presente trabalho, a retenção de P foi negativa para os tratamentos CC, FA e PC, -0,48, -0,95 e -1,00g/animal e positiva, mas muito próxima de zero para o tratamento FCO, 0,06g/animal, mostrando que a alta demanda de P e provavelmente a baixa disponibilidade do P podem ter provocado as retenções negativas de P.
As principais fontes de P das dietas eram de origem orgânica (milho e farelo de soja) e proveniente de grãos, que possuem grande proporção de P na forma de fitato, que, mesmo sendo uma fonte de P disponível para ruminantes, pode ter tido sua disponibilidade afetada por alguma característica da dieta que tenha alterado o ambiente ruminal. No entanto, não existem dados neste trabalho que possam confirmar tal suposição.
É possível que o fornecimento do bagaço de cana hidrolisado como principal volumoso tenha alterado o ambiente ruminal, pois é sabido que esse alimento é de baixa qualidade, porém há a necessidade de mais pesquisas para esclarecer essa hipótese.
Williams et al. (1991), estudando o efeito de dietas deficientes em P em novilhas, concluíram que as concentrações de P presentes nos ossos refletem a ingestão de P, enquanto as concentrações de P presentes no fígado, rins, coração e músculos não respondem às diferentes ingestões de P.
Os balanços de P negativo no animal, negativo nos ossos e positivo nos tecidos moles obtidos neste trabalho parecem confirmar tal afirmativa. Embora não se tenha estudado diferentes níveis de ingestão de P, a baixa disponibilidade do elemento pode ter refletido nos balanços negativos de P nos ossos. No entanto, o balanço de P nos tecidos moles manteve-se positivo.
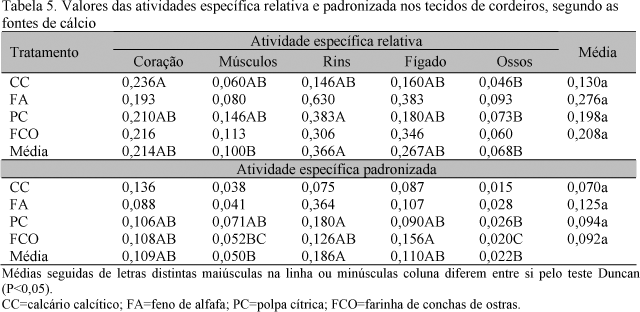
Em relação às médias da atividade específica relativa, os tratamentos apresentaram a seguinte ordem decrescente: rins, fígado, coração, músculos e ossos (P<0,01), sendo essa mesma ordem encontrada para atividade específica padronizada (P<0,01) (Tab. 5). Não foi observada correlação inversa significativa entre o teor de P dos tecidos e a atividade específica padronizada e relativa como observaram Bueno e Vitti (1999).
CONCLUSÕES
As diferentes fontes de cálcio não interferiram na retenção de 32P, concentração de P e atividades específicas relativa e padronizada dos tecidos estudados, mostrando que a cinética do P no tecido não foi influenciada pelas diferentes fontes de cálcio.
Recebido em 3 de maio de 2005
Aceito em 1 de dezembro de 2006
Apoio: FAPESP
E-mail: raquelsd@cena.usp.br
- ARRINGTON, L.R.; OUTLER, J.C.; AMMERMAN, C.B. et al. Absorption, retention and tissue deposition of labeled inorganic phosphates by cattle. J. Anim. Sci., v.22, p.940-942, 1963.
- BUENO, M.S.; VITTI, D.M.S.S. Níveis de P para caprinos: perda endógena fecal e exigência líquida para mantença. Pesq. Agrop. Bras, v.34, p.675-681, 1999.
- FISKE, C.H.; SUBBAROW, Y. The colorimetric determination of phosphorus. J. Biol. Chem., v.66, p.375-400, 1925.
- KNOWLTON, K.F.; RADCLIFFE, J.S.; NOVAK, C.L. et al. Animal management to reduce phosphorus losses to the environment. J. Anim. Sci, v.82, suppl., p.E173-E195, 2004.
- LABORATORY training manual on the use of nuclear techniques in animal research. Vienna: IAEA, 79, (Technical Report Series, 193), 1979. 299p.
- LOBÃO, A.O.; CROCOMO, O.J. Retenção de P radioativo (P32) em tecidos de ovinos. Bol. Ind. Anim, v.31, p.261 291, 1974.
- LOFGREEN, G.P. The availability of the phosphorus in dicalcium phosphate, bone, meal, soft phosphate and calcium phytates for mature wethers. J. Nutr, v.70, p.58-62, 1960.
- LOPES, J.B.; VITTI, D.M.S.S.; FIGUERÊDO, A.V. et al. Cinética do P em tecidos de suínos em crescimento. Rev. Bras. Zootec, v.28, p.779784, 1999.
- NUTRIENT requirement of sheep. 6.ed. Washington: Academic, 1985. 99p.
- OFFICIAL methods of the analysis of the Association of Official Analytical Chemists. 13.ed. Washington: AOAC 1980. 1018p.
- SARRUGE, J.R.; HAAG, H.P. Análises químicas em plantas Piracicaba: ESALQ/USP, 1974. 56p.
- SMITH, A.H.; KLEIBER, M.; BLACK, A.L. et al. Distribution of intravenously injected radioactive phosphorus (P32) among swine tissue. J. Anim. Sci, v.10, p.893-901, 1951.
- SMITH, A.H.; KLEIBER, M.; BLACK, A.L. et al. Distribution of intravenously injected radioactive phosphorus (P32) among sheep tissue. J. Anim. Sci, v.11, p.638645, 1952.
- STATISTICAL analysis system. The SAS system for windows. Release 8.01. Cary: SAS Institute, 2000.
- TEIXEIRA, A.O.; LOPES, D.C.; LOPES, J.B et al. Cinética do radiofósforo em tecidos de suínos em crescimento alimentados com dietas contendo diferentes fontes de P. Rev. Bras. Zootec, v.33, p.1238-1245, 2004.
- UNDERWOOD, E.J.; SUTTLE, N.F. The mineral nutrition of livestock 3.ed. Wallingford: CAB International, 1999.
- VALK, H.; METCALF, J.A.; WITHERS, P.J.A. Prospects for minimizing phosphorus excretion in ruminants by dietary manipulation. J. Environ. Qual, v.29, p.28-36, 2000.
- VITTI, D.M.S.S.; ABDALLA, A.L.; MEIRELLES, C.F. Cinética do P em ovinos suplementados com diferentes fontes fosfatadas através da técnica de diluição isotópica. Arq. Bras. Med. Vet. Zootec, v.44, p.227-233, 1992.
- VITTI, D.M.S.S.; KEBREAB, E.; LOPES, J.B. et al. A kinetic model of phosphorus metabolism in growing goats. J. Anim. Sci, v.78, p.2706-2712, 2000.
- WILLIAMS, S.N.; McDOWELL, L.R.; WARNICK, A.C. et al. Phosphorus concentrations in blood, milk, feces, bone and selected fluids and tissues of growing heifers as affected by dietary phosphorus. Livest. Res. Rural Dev, v.3, p.67-80, 1991.
- ZAGATTO, E.A.G.; KRUG, F.J.; BERGAMIM FILHO, H. et al. Margin zones in flow injection analysis. Part 2. Determination of calcium, magnesium and potassium in plant material by flow injection atomic and flame emission spectrometry. Anal. Chem. Acta, v.104, p.279-284, 1979.
Datas de Publicação
-
Publicação nesta coleção
02 Maio 2007 -
Data do Fascículo
Fev 2007
Histórico
-
Aceito
01 Dez 2006 -
Recebido
03 Maio 2005