Resumos
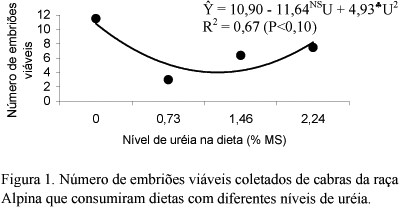
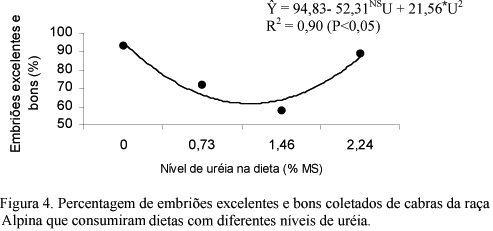
Utilizaram-se 22 cabras da raça Alpina, distribuídas aleatoriamente em quatro tratamentos (T): as cabras do T1 (n=5) formaram o grupo-controle; as do T2 (n=7) receberam 0,73% de uréia na matéria seca da dieta; as do T3 (n=4) receberam 1,46% de uréia; e as do T4 (n=6), 2,24% de uréia. As cabras foram superovuladas e os embriões, coletados entre sete e oito dias após a primeira monta, foram avaliados quanto à qualidade e ao estádio de desenvolvimento. Amostras de sangue para dosagem dos teores de uréia e glicose foram coletadas nos dias do estro e da coleta de embriões. Houve efeito linear crescente do nível de uréia nas dietas sobre o consumo de MS (kg/dia) e de proteína bruta (kg/dia). O peso das cabras não diferiu (P>0,05) entre os tratamentos nem entre as semanas experimentais. Dezoito cabras (81,8%) manifestaram estro após a sincronização. A duração do estro e o intervalo da remoção da esponja ao início do estro não foram influenciados (P>0,05) pelos tratamentos. Quatorze cabras (77,8%) responderam à superovulação. O número de estruturas e de embriões coletados não diferiu (P>0,05) entre os tratamentos. O número (Y= 10,90 - 11,64NS U + 4,93<FONT FACE=Symbol>§</FONT>U²; R² = 0,67; P<0,10) e a percentagem (Y= 94,08 - 39,59NS U + 18,16<FONT FACE=Symbol>ª</FONT>U²; R² = 0,94; P<0,07) de embriões viáveis, e o número (Y= 10,83- 12,18NS U + 5,02mU²; R² = 0,78; P<0,08) e a percentagem (Y= 94,83- 52,31NS U + 21,56*U²; R² = 0,90; P<0,05) de embriões excelentes e bons apresentaram comportamento quadrático em função do nível de uréia nas dietas. Os teores de uréia e glicose no plasma não foram influenciados (P>0,05) pelos tratamentos. A uréia pode ser fornecida no nível de 2,24% na MS da dieta de cabras não lactantes.
cabra; embrião; glicose; uréia
Twenty-two Alpine goats were allocated at random into four treatments: 0.0% (T1 - control, n=5); 0.73% (T2, n=7); 1.46% (T3, n=4) or 2.24% (T4, n=6) of urea in the dry matter (DM) of the diet. Embryos collected from 7 to 8 days after mating of superovulated goat were evaluated by quality and development stage. Blood samples for urea and glucose analyses were collected at estrus and at embryos collection day. The DM (kg/day) and crude protein (kg/day) intake increased linearly in function of dietary urea level. Goat body weights were not affected by treatments out experimental weeks (P>0.05). Eighteen goats (81.8%) came in estrus after the synchronization. The estrus length and the interval from sponge removal to the beginning of estrus were not affected (P>0.05) by treatments. Fourteen goats (77.8%) were responsive to superovulation protocol. The levels of urea (treatments) did not affect structures and embryo numbers (P>0.05). Number (Y= 10.90 - 11.64NS U + 4.93<FONT FACE=Symbol>§</FONT>U²; R²= 0.67; P<0.10) and percentage (Y= 94.08 - 39.59NS U + 18.16<FONT FACE=Symbol>ª</FONT>U²; R²= 0.94; P<0.07) of viable embryos, and number (Y= 10.83- 12.18NS U + 5.02mU²; R²= 0.78; P<0.08) and the percentage (Y= 94.83- 52.31NS U + 21.56*U²; R²= 0.90; P<0.05) of excellent and good embryos were quadraticaly effected by urea dietary level. The treatments did not affect plasma urea and glucose levels (P>0.05). Diets for no nursing goats can be supplied by urea at 2.24% of DM.
goat; embryo; glucose; urea
ZOOTECNIA E TECNOLOGIA E INSPEÇÃO DE PRODUTOS DE ORIGEM ANIMAL
Efeito do nível de uréia na dieta sobre o desempenho, a qualidade e o estádio de desenvolvimento embrionário em cabras Alpinas
Effect of the dietary urea level on performance, quality and embryonic development stage on Alpine goats
N.G. AlvesI; C.A.A. TorresII; M.T. RodriguesII; P.R. CeconIII; A.D.F. SantosIV; L.G.B. SiqueiraII; L.L. BitencourtI; E.A. MoraesII
IDepartamento de Zootecnia - UFLA Caixa Postal 3037 37200-000 Lavras, MG
IIDepartamento de Zootecnia - UFV Viçosa, MG
IIIDepartamento de Estatística - UFV Viçosa, MG
IVFaculdade Pio Décimo Aracaju, SE
RESUMO
Utilizaram-se 22 cabras da raça Alpina, distribuídas aleatoriamente em quatro tratamentos (T): as cabras do T1 (n=5) formaram o grupo-controle; as do T2 (n=7) receberam 0,73% de uréia na matéria seca da dieta; as do T3 (n=4) receberam 1,46% de uréia; e as do T4 (n=6), 2,24% de uréia. As cabras foram superovuladas e os embriões, coletados entre sete e oito dias após a primeira monta, foram avaliados quanto à qualidade e ao estádio de desenvolvimento. Amostras de sangue para dosagem dos teores de uréia e glicose foram coletadas nos dias do estro e da coleta de embriões. Houve efeito linear crescente do nível de uréia nas dietas sobre o consumo de MS (kg/dia) e de proteína bruta (kg/dia). O peso das cabras não diferiu (P>0,05) entre os tratamentos nem entre as semanas experimentais. Dezoito cabras (81,8%) manifestaram estro após a sincronização. A duração do estro e o intervalo da remoção da esponja ao início do estro não foram influenciados (P>0,05) pelos tratamentos. Quatorze cabras (77,8%) responderam à superovulação. O número de estruturas e de embriões coletados não diferiu (P>0,05) entre os tratamentos. O número (Y= 10,90 - 11,64NSU + 4,93§U2; R2 = 0,67; P<0,10) e a percentagem (Y= 94,08 - 39,59NSU + 18,16ªU2; R2 = 0,94; P<0,07) de embriões viáveis, e o número (Y= 10,83- 12,18NSU + 5,02mU2; R2 = 0,78; P<0,08) e a percentagem (Y= 94,83- 52,31NSU + 21,56*U2; R2 = 0,90; P<0,05) de embriões excelentes e bons apresentaram comportamento quadrático em função do nível de uréia nas dietas. Os teores de uréia e glicose no plasma não foram influenciados (P>0,05) pelos tratamentos. A uréia pode ser fornecida no nível de 2,24% na MS da dieta de cabras não lactantes.
Palavras-chave: cabra, embrião, glicose, uréia
ABSTRACT
Twenty-two Alpine goats were allocated at random into four treatments: 0.0% (T1 - control, n=5); 0.73% (T2, n=7); 1.46% (T3, n=4) or 2.24% (T4, n=6) of urea in the dry matter (DM) of the diet. Embryos collected from 7 to 8 days after mating of superovulated goat were evaluated by quality and development stage. Blood samples for urea and glucose analyses were collected at estrus and at embryos collection day. The DM (kg/day) and crude protein (kg/day) intake increased linearly in function of dietary urea level. Goat body weights were not affected by treatments out experimental weeks (P>0.05). Eighteen goats (81.8%) came in estrus after the synchronization. The estrus length and the interval from sponge removal to the beginning of estrus were not affected (P>0.05) by treatments. Fourteen goats (77.8%) were responsive to superovulation protocol. The levels of urea (treatments) did not affect structures and embryo numbers (P>0.05). Number (Y= 10.90 11.64NSU + 4.93§U2; R2= 0.67; P<0.10) and percentage (Y= 94.08 39.59NSU + 18.16ªU2; R2= 0.94; P<0.07) of viable embryos, and number (Y= 10.83- 12.18NSU + 5.02mU2; R2= 0.78; P<0.08) and the percentage (Y= 94.83- 52.31NSU + 21.56*U2; R2= 0.90; P<0.05) of excellent and good embryos were quadraticaly effected by urea dietary level. The treatments did not affect plasma urea and glucose levels (P>0.05). Diets for no nursing goats can be supplied by urea at 2.24% of DM.
Keywords: goat, embryo, glucose, urea
INTRODUÇÃO
Estudos têm sugerido que o fornecimento de dietas com elevado nível de proteína bruta (PB) e de proteína degradada no rúmen (PDR) influencia a reprodução, reduzindo a taxa de concepção ao primeiro serviço (Canfield et al., 1990) e resultando em maior número de dias abertos e de serviços por concepção (Jordan e Swanson, 1979) em vacas.
O excesso de PB ou de PDR e o fornecimento de uréia na dieta foram associados ao elevado teor sistêmico de N-uréico em ovelhas (Bishonga et al., 1994) e novilhas (Gath et al., 1999), à alteração da composição iônica da secreção uterina (Jordan et al., 1983) e à redução do pH uterino (Elrod et al., 1993; Elrod e Butler, 1993) em vacas. Elevação dos teores de uréia e amônia no fluido útero-tubárico de ovelhas (McEvoy et al., 1997) e de amônia no fluido folicular de novilhas (Sinclair et al., 2000) também foram observados.
Falhas de fertilização e degeneração de embriões bovinos foram relacionadas ao excesso de PDR na dieta (Blanchard et al., 1990). Em ovelhas, o fornecimento de uréia na dieta ocasionou redução da viabilidade (Bishonga et al., 1994; McEvoy et al., 1997) e do desenvolvimento embrionário (Fahey et al., 2001; Papadopoulos et al., 2001). Sinclair et al. (2000) verificaram que ovócitos coletados de novilhas com elevado teor de amônia plasmática resultaram em reduzido desenvolvimento embrionário in vitro. Da mesma forma, a proporção de ovócitos que se desenvolveram em blastocistos in vitro foi reduzida quando os ovócitos foram maturados em meios com 7,5 mmols/l de uréia (Ocon e Hansen, 2003).
Este estudo teve como objetivo verificar o efeito de níveis crescentes de uréia na MS da dieta sobre o desempenho, o número de estruturas e de embriões coletados, a qualidade e o estádio de desenvolvimento embrionário e os teores de uréia e glicose no plasma de cabras Alpinas.
MATERIAL E MÉTODOS
O experimento, realizado nos meses de junho e julho de 2003, utilizou 22 cabras da raça Alpina, nulíparas, primíparas e multíparas, não-lactantes, com média de idade de 40,55±18,48 meses, distribuídas aleatoriamente em quatro tratamentos (T). As cabras do T1 (n=5) formaram o grupo-controle; as do T2 (n=7), receberam 0,73% de uréia na MS da dieta; as do T3 (n=4), 1,46% de uréia; e as do T4 (n=6), 2,24% de uréia.
As dietas à base de silagem de milho e concentrado (Tab. 1) foram calculadas para serem isoenergéticas (1,52 Mcal/kg MS no T1 e 1,54 Mcal/kg MS no T2, T3 e T4) e isoprotéicas (14% PB na MS) e conterem 28,8%; 39,0%; 49,5%; e 60,8% da proteína total na forma de nitrogênio não-protéico (NNP) nos tratamentos T1, T2, T3 e T4, respectivamente. As percentagens de NNP advindas da uréia em relação à proteína total da dieta foram 0%, 13%, 26% e 40% nos tratamentos T1, T2, T3 e T4, respectivamente.
As cabras foram adaptadas de forma gradual às rações com uréia, utilizando-se 0,73% de uréia na primeira semana no T2; 0,73% e 1,46% de uréia na primeira e segunda semanas, respectivamente, no T3; e 0,73%; 1,46%; e 2,24% de uréia na primeira, segunda e terceira semanas, respectivamente, no T4. Após o período de adaptação, as dietas foram fornecidas por 81 dias antes do início do experimento.
As cabras de um mesmo tratamento foram mantidas em baia coletiva. O alimento concentrado foi fornecido sobre a silagem, duas vezes ao dia, sendo a quantidade total de alimento oferecido ajustada diariamente para permitir, no mínimo, 5% de sobras. Água e sal mineral foram fornecidos ad libitum. As sobras de cada baia foram retiradas antes do fornecimento diário do alimento e pesadas, para determinação do consumo. Amostras da silagem fornecida e das sobras foram coletadas diariamente, acondicionadas em sacos plásticos e armazenadas a 20ºC. Amostras compostas de cada três semanas para a silagem e de cada nove semanas para as sobras do T1, T2, T3 e T4 e amostras do fubá de milho e do farelo de soja utilizados tiveram a composição bromatológica determinada segundo os procedimentos descritos por Silva e Queiroz (2002). A composição bromatológica da silagem de milho, do fubá de milho e do farelo de soja utilizados é apresentada na Tab. 2.
As cabras foram pesadas uma vez por semana, no período da manhã, antes do fornecimento da alimentação.
As cabras foram sincronizadas e superovuladas segundo os protocolos descritos por Salles et al. (2002) com modificações. O estro foi sincronizado com a inserção de esponja intravaginal impregnada com 60mg de acetato de medroxiprogesterona1 1 Progespon ® - Schering-Plough Cotia, Brasil. por 11 dias. A superovulação foi realizada com 200mg de NIH-FSH-P12 2 Foltropin-V ® - Vetrepharm - Canadá. , fracionados em seis doses decrescentes, com intervalo de 12 horas entre aplicações e a partir de 48 horas antes da remoção da esponja. Simultaneamente à primeira aplicação de FSH foram administrados 125µg de cloprostenol sódico3 3 Ciosin ® Coopers Schering-Plough Cotia, Brasil. , por via intramuscular (IM). Setenta e duas horas após a remoção da esponja iniciou-se o tratamento antiluteolítico com flunixin meglumine4 4 Banamine ®, Schering Plough Cotia, Brasil. , na dose de 1,1mg/kg/dia, durante três dias consecutivos.
A partir de 12 horas após a retirada das esponjas, as cabras foram observadas quanto à manifestação de estro, duas vezes ao dia, às 7 e às 19 horas, por, no mínimo, 15 minutos cada, com auxílio de rufiões. As cabras em estro foram cobertas por bodes de fertilidade comprovada, a cada 12 horas, até a não aceitação da monta.
Os embriões foram coletados pela técnica transcervical em circuito fechado, entre o sétimo e o oitavo dia do ciclo estral (estro = dia zero). Aproximadamente 24 horas antes do início da coleta de embriões, as cabras receberam uma aplicação de 125mg de cloprostenol sódico3, por via IM.
A cérvice uterina foi visualizada com o auxílio de um espéculo vaginal e uma fonte de luz e tracionada com duas pinças de Pozzi em direção ao vestíbulo da vagina. Uma sonda nasogástrica nº 10 ou 12 foi introduzida através da cérvix e em direção ao lúmen de um dos cornos uterinos. A extremidade livre da sonda foi conectada ao equipo, ao qual foram acoplados o filtro de coleta de embriões e uma seringa de 50ml, para infusão do meio de lavagem - solução-tampão salina fosfato modificada por Whittingham (1971), acrescida de 50µg/ml de gentamicina e 1,0% de soro fetal bovino -, a 37ºC. O meio de coleta de embriões foi infundido até que houvesse resistência do corno uterino, aproximadamente 20ml, quando, então, a via de infusão foi fechada e a de comunicação com o filtro foi aberta. Foram utilizados, em média, 200ml de meio de coleta por corno uterino.
Os embriões coletados foram avaliados ao estereomicroscópio quanto à qualidade, I excelente e V degenerado, e ao estádio de desenvolvimento (Stringfellow e Seidel, 1999).
Foram coletadas amostras de sangue por punção da veia jugular, em tubos de vidro de coleta a vácuo com anticoagulante EDTA para dosagem de uréia e anticoagulante fluoreto para dosagem de glicose, no dia do estro e no da coleta de embriões. As amostras coletadas antes e aproximadamente quatro horas após a alimentação foram centrifugadas a 1.000 x g, durante 15 minutos, para a obtenção de plasma. Este foi armazenado a 20ºC até a dosagem dos teores de uréia e glicose, por método colorimétrico-enzimático, utilizando kits comerciais5 5 Doles Reagentes e Equipamentos para Laboratórios Ltda, Brasil .
Os dados foram analisados pelo programa SAEG (Sistema..., 1997). Os dados relativos ao consumo médio individual de MS e PB, duração do estro, intervalo da remoção da esponja ao início do estro, números de estruturas, de embriões, de embriões viáveis e de embriões excelentes e bons, percentagens de embriões viáveis e de embriões excelentes e bons, e concentrações de uréia e glicose no plasma foram submetidos às análises de variância e de regressão. As variáveis relativas ao estádio de desenvolvimento embrionário foram apresentadas na forma de estatística descritiva. Os dados relativos ao peso dos animais foram analisados por meio da metodologia de superfície de resposta.
RESULTADOS E DISCUSSÃO
Houve efeito linear crescente da adição de uréia às dietas sobre o consumo médio individual de MS e PB, expressos em kg/dia (Tab. 3). Segundo o NRC (Nutrient..., 1981), a adição de uréia à dieta dos caprinos não deve exceder 1/3 da PB total ou mais que 50,0% da PB do concentrado. Neste estudo, o fornecimento de até 40,0% da proteína total na forma de uréia resultou em aumento linear no consumo de MS. Da mesma forma, Fernandez et al. (1997) relataram que a uréia pode ser fornecida em quantidades acima das preconizadas pelo NRC (Nutrient..., 1981) sem que o consumo de MS seja reduzido.
Os diferentes níveis de uréia nas dietas não influenciaram o peso vivo das cabras ( = 56,15; P>0,05), semelhante entre tratamentos e constante ao longo das sete semanas experimentais, mostrando que a quantidade de alimento oferecida foi suficiente para atender às exigências de mantença.
= 56,15; P>0,05), semelhante entre tratamentos e constante ao longo das sete semanas experimentais, mostrando que a quantidade de alimento oferecida foi suficiente para atender às exigências de mantença.
Dezoito cabras (81,8%) manifestaram estro após a sincronização. A duração do estro e o intervalo da remoção da esponja ao início do estro não foram influenciados (P>0,05) pelos tratamentos (Tab. 4), sendo representados pelas equações = 33,23 e = 40,71, respectivamente. Fahey et al. (2001) não verificaram diferença no intervalo da remoção do dispositivo intravaginal impregnado com progesterona e o início do estro entre as ovelhas suplementadas com 50g de uréia/dia e as ovelhas do grupo-controle.
As dietas experimentais foram fornecidas por 110,60±6,19; 115,33±6,25; 108,50±5,45; e 110,67±10,60 dias antes da primeira coleta de embriões nos T1, T2, T3 e T4, respectivamente.
Das 18 cabras que manifestaram estro e foram submetidas à coleta de embriões, uma de cada em T1, T2 e T3 (16,7%) não produziu estrutura, e outra, em T4 (5,6%) produziu um ovócito e foram excluídas das análises subseqüentes. Quatorze cabras (77,8%) responderam à superovulação com produção de 135 estruturas, sendo 119 (88,2%) embriões, nove ovócitos (6,7%) e sete zonas pelúcidas (5,2%).
Cinqüenta (42,0%), 20 (16,8%), 32 (26,9%) e 17 (14,3%) embriões foram coletados das cabras do T1, T2, T3 e T4, respectivamente. Embora mais embriões tenham sido coletados das cabras do T1 e T3, os números de estruturas e de embriões não foram influenciados (P>0,05) pelos tratamentos (Tab. 5), sendo representados pelas equações = 9,64 e = 8,50, respectivamente.
Em novilhas, Gath et al. (1999) não verificaram efeito do fornecimento de uma dieta-controle e dietas de alta ou baixa energia suplementadas com 250g de uréia sobre o número de embriões coletados. Em ovelhas, McEvoy et al. (1997) observaram que o número de embriões coletados não diferiu entre aquelas do grupo-controle e as suplementadas com 15g ou 30g de uréia/dia. Fahey et al. (2001) e Papadopoulos et al. (2001) também não verificaram efeito do fornecimento de 50g de uréia/ovelha/dia sobre o número de embriões coletados por doadora.
Ao utilizarem a mesma técnica de coleta deste estudo, em cabras da raça Bôer, Gusmão et al. (2003) verificaram média de 8,4 estruturas/doadora e, em cabras de raça não definida, Salles (2003) citou média de 7,8 estruturas/doadora. Na raça Saanen, Lima-Verde et al. (2003) coletaram, em média, 6,3 embriões/doadora. As médias verificadas neste estudo são pouco superiores às mencionadas na literatura, o que pode estar relacionado ao fato de que apenas as cabras que responderam à superovulação foram incluídas nas análises estatísticas.
No T1, T2, T3 e T4, 47 (94%), 15 (75%), 23 (72%) e 16 (94%) foram classificados como viáveis (excelentes, bons e regulares) e 46 (92%), 15 (75%), 19 (59%) e 15 (88%) foram classificados como embriões excelentes e bons, respectivamente. Essas variáveis apresentaram comportamento quadrático em função do nível de uréia na dieta. Conforme a derivação das equações, o menor número e a menor percentagem de embriões viáveis, quatro embriões e 72,5%, ocorreram com o consumo de 1,2% e 1,1% de uréia na MS da dieta, respectivamente (Fig. 1 e 2). O menor número e a menor percentagem de embriões excelentes e bons, 3,4 embriões e 63,0%, coincidiram com o consumo de 1,2% de uréia na MS da dieta (Fig. 3 e 4).
Em ovelhas, McEvoy et al. (1997) observaram que o fornecimento de 30g de uréia/dia reduziu a percentagem de embriões viáveis, comparado ao fornecimento de 15g de uréia/dia. Segundo Dawuda et al. (2002), o efeito prejudicial da utilização de uréia na dieta sobre a qualidade embrionária pode estar relacionado ao período de seu fornecimento, visto que, a suplementação com 250g de uréia/vaca/dia, durante sete dias, reduziu a percentagem de embriões viáveis, comparado à suplementação por 17 dias.
Ocon e Hansen (2003) verificaram que a maturação de ovócitos bovinos em meios contendo 0; 5; 7,5; e 10mmols/l de uréia, equivalente a 0; 14; 21; e 28mg/dl de N-uréico plasmático, respectivamente, não influenciou a taxa de clivagem no terceiro dia após a fertilização. Todavia, reduzida proporção de ovócitos maturados em meios com 7,5mmols/l de uréia desenvolveu-se em blastocistos no oitavo dia de cultivo in vitro. Segundo os autores, a alta concentração de uréia (>7,5mmols/l) pode desencadear algum mecanismo compensatório no ovócito que aumenta sua resistência à toxicidade, como, por exemplo, alterações no transporte de uréia pela membrana, inibindo a sua entrada na célula ou aumentando sua remoção do citoplasma. Embora tenha sido demonstrado que o transporte de uréia mediado por carreadores ocorre em outros tipos celulares (Hisanaga et al., 1991, citados por Ocon e Hansen, 2003), o sistema de transporte de uréia no ovócito, bem como sua regulação, ainda não são conhecidos.
Em relação ao estádio de desenvolvimento, foram coletados 1 (0,84%) embrião de quatro células, 25 (21,0%) mórulas, 18 (15,1%) blastocistos iniciais, 22 (18,5%) blastocistos, 28 (23,5%) blastocistos expandidos, 13 (10,9%) blastocistos eclodidos e 12 (10,1%) embriões degenerados. O número e a percentagem de embriões coletados em cada estádio de desenvolvimento, em cada tratamento, são apresentados na Tab. 6.
Segundo Baril et al. (1993), citados por Gordon (1997), no sétimo dia após o início do estro, são coletados embriões principalmente no estádio de mórula compacta a blastocisto expandido e no oitavo dia podem ser coletados embriões nos estádios de blastocistos expandidos e eclodidos. Além disso, pode haver variação no estádio de desenvolvimento embrionário entre cabras submetidas à coleta no mesmo intervalo após o estro, bem como entre os embriões coletados de uma mesma cabra.
A alta percentagem (53%) de embriões no estádio de blastocisto eclodido, coletados das cabras do T4, pode ser sugestivo de que o excesso de uréia na alimentação teve algum efeito estimulatório sobre o desenvolvimento embrionário. McEvoy et al. (1997) observaram que os embriões coletados de ovelhas que receberam 30g de uréia/kg de alimento estavam em estado mais avançado de desenvolvimento do que os coletados de ovelhas que consumiram 2,5g de uréia/kg de alimento.
Os teores de uréia e glicose no plasma, no dia do estro e no dia da coleta de embriões, antes da alimentação e quatro horas após, em função do nível de uréia nas dietas, são apresentados na Tab. 7.
O teor de uréia no plasma no dia do estro e no da coleta de embriões, antes da alimentação e quatro horas após, não foi influenciado (P>0,05) pelos tratamentos. Oliveira (2001), ao utilizar as mesmas fontes de PDR (farelo de soja) e NNP (uréia) deste estudo e níveis semelhantes de uréia (0%; 0,7%; 1,4%; e 2,1% na MS da dieta), também não verificou diferença no teor de uréia no plasma de vacas lactantes, em função da dieta oferecida, o que pode estar relacionado à alta degradabilidade ruminal da fonte protéica utilizada nos concentrados. Outra possibilidade é que os animais tenham se adaptado ao alto nível de uréia na dieta, devido às alterações na atividade dos microrganismos ruminais e também à maior concentração das enzimas hepáticas relacionadas ao ciclo da uréia (Payne e Morris, 1969, citados por Jordan et al., 1983). Embora o comportamento ingestivo não tenha sido avaliado, é provável que as cabras que receberam uréia nas dietas aumentaram sua freqüência de alimentação, ingerindo pequenas porções do alimento por vez, o que resultou no teor plasmático de uréia semelhante ao das cabras do grupo-controle. Por outro lado, a composição percentual das dietas experimentais pode estar relacionada à similaridade observada no teor plasmático de uréia entre cabras dos diferentes tratamentos. O amido proveniente do fubá de milho, ausente na dieta oferecida às cabras do T1, mas presente em quantidades crescentes nas dietas oferecidas à cabras dos T2 (10,1%), T3 (16,7%) e T4 (23,6%), serviu como fonte de carboidratos prontamente fermentáveis, permitindo a conversão adequada do NNP proveniente da uréia em proteína microbiana.
Em estudos com novilhas (Gath et al., 1999; Kenny et al., 2002), com vacas lactantes (Dawuda et al., 2002), com ovelhas (McEvoy et al., 1997; Fahey et al., 2001) e com cabras (Fernandez et al., 1997), o fornecimento de uréia nas dietas resultou em aumento do teor sistêmico desse metabólito.
O teor de glicose no plasma antes da alimentação, no dia do estro, aumentou linearmente com o aumento do nível de uréia nas dietas ( = 71,82 + 4,38U; R2 = 0,59; P<0,05). No entanto, o teor desse metabólito após a alimentação não diferiu (P>0,05) entre os tratamentos. No dia da coleta de embriões, o teor de glicose plasmática, antes da alimentação e quatro horas após, não diferiu (P>0,05) entre os animais dos tratamentos. Também Fernandez et al. (1997) não verificaram diferenças nos teores de glicose no plasma de cabras Alpinas suplementadas com 1,4%; 1,8%; e 2,9% de uréia na MS da dieta. Em novilhas, Kenny et al. (2002) relataram que o teor sistêmico de glicose não foi influenciado pelo consumo de uréia (0 ou 240g/dia), assim como não foi relacionada à sobrevivência embrionária no 40º dia de gestação. Da mesma forma, neste estudo o teor plasmático de glicose não apresentou correlação (P>0,05) com o número de estruturas e de embriões coletados ou com a qualidade embrionária.
CONCLUSÕES
A uréia pode ser fornecida no nível de 2,24% na MS da dieta de cabras não lactantes, sem que haja redução no desempenho dos animais, no número e na qualidade dos embriões coletados.
Recebido em 3 de novembro de 2005
Aceito em 10 de abril de 2007
E-mail: nadja@ufla.br
- BARIL, G.; BREBION, P.; CHESNE, P. (Eds). Manuel de formation pratique pour la transplantation embryonnaire chez la brebis et la chevre Roma: FAO (Publication 115), 1993. 183p.
- BARIL, G.; CASAMITJANA, P.; PERRIN, J. et al. Embryo production, freezing and transfer in Angora, Alpine and Saanen goats. In: MEETING EUROPEAN EMBRYO TRANSFER ASSOCIATION, 4., 1988. Lion. Proceeding… Lion, 1988. p.67-93.
- BISHONGA, C.; ROBINSON, J.J.; McEVOY, T.G. et al. The effects of excess rumen degradable protein in ewes on ovulation rate, fertilization and embryo survival in vivo and during in vitro culture. Anim. Prod., v.58, p.447, 1994. (Abstract).
- BLANCHARD, T.; FERGUSON, J.; LOVE, L. et al. Effect of dietary crude-protein type on fertilization and embryo quality in dairy cattle. Am. J. Vet. Res., v.51, p.905-908, 1990.
- CANFIELD, R.W.; SNIFFEN, C.J.; BUTLER, W.R. Effects of excess degradable protein on postpartum reproduction and energy balance in dairy cattle. J. Dairy Sci., v.73, p.2342-2349, 1990.
- DAWUDA, P.M.; SCARAMUZZI, R.J.; LEESE, H.J. et al. Effect of timing of urea feeding on the yield and quality of embryos in lactating dairy cows. Theriogenology, v.58, p.1443-1455, 2002.
- ELROD, C.C.; BUTLER, W.R. Reduction of fertility and alteration of uterine pH in heifers fed excess ruminally degradable protein. J. Anim. Sci., v.71, p.694-701, 1993.
- ELROD, C.C.; VAN AMBURGH, M.; BUTLER, W.R. Alterations of pH in response to increased dietary protein in cattle are unique to the uterus. J. Anim. Sci., v.71, p.702-706, 1993.
- FAHEY, J.; BOLAND, M.P.; O'CALLAGHAN, D. The effects of dietary urea on embryo development in superovulated donor ewes and on early embryo survival and development in recipient ewes. Anim. Sci., v.72, p.395-400, 2001.
- FERNANDEZ, J.M.; SAHLU, T.; LU, C.D. et al. Production and metabolic aspects of nonprotein nitrogen incorporation in lactation rations of dairy goats. Small Rum. Res., v.26, p.105-117, 1997.
- GATH, V.; LONERGAN, P.; BOLAND, M.P. et al. Effects of diet type on establishment of pregnancy and embryo development in beef heifers. Theriogenology, v.51, p.224, 1999 (Abstract).
- GORDON, I. Embryo transfer and associated techniques in goats. In: ____ Controlled reproduction in sheep and goats Cambridge, UK: University Press, 1997, v.2, p.416-439.
- GUSMÃO, A.L.; MOURA, J.C.A.; CHALHOUB, M. et al. Colheita, avaliação e criopreservação de embriões caprinos. Rev. Bras. Reprod. Anim., v.27, p.115-120, 2003.
- HISANAGA, H.; IIOKA, H.; MORIYAMA, I. et al. The mechanism of human placental urea transport: a study using placental brusch border (microvillous) membrane vesicles. Asia Oceania J. Obstet. Gynaecol., v.17, p.67-72, 1991.
- JORDAN, E.R.; CHAPMAN, T.E.; HOLTAN, D.W. et al. Relationship of dietary crude protein to composition of uterine secretions and blood in high-producing postpartum dairy cows. J. Dairy Sci., v.66, p.1854-1862, 1983.
- JORDAN, E.R.; SWANSON, L. V. Effect of crude protein on reproductive efficiency, serum total protein, and albumin in the high-producing dairy cows. J. Dairy Sci., v.62, p.58-63, 1979.
- KENNY, D.A.; BOLAND, M.P.; DISKIN, M.G. et al. Effect of rumen degradable protein with or without fermentable carbohydrate supplementation on blood metabolites and embryo survival in cattle. Anim. Sci., v.74, p.529-537, 2002.
- LIMA-VERDE, J.B.; LOPES JÚNIOR, E.S.; TEIXEIRA, D.I.A. et al. Colheita de embriões pela técnica trans-cervical em cabras da raça Saanen criadas nos trópicos. Rev. Bras. Reprod. Anim., v.27, p.489-490, 2003.
- McEVOY, T.G.; ROBINSON, J.J.; AITKEN, R.P. et al. Dietary excesses of urea influence the viability and metabolism of preimplantation sheep embryos and may affect fetal growth among survivors. Anim. Reprod. Sci., v.47, p.71-90, 1997.
- NUTRIENT requirements of goats: Angora, dairy, and meat goats in temperate and tropical countries. Washington, D.C.: National Academy, 1981. 91p
- OCON, O.M.; HANSEN, P.J. Disruption of bovine oocytes and preimplantation embryos by urea and acidic pH. J. Dairy Sci., v.86, p.1194-1200, 2003.
- OLIVEIRA, M.M.N.F. Uréia para vacas leiteiras no pós-parto: dinâmica folicular e características reprodutivas 2001. 93f. Tese (Doutorado) – Universidade Federal de Viçosa, Viçosa, MG.
- PAPADOPOULOS, S.; LONERGAN, P.; GATH, V. et al. Effect of diet quantity and urea supplementation on oocyte and embryo quality in sheep. Theriogenology, v.55, p.1059-1069, 2001.
- PAYNE, E.; MORRIS, J.G. The effect of protein content of the diet on the rate of urea formation in sheep liver. Biochem. J., v. 113, p. 659, 1969.
- SALLES, H.O.; ANDRIOLI, A.; SIMPLÍCIO, A.A. et al. Manual de transferência de embriões em caprinos Sobral: Embrapa Caprinos, 2002. 64p. (Embrapa Caprinos, Documentos, 40).
- SALLES, H.O. Circuito fechado para a colheita de embriões em caprinos. Disponível em: http://www.cnpc.embrapa.br/topicos/capaartigos/artigo11.htm Acessado em: 12 nov. 2003.
- SILVA, D.J.; QUEIROZ, A.C. Análise de alimentos: métodos químicos e biológicos 3.ed. Viçosa, MG: UFV, 2002. 235p
- SINCLAIR, K.D.; KURAN, M.; GEBBIE, F.E. et al. Nitrogen metabolism and fertility in cattle: II. Development of oocytes recovered from heifers offered diets differing in their rate of nitrogen release in the rumen. J. Anim. Sci., v.78, p.2670-2680, 2000.
- STRINGFELLOW, D.A.; SEIDEL, S.M. Manual da Sociedade Internacional de Transferência de Embriões 3.ed. Illinois: International Embryo Transfer Society, 1999. 180p.
- SISTEMA de análises estatísticas e genéticas. Versão 8.0. Viçosa: UFV, 1997. 150p.
- WHITTINGHAM, D. G. Survival of mouse embryos after freezing and thawing. Nature, v.233, p.125, 1971.
Datas de Publicação
-
Publicação nesta coleção
25 Set 2007 -
Data do Fascículo
Ago 2007
Histórico
-
Aceito
10 Abr 2007 -
Recebido
03 Nov 2005