COMUNICAÇÃO COMMUNICATION
Immunocytochemical characterization of the cytopathic effect induced by bovine respiratory syncytial virus strain RC 98 on Hep-2 cells
Caracterização imunocitoquímica do efeito citopático induzido pelo vírus respiratório sincicial bovino amostra RC98 em células Hep-2
J.M. RavioloI; G. BagnisI; J. AguilarI; J.A. GiraudoI; G.C. ZielinskiII; J.E. RavioloI; C.W. ArnsIII; F.R. SpilkiIV
IUniversidad Nacional de Río Cuarto Enlace Ruta 8 y 36 Km 601 Río Cuarto - Córdoba, Argentina
IISanidad Animal Estación Experimental Marcos Juárez - Córdoba, Argentina
IIIDepartamento de Microbiologia e Imunologia - IB-UNICAMP - Campinas, SP
IVInstituto de Ciências da Saúde -Feevale - Novo Hamburgo, RS
Keywords: BRSV, cytopathic effect, inclusion bodies, syncytial cells, immunocytochemistry
RESUMO
Caracterizou-se o efeito citopático produzido pela amostra de vírus respiratório sincicial bovino (BRSV) RC-98, isolada na Argentina, por meio de imunocitoquímica em cultivos de células da linhagem Hep-2. Um soro policlonal anti-BRSV foi utilizado para a imunocitoquímica em células Hep-2 infectadas. Sinais específicos do virus foram observados no citoplasma de um grande número de células, consistindo em inclusões citoplasmáticas e células sinciciais. Efeitos citopáticos distintos foram observados, com frequência, no núcleo das células infectadas, aparecendo como sinais específicos fortes, podendo corresponder a inclusões intranucleares. A presença de sinais intranucleares pode consistir uma característica particular da amostra RC98 do BRSV.
Palavras-chave: BRSV, efeito citopático, corpúsculos de inclusão, células sinciciais, imunocitoquímica
Bovine respiratory syncytial virus (BRSV) is an enveloped, single-stranded non-segmented, negative-sense RNA virus which belongs to the Pneumovirus genus of the Paramyxoviridae family (Meyer et al., 2008). Its genome has approximately 15,140 nucleotides and encodes 10 different mRNA molecules (Pastey and Samal, 1995; Buchholz et al., 1999; Arns et al., 2003). The genome of BRSV encodes 11 proteins, three of them are transmembrane surface proteins: the G, F, and SH and three other proteins are nonglycosylated virion matrix proteins: M, M2-1, and M2-2. In addition, three proteins associate with genomic RNA to form the viral nucleocapsid: N, P, and L, the last two proteins, NS1 and NS2, are nonstructural (Collins and Crowe, 2007; Valarcher and Taylor, 2007). All steps in the viral replication cycle occur in the cytoplasm, probably following the general schedule for negative strand viruses (Larsen, 2000). The virus was named due to the characteristic cytopathic effect (CPE) that it produces in tissue culture, causing the formation of syncytial cells. The fusion protein (F) is responsible for the fusion of the viral and the host cell membranes and for the generation of syncytia between infected cells (Matheise et al., 1995). Late in the process, the F protein accumulates at the plasma membrane and mediates fusion with neighboring cells to form syncytia (Collins and Crowe, 2007).
Another characteristic cytopathic effect for this virus, is the production of intracytoplasmic inclusion bodies. The nucleocapsid protein N is responsible for the inclusion bodies which are seen in the cytoplasm of BRSV infected cells. Apparently, inclusion bodies lack membranes and contain amorphous material and are exclusively present in the cytoplasm in the vicinity of the cell nuclei (Larsen, 2000).
However, the nucleoproteins (N protein) of porcine reproductive and respiratory syndrome virus (PRRSV) and Coronavirus have been found to have nuclear atypical location (Rowland et al., 1999; Chen et al., 2002). Almeida et al. (2006), using a monoclonal antibody against Human RSV (HRSV) N protein, detected antigens in the nuclei of BRSV infected chicken embryo related cells (CER) in cell cultures. Viral interactions with the nucleolus and its proteins have been found for Retroviruses, RNA and DNA virus (Hiscox, 2002). Many positive and negative strand RNA viruses, whose primary site of replication is the cytoplasm, use the nucleus and/or nuclear components in order to facilitate their replicative processes (Hiscox, 2003).
Although numerous researchers report the cytopathic characteristics of the BRSV, until the present, the cytopathic characteristics produced in vitro by the Argentine BRSV strain RC 98 are not known. The aim of this work was to characterize the cytopathic effect of BRSV strain RC 98, isolated in Argentina, by immunocytochemistry in cultures of Hep-2 cells.
The Argentinean strain BRSV RC98 in passage 33 was used in this work (Bagnis et al., 1999). This strain constitutes the first and only isolation of BRSV from Argentinean cattle up to date. This strain was isolated from a broncoalveolar washing of a dairy calf with respiratory signs in a farm located in Southern Cordoba province (Argentina). Cell line Hep-2, (continuous line of human laryngeal carcinoma) in passage 198 was used in this study. This cell line is known for allowing the infection of the respiratory syncytial virus (Collins and Crowe, 2007). This cell line was also used to detect the first isolation of BRSV in Argentina (Bagnis et al., 1999). Cells were maintained in Eagle's minimum essential medium with Earle's balanced salt solutions (EMEM), supplemented with 200mM L-glutamine, 2% (v/v) fetal calf serum, antibiotics and antimicotic (100IU/mL penicillin, 100µg/mL streptomycin, and 0,25µg/mL anfotericine B1 1 Gibco - USA. ).
Cells were removed from the culture flask and settled in a multi-well plate2 2 Cellstart ® Greiner Bio-One - USA. . A sterilized round coverslip was previously placed in each well. A sterile poly-L-lisine drop was added to the coverslip, which had been dried in an incubator at 37ºC for 24 hours. Hep-2 cells grew over the coverslips during 24 hours. Confluent monolayers were infected with the strain BRSV RC 98 (104,5 DICC50/mL, multiplicity of infection 0.01) and incubated at 37ºC with EMEM, supplemented with 200mM L-glutamine, 5% (v/v) fetal calf serum for 72 hours, until the observation of cytopathic effect. Cells of the same passage were cultured under the same conditions without infecting as negative controls. The culture medium was removed when the CPE extensively affected the cell cultures. Cells were washed with phosphate buffered saline (PBS) and fixed with 100% methanol for 7min at room temperature, passed twice quickly through ethanol (100% and 70%, respectively) and then washed again in PBS (Martin-Cordoba et al., 1996). Endogenous peroxidase was inhibited with 3% hydrogen peroxide in distilled water for 10min at 37ºC. Cells were then washed with PBS and incubated with normal-goat serum (1:20 in PBS) for 20min at room temperature. A polyclonal serum anti BRSV of goat origin with proved reactivity3 3 VMRD Inc. - USA. was used as primary antibody diluted 1:100 and incubated for one hour at 37ºC in a wet chamber.
Cells were rinsed in PBS, and a secondarybiotinylated anti-goat was added and then incubated for 30min, at 37ºC in a wet chamber. After another rinse, the signal was amplified by incubation with an avidin-biotinylated peroxidase complex4 4 Vectastain Elite ABC Kit - Vector Laboratories, Inc. - USA. for 30min at room temperature in a wet chamber. Next, cells were washed and the reaction was developed by adding diaminobenzidine (DAB) substrate Kit for peroxidase5 5 Vector Laboratories, Inc. -USA. to detect the immunoreactive sites.
Nuclear counterstaining was performed with Harris hematoxylin for 15-30 sec. The cells were washed six times with distilled water, then passed in increasing-alcohol concentrations (70%, 80%, 96%, and 100%), 15 to 30sec each for rapid dehydration; finally a rapid pass in xylene (15sec) was done. The coverslips were mounted with Entellan®6 6 Merck - Darmstadt, Germany. for examination under light microscopy.
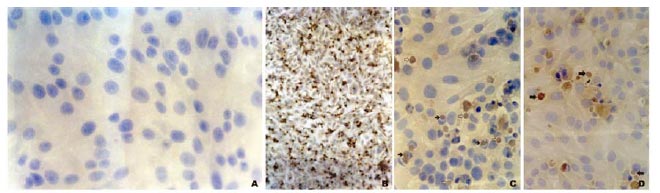
A very strong signal in BRSV infected Hep-2 cells was detected, with no background reaction. The negative control of non-infected cells showed no signal at all (Fig.1-A). The first signs of cytopathic effect were visible approximately at 30 hours post infection, and consisted in rounding of some dispersed cells. This change was explained by Fenner et al. (1992), in which viral proteins were inserted in the cytoplasmic membrane, in initial phases of the replicative cycle, causing damages to the permeability of the membrane that gave rise to rounding by osmotic disbalance. By 72 hours, the CPE was very obvious and involved the majority of the monolayer. This incubation period of 72 hours seems to be very short when compared with other studies that describe a considerable delay (Inaba et al., 1970; Paccaud and Jacquier, 1970; Wellemans et al., 1970; Smith et al., 1975; Kovarcik, 1999). In this study, the ability to produce CPE so rapidly (72 hours post-infection) could have been acquired by this strain due to the high number of passages (33) in the Hep-2 cells.
The typical cytopathic characteristics of BRSV described by several authors were observed (Mohanti and Dutta, 1988; Jubb et al., 1990; Fenner et al., 1992; Larsen, 2000; Spilki et al., 2006; Collins and Crowe, 2007).
In all infected cultures, viral specific signals were observed in the cytoplasm and nucleus of a high number of cells (Fig. 1-B and C). The cytoplasmic signals consisted in large perinuclear intracytoplasmic protein clusters of multiple irregular forms surrounded by a clear haul, which were considered inclusion bodies (Fig. 1-D and E). This perinuclear reaction could be triggered by viral glycoproteins present in the rough endoplasmic reticulum where the synthesis occurs (Almeida et al., 2006). Another characteristic finding was the presence of syncytial cells (Fig. 1-F and G), which were very evident and showed specific signal in the cytoplasm of polykaryocytes. An interesting finding was the frequent detection of strong signal in the nuclei of infected cells. Specific signals were found in the nuclei of a high number of cells which could correspond with intranuclear inclusion bodies (Fig. 1-H and I). This CPE appears to be a particular characteristic induced by the RC 98 strain of BRSV. This finding is very intriguing and appears to disagree with investigations on the replicative cycle of BRSV showing that the events leading into viral replication occur in the cytoplasm, without involving the nucleus (Larsen, 2000).
Intranuclear inclusions have also been described for other viruses of the same family such as canine distemper virus and rinderpest virus, both morbilivirus that produce intracytoplasmic and intranuclear inclusion bodies within the same cell although its replication is totally cytoplasmic (Mohanti and Dutta, 1988; Fenner et al., 1992). Also, Jones et al. (1997) reported the presence of intranuclear inclusions for HRSV. Moreover, a recent study, on cellular culture infected with BRSV, described some infected cells presenting the nuclei stained intensely Almeida et al., 2006. Studies with monoclonal and polyclonal antibodies have detected the N viral protein in the cytoplasm and nucleus of cells infected with other RNA viruses, such as PRRSV and Coronaviruses (Rowland et al., 1999; Chen et al., 2002)
According to Hiscox et al. (2001) and Wurm et al. (2001), the N protein could be actively transported to the nucleolus. Molecules can enter the nucleus by passive diffusion or active transport mechanisms, depending on their size (Macara, 2001). Small molecules up to 50, 60kDa or less than 10nm diameter can passively diffuse through the nuclear pore complex, but most proteins are transported by energy driven transport mechanisms (Richardson et al., 1988). Active transport of proteins is mediated by nuclear localization signals. These signals are recognized by proteins of the importin super-family that mediate the transport across the nuclear envelope (Macara, 2001). It could be speculated that these events could also occur with the N protein of BRSV.
Although cytoplasmic RNA viruses confine their principal replicative functions to membrane bound structures in the cytoplasm (Kujala et al., 2001; Gosert et al., 2002; Lyle et al., 2002); many positive and negative strand RNA viruses, whose primary site of replication is the cytoplasm, sequester nuclear factors in order to facilitate their replication and, by altering the nuclear-cytoplasmic trafficking, can disrupt host cell functions and cellular responses to viral infections (Hiscox, 2003). Gustin and Sarnow (2002) suggested that the redistribution of nuclear proteins and disruption of the nuclear pore complex might be part of a strategy by which cytoplasmic RNA viruses could avoid triggering the host immune response by blocking nuclear signaling pathways.
The identification of nuclear signal in this study might be attributed to the multiplicity of infection, to the type of cellular culture, and/or to the sensitivity and specificity of the technique. In this sense, the immunocytochemistry technique applied here by means of a highly specific policlonal antibody, plus the nuclear counterstain with haematoxylin, allowed to identify with clarity the cellular compartments where the signals were observed. Other authors may have not seen the nuclear changes in the past because of the use of less accurate techniques: cell cultures infected by BRSV without staining (Fenner et al., 1992), stained with haematoxylin and eosin (Fenner et al., 1992; Kovarcik, 1999), or histological cuts (Jubb et al., 1990).
In conclusion, a strong permissiveness of the cellular line Hep-2 was observed for the infection of BRSV strain RC 98. Immunocytochemistry allowed a clear detection and characterization of the cytopathic effect of BRSV on Hep-2 cells consisting of the typical syncytial cells and intracytoplasmic inclusion bodies, plus the intranuclear inclusions described here. The functional relevance of these viral proteins localized in the nucleus and how this relates to virus replication are still unknown. More studies are necessary to analyze the significance of this result.
Recebido em 23 de setembro de 2008
Aceito em 16 de março de 2009
E-mail: jraviolo@ayv.unrc.edu.ar
- ALMEIDA, R.S.; SPILKI, F.R.; ROEHE, P.M. et al. Bovine respiratory syncytial virus: immunohistochemichal detection in mouse and bovine tissues using a Mab against human respiratory syncytial virus. Arq. Bras. Med. Vet. Zootec, v.58, p.973-981, 2006.
- ARNS, C.W.; CAMPALANS, J.; COSTA, S.C.B. et al. Characterization of bovine respiratory syncytial virus isolated in Brazil. Braz. J. Med. Biol. Res, v.36, p.213-218, 2003.
- BAGNIS, G.; GIRAUDO, J.A.; SUTIL, S. et al. Aislamiento y detección antigénica del virus sincitial respiratorio bovino en la Argentina. Rev. Med. Vet, v.80, p.173-177, 1999.
- BUCHHOLZ, U.J.; FINKE, S.; CONZELMANN, K.K. Generation of bovine respiratory syncytial virus (BRSV) from cDNA: BRSV NS2 is not essential for virus replication in tissue culture, and the human RSV leader region acts as a functional BRSV genome prometer. J. Virol., v.73, p.251-259, 1999.
- CHEN, H.; WURM, T.; BRITTON, P. et al. Interation of the coronavirus nucleoprotein with nucleolar antigens and the host cell. J. Virol, v.76, p.5233-5250, 2002.
- COLLINS, P.L.; CROWE, J.E. Respiratory syncytial virus and metapneumovirus. In. KNIPE, D.M.; HOWLEY. P.M. (Eds). Fields Virology. Philadelphia:Lippincott-Williams and Wilkins, 2007. p.1601-1646.
- FENNER, F.; BACHMANN, P.A.; GIBBS, E.P.J. et al. Virologia veterinaria. Zaragoza, España: Acríbia, 1992.
- GOSERT, R.; KANJANAHALUETHAI, A.; EGGER, D. et al. RNA replication of mouse hepatitis virus takes place at double-membrane vesicles. J. Virol, v.76, p.3697-3708, 2002.
- GUSTIN, K.E., SARNOW, P. Inhibition of nuclear import and alteration of nuclear pore complex composition by rhinovirus. J. Virol, v.76, p.8787-8796, 2002.
- HISCOX, J.A. Brief review: the nucleolus a gateway to viral infection. Arch. Virol., v.147, p.1077-1089, 2002.
- HISCOX, J.A. The interaction of animal cytoplasmic RNA viruses with the nucleus to facilitate replication. Virus Res., v.95, p.13-22,2003.
- HISCOX, J.A.; WURM, T.; WILSON, L. et al. The coronavirus infectious bronchitis virus nucleoprotein localizes to the nucleolus. J. Virol., v.75, p.506-512, 2001.
- INABA, Y.; TANAKA, Y.; OMORI, T. et al. Isolation of bovine respiratory syncytial virus. Jpn. J. Exp. Med., v.40, p.473-474, 1970.
- JONES, T.C.; HUNT, R.D.; KING, N.W. Veterinary Pathology 6.ed. Lippincot Williams & Wilkins, 1997.
- JUBB, K.V.F.; KENNEDY, P.C.; PALMER, N. Patología de los animales domésticos 3.ed. Montevideo: Agropecuaria Hemiferio Sur Montevideo, 1990. v.2
- KOVARCIK, K. Isolation of bovine respiratory syncytial virus during and outbreak of acute respiratory disease in calves. Vet. Med. Czech, v.44, p.121-127, 1999.
- KUJALA, P.; IKAHEIMONEN, A.; EHSANI, N. et al. Biogenesis of the Semliki Forest virus RNA replication complex. J. Virol, v.75, p.3873-3884, 2001.
- LARSEN, L.E. Bovine respiratory syncytial virus (BRSV): a review. Acta Vet. Scand, v.41, p.1-21, 2000.
- LYLE, J.M.; BULLITT, E.; BIENZ, K. et al. Visualization and functional analysis of RNA-dependent RNA polymerase lattices. Science, v.296, p.2218-2222, 2002.
- MACARA, I.G. Transport into and out of the nucleus. Microbiol. Mol. Biol. Rev., v.65, p.570-594, 2001.
- MARTIN-CORDOBA, C.G.; REGADERA, J.; HERNANDEZ, A. et al. Immunocytochemical methods in cell cultures for evaluations of cell cycle and differentiations in normal and neoplastic tissues. J. Histotechnol., v.19, p.109-114, 1996.
- MATHEISE, J.P.; WALRAVENS, K.; COLLARD, A. et al. Antigenic analysis of the F protein of the bovine respiratory syncytial virus: identification of two distinct antigenic sites involved in fusion inhibition. Arch Virol., v.140, p.993-1005, 1995.
- MEYER, G.; DEPLANCHE, M.; SCHELCHER, F. Human and bovine respiratory syncytial virus vaccine research and development. Comp. Immun. Microbiol. Infect. Dis., v.31, p.191-225, 2008.
- MOHANTI, S.B.; DUTTA, S.K. Virología veterinaria 1.ed. México: Interamericana, 1998.
- PACCAUD, M.F.; JACQUIER, C. A respiratory virus of bovine origin. Arch. Ges. Virusforsh., v.303, p.27-42, 1970.
- PASTEY, M.K.; SAMAL, S.K. Nucleotide sequence analysis of the non-structural NS1 (1C) and NS2 (1B) protein genes of bovine respiratory syncytial virus. J. Gen. Virol., v.76, p.193-197, 1995.
- RICHARDSON, W.D.; MILLS, A.D.; DILWORTH, S.M. et al. Nuclear protein migration involves two steps: rapid binding at the nuclear envelope followed by slower translocation through nuclear pores. Cell, v.52, p.655-664, 1988.
- ROWLAND, R.R.; KERVIN, R.; KUCKLEBURG, C. et al. The localization of porcine reproductive and respiratory syndrome virus nucleocapsid protein to the nucleolus of infected cells and identification of a potential nucleolar localization signal sequence. Virus Res., v.64, p.1-12, 1999.
- SMITH, M.H.; FREY, M.L.; DIERKS, R.E. Isolation, characterization and pathogenicity studies of a bovine respiratory syncytial virus. Arch. Virol., v.47, p.237-247, 1975.
- SPILKI, F.R.; ALMEIDA, R.S.; CAMPALANS, J. et al. Susceptibility of different cell lines to infection with bovine respiratory syncytial virus. J. Virol. Methods, v.131, p.130-131, 2006.
- VALARCHER, J.F.; TAYLOR, G. Bovine respiratory syncytial virus infection. Vet. Res., v.38, p.153-180, 2007.
- WELLEMANS, G.; LEUNEN, J.; LUCHSINGER, E. Isolement d'un virus (220/69) serologiquement semblable au virus respiratoire syncytial (RS) humain (Isolation of a virus (220/69) serologically resembling human respiratory syncytial virus). Ann. Med. Vet., v.114, p.89-93, 1970.
- WURM, T.; CHEN, H.; BRITTON, P. et al. Localization to the nucleolus is a common feature of coronavirus nucleoproteins and the protein may disrupt host cell division. J. Virol., v.75, p.9345-9356, 2001.
Publication Dates
-
Publication in this collection
27 Aug 2009 -
Date of issue
Aug 2009