Resumos
Este estudo buscou clonar o cDNA do sbGnRH, identificar sua expressão em diferentes tecidos do linguado, bem como avaliar possíveis diferenças no RNA mensageiro (RNAm) desse gene no cérebro de linguados machos juvenis e adultos. Por meio da RT-PCR, demonstrou-se pela primeira vez, a clonagem da região codificadora do sbGnRH contendo 297 nucleotídeos do cérebro do linguado. A expressão do sbGnRH foi detectada em vários tecidos periféricos. Foram detectados níveis mais elevados de RNAm do sbGnRH no hipotálamo dos animais adultos. Estes resultados sugerem que o sbGnRH está envolvido na puberdade do linguado.
peixe; sbGnRH; RNAm; puberdade
The objectives of this study were to clone sbGnRH cDNA, evaluate the mRNA levels in different tissues of flounder, and also evaluate brain sbGnRH expression in juvenile and adult males. Using RT-PCR the cloning of a 297 nucleotides coding region of sbGnRH from Brazilian flounder brain was demonstrated for the first time. Expression of sbGnRH was detected in several peripheral tissues. Brain gene expression in the adult flounder was higher than those found in juvenile. These results suggest that sbGnRH is involved on the Brazilian flounder puberty.
fish; sbGnRH; mRNA; puberty
ZOOTECNIA E TECNOLOGIA E INSPEÇÃO DE PRODUTOS DE ORIGEM ANIMAL
Clonagem e avaliação da expressão gênica do sbGnRH em machos juvenis e adultos de linguado, Paralichthys orbignyanus
Cloning and evaluation of sbGnRH gene expression in juvenile and adult males of Brazilian flounder Paralichthys orbignyanus
V.F. CamposI; T. CollaresI; J.C. DeschampsI; F.K. SeixasI; M.H. OkamotoII; L.A. SampaioII; L.F. MarinsII; R.B. RobaldoI,** Autor para correspondência ;
IUniversidade Federal de Pelotas - UFPel Caixa Postal 354 96010-900 - Pelotas, RS
IIUniversidade Federal do Rio Grande - Rio Grande, RS
RESUMO
Este estudo buscou clonar o cDNA do sbGnRH, identificar sua expressão em diferentes tecidos do linguado, bem como avaliar possíveis diferenças no RNA mensageiro (RNAm) desse gene no cérebro de linguados machos juvenis e adultos. Por meio da RT-PCR, demonstrou-se pela primeira vez, a clonagem da região codificadora do sbGnRH contendo 297 nucleotídeos do cérebro do linguado. A expressão do sbGnRH foi detectada em vários tecidos periféricos. Foram detectados níveis mais elevados de RNAm do sbGnRH no hipotálamo dos animais adultos. Estes resultados sugerem que o sbGnRH está envolvido na puberdade do linguado.
Palavras-chave: peixe, sbGnRH, RNAm, puberdade
ABSTRACT
The objectives of this study were to clone sbGnRH cDNA, evaluate the mRNA levels in different tissues of flounder, and also evaluate brain sbGnRH expression in juvenile and adult males. Using RT-PCR the cloning of a 297 nucleotides coding region of sbGnRH from Brazilian flounder brain was demonstrated for the first time. Expression of sbGnRH was detected in several peripheral tissues. Brain gene expression in the adult flounder was higher than those found in juvenile. These results suggest that sbGnRH is involved on the Brazilian flounder puberty.
Keywords:fish, sbGnRH, mRNA, puberty
INTRODUÇÃO
O hormônio liberador de gonadotrofinas (GnRH) desempenha papel-chave no desempenho e na manutenção das funções reprodutivas nos vertebrados (Guilgur et al., 2006). Três principais isoformas do GnRH foram identificadas em pleuronectiformes: sGnRH (salmon GnRH), cGnRH-II (chicken GnRH-II) e sbGnRH (sea bream GnRH) (Amano et al., 2002; Pham et al., 2007), e a isoforma sbGnRH foi identificada como a provável responsável pela maturação gonadal (puberdade) nesses peixes (Amano et al., 2008). Além disso, a expressão desse gene parece não estar restrita ao sistema nervoso central dos peixes. Alguns relatos têm demonstrado a expressão do sbGnRH fora do cérebro, em tecidos e órgãos como testículos, ovários, olhos, baço, brânquias e coração (Guilgur et al., 2007). Entretanto, ainda são desconhecidas as funções fisiológicas deste hormônio nestes tecidos.
Diversos estudos têm demonstrado que o conhecimento dos aspectos moleculares de regulação da puberdade pode contribuir significativamente para o incremento da criação de peixes em cativeiro (Taranger et al., 2010). Contudo, para um controle seguro da puberdade na piscicultura, é necessário compreender os mecanismos subjacentes ao desencadeamento da primeira maturação gonadal, bem como o impacto de vários fatores internos e externos que regulam e modulam esse processo.
O linguado, Paralichthys orbignyanus, habita águas de estuários e costeiras desde o Rio de Janeiro, no Brasil, até Mar del Plata, na Argentina (Figueiredo e Menezes, 2000), onde reproduz durante os meses da primavera, atingindo a primeira maturação gonadal com tamanho mínimo de 26cm para os machos e 38cm para as fêmeas (Silveira et al., 1995). Estudos recentes relacionados à larvicultura (Sampaio et al., 2007), reprodução (Lanes et al., 2008; Sampaio et al., 2008), potencial para transgênese (Lanes et al., 2009) e expressão de genes ligados ao crescimento e ao controle da fome (Meier et al., 2009; Campos et al., 2010) demonstram a viabilidade do cultivo desta espécie. No entanto, ainda não há estudos sobre os mecanismos moleculares e fisiológicos sobre o controle da puberdade nesta espécie. Este estudo teve o objetivo de isolar e caracterizar o cDNA do sbGnRH no cérebro do linguado, identificar em quais tecidos este gene é expresso e comparar a expressão gênica no cérebro do sbGnRH em linguados pré e pós-púberes.
MATERIAL E MÉTODOS
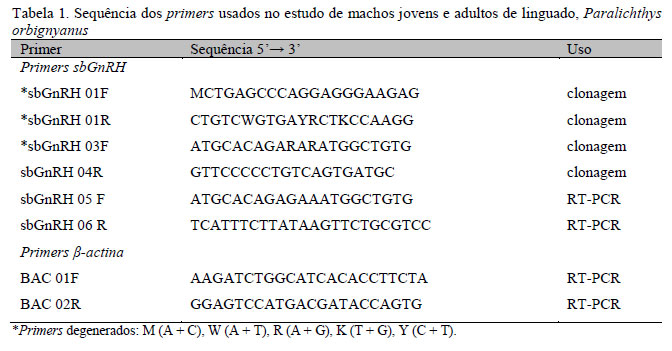
Os peixes usados foram obtidos por meio de desova artificial. Dois linguados machos adultos, de 16 meses de idade, com pesos corporais de 285 e 283 gramas e 28 e 25cm de comprimento padrão, foram usados para a clonagem do sbGnRH. Os peixes foram anestesiados com benzocaína (50mg.L-1) e sacrificados pela ruptura da medula espinhal. Imediatamente após a eutanásia, o cérebro foi dissecado e o hipotálamo foi isolado e armazenado em nitrogênio líquido até o uso (González-Martínez et al., 2002; Amano et al., 2004). O RNA total foi isolado usando-se o reagente TRIzol® (InvitrogenTM, EUA), e o tratamento com DNase foi realizado com o kit DNA-free® (AmbionTM, EUA) seguindo as instruções do fabricante. Dois microgramas de RNA foram usados para a síntese do cDNA, realizada com o kit High Capacity cDNA Reverse Transcription® (Applied BiosystemsTM, EUA), seguindo as instruções do fabricante. Para a clonagem do sbGnRH, foram construídos primers (Tab. 1) baseados no alinhamento de sequências do sbGnRH ou GnRH tipo I de outras espécies de peixes. As PCR seguiram os seguintes parâmetros: desnaturação inicial de 94ºC/5min, seguida de 35 ciclos de 94ºC/30s, 50ºC/30s, 72ºC/1min, seguida por uma extensão final de 72ºC/5min. Os produtos das reações foram sequenciados no sequenciador automático MegaBACE 1000 (Amersham Biosciences, EUA).
Várias sequências codificadoras do sbGnRH, incluindo a sequência do P. orbignyanus, foram alinhadas usando-se o CLUSTAL X (Thompson et al., 1997), e a análise filogenética foi realizada usando-se o software Phylogeny Inference Package PHYLYP 3.6 (Felsenstein, 1993) e o método de máxima parcimônia com a espécie Xenopus laevis como outgroup. Uma análise de bootstrapping com 1000 interações foi realizada utilizando-se SEQBOOT (Felsenstein, 1993).
Para avaliação da distribuição dos tecidos do sbGnRH, amostras de brânquias, coração, intestinos, fígado, baço, rins, músculos, estômago, testículos e cérebro foram coletadas e processadas, e a extração de RNA total, o tratamento com DNase e a síntese de cDNA foram realizadas como descrito anteriormente. As PCRs seguiram os parâmetros descritos antes usando-se os primers sbGnRH 05 e 06 (Tab. 1). Os produtos de PCR foram submetidos à eletroforese em géis de agarose 1% contendo 0,5µgml-1 de brometo de etídio.
A avaliação dos efeitos do estádio de maturação na expressão gênica do sbGnRH no cérebro foi realizada em cinco machos juvenis com 10 meses de idade e peso e comprimento padrão de 71,4±13,0g e 17,8±1,0cm, respectivamente, e em cinco machos adultos com 20 meses de idade e peso e comprimento padrão de 463,2±42,2g e 34,1±0,6cm, respectivamente. Os peixes foram mantidos em um mesmo tanque com volume útil de água de 2,0m3, sob densidade de estocagem de 3,5kg.m-3, durante três semanas para aclimatação, a 21.5º±0.8ºC de temperatura, 32.6±1.1‰ de salinidade, 6.8±0.3mg O2.L-1 de oxigênio dissolvido e fotoperíodo de 12L:12E. Todos os peixes foram alimentados uma vez ao dia com ração comercial (Supra SalmonídeosTM - Alisul, Brasil) contendo 46% de proteína bruta e 6% de lipídios. Ao término da aclimatação, os exemplares foram aleatoriamente amostrados. Os animais foram anestesiados, eutanasiados, e a coleta das amostras de hipotálamo foram realizadas como descrito anteriormente. A extração de RNA total, o tratamento com DNase e a síntese de cDNA foram realizadas como descrito anteriormente. A análise da expressão gênica foi realizada por meio de RT-PCR semiquantitativa, sendo utilizada a β-actina com gene normalizador (Mori et al., 2008). As condições da PCR também seguiram os procedimentos já descritos, e usaram-se os primers sbGnRH 05 e 06 e BAC 01 e 02 (Tab. 1). Para ambos os genes analisados, foram realizados vários testes para a determinação do número de ciclos necessários para produção de uma amplificação detectável. Para o sbGnRH, foram usados 27 ciclos, e para o controle interno β-actina, 32 ciclos. Todas as amostras foram submetidas à eletroforese e aplicadas no mesmo gel para posterior comparação. Para densidade óptica das bandas, usou-se o software TotalLab TL 100 (Nonlinar Dynamics, Inglaterra). A taxa de densidade óptica sbGnRH/β-actina foi utilizada para o cálculo de expressão gênica, segundo Meier et al. (2009).
Os dados referentes à expressão gênica no hipotálamo de linguados juvenis e adultos foram comparados por meio do teste t e nível de significância estabelecido em P<0.05.
RESULTADOS E DISCUSSÃO
A região codificadora (CDS) do sbGnRH contendo 297 nucleotídeos foi clonada do cérebro do linguado e depositada no GenBank sob o acesso GQ220297. Como outros neuropeptídeos, o cDNA do sbGnRH do linguado codifica para um peptídeo tripartido formado por 98 resíduos de aminoácidos com um peptídeo sinal (resíduos 1-26), o sbGnRH (resíduos 27-36), e um peptídeo associado (resíduos 40-98). Este último e o sbGnRH são unidos por um sítio proteolítico Gly-Lys-Arg (resíduos 37-39). Vários estudos têm demonstrado a mesma organização geral para o gene do sbGnRH com um peptídeo sinal, um decapeptídeo correspondente ao sbGnRH, um sítio proteolítico e um peptídeo associado (Guilgur et al., 2007; An et al., 2008). A análise filogenética (Fig. 1) demonstrou estreita relação do sbGnRH entre os membros da ordem dos Pleuronectiformes, incluindo o P. orbignyanus. Por estes resultados, verifica-se que a sequência de cDNA clonada é realmente a sequência codificadora do sbGnRH (GQ220297) do linguado.
Vários estudos têm documentado a expressão gênica do GnRH no sistema nervoso central, entretanto apenas recentemente foi documentada a presença do GnRH em tecidos periféricos. No presente estudo, a expressão do sbGnRH foi detectada nos tecidos periféricos dos seguintes órgãos: estômago, fígado, intestinos, rins, baço, coração, testículos, brânquias e cérebro (Fig. 2). A exceção foram os tecidos musculares. Experimentos em diferentes espécies de peixes têm demonstrado evidências da expressão periférica do mRNA do sbGnRH (Guilgur et al., 2009), no entanto Hu et al. (2008) demonstraram a ausência da expressão do sbGnRH nos tecidos periféricos. O papel deste peptídeo nesses tecidos, contudo, ainda é desconhecido e sugere que este hormônio esteja envolvido em várias funções não relacionadas à reprodução.
Foram detectados níveis mais elevados de RNAm do sbGnRH no hipotálamo dos animais adultos (Fig. 3). Estudos realizados durante a estação reprodutiva em fêmeas de Paralichthys olivaceus demonstraram níveis significativamente mais elevados de sbGnRH no hipotálamo, telencéfalo e pituitária que das variantes cGnRH-II e sGnRH, e redução desses níveis fora da estação reprodutiva (Pham et al., 2006a, b). Também em fêmeas do linguado japonês foram detectados neurônios hipotalâmicos inervando a pituitária e contendo altos níveis do sbGnRH (Pham et al., 2007). Esses estudos demonstraram que o sbGnRH está provavelmente envolvido na maturação gonadal no P. olivaceus. Em outro pleuronectiforme, Verasper moseri, os níveis de RNAm do sbGnRH no cérebro e a quantidade de sbGnRH na pituitária elevaram-se à medida que o índice gonadossomático aumentou (Amano et al., 2004). Da mesma forma, os resultados apresentados neste trabalho indicaram que o sbGnRH está envolvido na maturação testicular no P. orbignyanus.
Em outras espécies como a lubina Dicentrarchus labrax entre as três formas de GnRH, os níveis de sbGnRH aumentaram na pituitária durante a estação reprodutiva (Rodriguez et al., 2000). No badejo, Sebastes rastrelliger, apenas o sbGnRH foi detectado em níveis significativos no cérebro e na pituitária (Collins et al., 2001). Todos esses resultados em diferentes espécies, nas quais os níveis do sbGnRH no cérebro e na pituitária aumentaram concomitantemente com a maturação gonadal, suportam os resultados apresentados neste trabalho.
Embora os mecanismos precisos do início da puberdade nos peixes ainda não tenham sido elucidados, sabe-se que sinais derivados de fatores internos e externos estimulam a liberação hipotalâmica de GnRH, o qual, por sua vez, estimula a produção e liberação das gonadotrofinas (GTH) na pituitária (Dufour e Rousseau, 2007). Estudos recentes com o pargo negro, Acanthopagrus schlegeli, demonstraram que o GnRH é responsável pela indução da secreção das subunidades de GTH (GTH-α, FSH-β e LH-β) (An et al., 2009). De acordo com Kumakura et al. (2004), da mesma forma que aqui demonstrado para P. orbignyanus, exemplares pré-púberes do pargo, Pagrus major, apresentam reduzidos níveis de RNAm do sbGnRH, se comparados aos de adultos. Entretanto, em contraste com os resultados evidenciados para o pargo negro, juvenis de P. major, quando tratados com análogos do GnRH, apresentaram aumento dos níveis de expressão da gonadotrofinas FSH-β e LH-β, apesar de não terem alterado a expressão gênica do sbGnRH (Kumakura et al., 2004).
Recentemente, um novo fator foi reconhecido no campo da fisiologia reprodutiva de peixes: o peptídeo Kiss1 e seu receptor GPR54, os quais estão envolvidos no estímulo da produção e secreção do GnRH (Martinez-Chavez et al., 2008; Zohar et al., 2010).
Para uma melhor compreensão da maneira como o sbGnRH age na maturação gonadal e no controle da puberdade, são necessários estudos que envolvam as relações entre Kiss1/GPR54, sbGnRH e, consequentemente, a liberação das gonadotrofinas no P. orbignyanus.
CONCLUSÕES
A clonagem do cDNA do sbGnRH do linguado P. orbignyanus foi obtida com êxito. Foi detectada expressão do sbGnRH no estômago, fígado, intestinos, rins, baço, coração, testículos, brânquias e cérebro. Machos adultos apresentaram maior expressão do sbGnRH, indicando que este está associado à puberdade da espécie, porém deve desempenhar funções ainda desconhecidas em tecidos periféricos, onde se expressa.
AGRADECIMENTOS
Este trabalho contou com o apoio do MCT/CNPq-Edital Universal (# 47438/2006-7).
Recebido em 26 de janeiro de 2010
Aceito em 28 de dezembro de 2010
E-mail: ricardo.robaldo@pq.cnpq.br
- AMANO, M.; OKUBO, K.; YAMANOME, T. et al. Changes in brain GnRH mRNA and pituitary GnRH peptide during testicular maturation in barfin flounder. Comp. Biochem. Physiol. B, v.138, p.435-443, 2004.
- AMANO, M.; PHAM, K.X.; AMIYA, N. et al. Changes in brain seabream GnRH mRNA and pituitary seabream GnRH peptide levels during ovarian maturation in female barfin flounder. Gen. Comp. Endocrinol, v.158, p.168-172, 2008.
- AMANO, M.; TAKAHASHI, A.; YAMANOME, T. et al. Molecular cloning of three cDNAs encoding different GnRHs in the brain of barfin flounder. Gen. Comp. Endocrinol., v.126, p.325-333, 2002.
- AN, K.W.; LEE, K.; YUN, S.G. et al. Molecular characterization of gonadotropin subunits and gonadotropin receptors in black porgy, Acanthopagrus schlegeli: Effects of estradiol-17β on mRNA expression profiles. Comp. Biochem. Physiol. B, v.152, p.177-188, 2009.
- AN, K.W.; NELSON, E.R.; HABIBI, H.R. et al. Molecular characterization and expression of three GnRH forms mRNA during gonad sex-change process, and effect of GnRHa on GTH subunits mRNA in the protandrous black porgy (Acanthopagrus schlegeli). Gen. Comp. Endocrinol, v.159, p.38-45, 2008.
- CAMPOS, V.F.; COLLARES, T.; DESCHAMPS, J.C. et al. Identification, tissue distribution and evaluation of brain neuropeptide Y gene expression in the Brazilian flounder Paralichthys orbignyanus. J. Biosc, v.35, p.405-413, 2010.
- COLLINS, P.M.; O'NEILL, D.F.; BARRON, B.R. et al. Gonadotropin-releasing hormone content in the brain and pituitary of male and female grass rockfish (Sebastes rastrelliger) in relation to seasonal changes in reproductive status. Biol. Reprod., v.65, p.173-179, 2001.
- DUFOUR, S.; ROUSSEAU, K. Neuroendocrinology of fish metamorphosis and puberty: evolutionary and ecophysiological perspectives. J. Mar. Sci. Technol., v.15, p.55-68, 2007.
- FELSENSTEIN, J. Phylogeny Inference Package (PHYLIP). Version 3.5. Seattle: University of Washington, 1993.
- FIGUEIREDO, J.L.; MENEZES, N.A. (Eds.). Manual de peixes marinhos do sudeste do Brasil VI. Teleostei (5). São Paulo: Museu de Zoologia/USP, 2000. 116p.
- GONZÁLEZ-MARTÍNEZ, D.; ZMORA, N.; ZANUY, S. et al. Developmental expression of three different prepro-GnRH (Gonadotrophin-releasing hormone) messengers in the brain of the European sea bass (Dicentrarchus labrax). J. Chem. Neuroanat., v.23, p.255-267, 2002.
- GUILGUR, L.G.; MONCAUT, N.P.; CANÁRIO, A.V.M. et al. Evolution of GnRH ligands and receptors in Gnathostomata. Comp. Biochem. Physiol. A, v.144, p.272-283, 2006.
- GUILGUR, L.G.; ORTÍ, G.; STROBL-MAZZULLA, P.H. et al. Characterization of the cDNAs encoding three GnRH forms in the pejerrey fish Odontesthes bonariensis (Atheriniformes) and the evolution of GnRH precursors. J. Mol. Evol, p.64, p.614-627, 2007.
- GUILGUR, L.G.; STRÜSSMANN, C.A.; SOMOZA, G.M. mRNA expression of GnRH variants and receptors in the brain, pituitary and ovaries of pejerrey (Odontesthes bonariensis) in relation to the reproductive status. Fish Physiol. Biochem, v.35, p.157-166, 2009.
- HU, S.Y.; CHEN, M.H.; LIN, Y. et al. Cloning and functional analysis of the proximal promoter region of the three GnRH genes from the silver sea bream (Sparus sarba). Comp. Biochem. Physiol. B, v.151, p.373-380, 2008.
- KUMAKURA, N.; OKUZAWA, K.; GEN, K. et al. Effects of gonadotropin-releasing hormone on pituitary-ovarian axis of one-year old pre-pubertal red seabream. Gen. Comp. Endocrinol., v.138, p.105-112, 2004.
- LANES, C.F.C.; OKAMOTO, M.; CAVALCANTI, P.V. et al. Cryopreservation of Brazilian flounder (Paralichthys orbignyanus) sperm. Aquaculture, v.275, p.361-365, 2008.
- LANES, C.F.C.; SAMPAIO, L.A.; MARINS, L.F. Evaluation of DNase activity in seminal plasma and uptake of exogenous DNA by spermatozoa of the Brazilian flounder Paralichthys orbignyanus. Theriogenology, v.71, p.525-533, 2009.
- MARTINEZ-CHAVEZ, C.C.; MINGHETTI, M.; MIGAUD, H. GPR54 and rGnRH I gene expression during the onset of puberty in Nile tilapia. Gen. Comp. Endocrinol, v.156, p.224-233, 2008.
- MEIER, K.M.; FIGUEIREDO, M.A.; KAMIMURA, M.T. et al. Increased growth hormone (GH), growth hormone receptor (GHR), and insulin-like growth factor I (IGF-I) gene transcription after hyperosmotic stress in the Brazilian flounder Paralichthys orbignyanus. Fish Physiol. Biochem, v.35, p.501-509, 2009.
- MORI, R.; WANG, Q.; DANENBERG, K.D. et al. Both beta-actin and GAPDH are useful reference genes for normalization of quantitative RT-PCR in human FFPE tissue samples of prostate cancer. Prostate, v.68, p.1555-1560, 2008.
- PHAM, K.X.; AMANO, M.; AMIYA, N. et al. Immunohistochemical localization of three GnRH systems in brain and pituitary of Japanese flounder. Fish Sci, v.73, p.1113-1122, 2007.
- PHAM, K.X.; AMANO, M.; AMIYA, N. et al. Distribution of three GnRHs in the brain and pituitary of the wild Japanese flounder Paralichthys olivaceus. Fish Sci, v.72, p.89-94, 2006a.
- PHAM, K.X.; AMANO, M.; AMIYA, N. et al. Changes in brain and pituitary GnRH levels during ovarian maturation in wild female Japanese flounder. Fish Physiol. Biochem, v.32, p.241-248, 2006b.
- RODRIGUEZ, L.; CARRILLO, M.; SORBERA, L.A. et al. Pituitary levels of three forms of GnRH in the male European sea bass (Dicentrarchus labrax, L.) during sex differentiation and first spawning season. Gen. Comp. Endocrinol, v.120, p.67-74, 2000.
- SAMPAIO, L.A.; FREITAS, L.S.; OKAMOTO, M.H. et al. Effects of salinity on Brazilian flounder Paralichthys orbignyanus from fertilization to juvenile settlement. Aquaculture, v.262, p.340-346, 2007.
- SAMPAIO, L.A.; ROBALDO, R.B.; BIANCHINI, A. Hormone-induced ovulation, natural spawning and larviculture of Brazilian flounder Paralichthys orbignyanus (Valenciennes, 1839). Aquac. Res, v.39, p.712-717, 2008.
- SILVEIRA, M.P.M.; COUSIN, J.C.B.; HAIMOVICI, M. Estrutura ovárica e testicular do linguado Paralichthys orbignyanus (Valenciennes, 1839). Atlântica, v.17, p.135-152, 1995.
- TARANGER, G.L.; CARRILLO, M.; SCHULZ, R.W. et al. Control of puberty in farmed fish. Gen. Comp. Endocrinol, v.165, p.483-515, 2010.
- THOMPSON, J.D.; GIBSON, T.J.; PLEWNIAK, F. et al. The Clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res, v.24, p.4876-4882, 1997.
- ZOHAR, Y.; MUÑOZ-CUETO, J.A.; ELIZUR, A. et al. Neuroendocrinology of reproduction in teleost fish. Gen. Comp. Endocrinol, v.165, p.438-455, 2010.
Datas de Publicação
-
Publicação nesta coleção
11 Abr 2011 -
Data do Fascículo
Fev 2011
Histórico
-
Recebido
26 Jan 2010 -
Aceito
28 Dez 2010