Resumos
O objetivo deste trabalho foi verificar os efeitos da ingestão materna de diferentes doses de cafeína durante a gestação e a lactação, na pele de ratas-mães e filhotes, bem como sua relação com as concentrações séricas do cortisol materno. Vinte e quatro ratas Wistar adultas foram distribuídas em quatro grupos, representados pelo controle e tratados, com cafeína nas doses de 25, 50 e 100mg/kg. Os grupos tratados receberam cafeína por sonda orogástrica durante toda a gestação e a lactação. O controle recebeu água destilada como placebo. Foram avaliados e quantificados os diferentes tipos de folículos pilosos e a espessura da epiderme. A técnica de imuno-histoquímica, com o uso do anticorpo anti-CDC47, foi utilizada para avaliar a proliferação celular da epiderme e dos folículos pilosos das mães. Na mãe, também foram mensurados os níveis séricos de cortisol pela técnica da quimioluminescência. Os dados foram submetidos à análise de variância com comparação das médias pelos testes Kruskal-Wallis e SNK. Nos grupos tratados com cafeína nas doses de 25 e 50mg/kg, tanto as mães quanto seus filhotes apresentaram hipotricose e/ou alopecia focal. Apesar de a frequência de alterações macroscópicas das mães ter sido superior a dos filhotes, nestes as lesões, quando presentes, foram difusas. A análise histológica demonstrou calcinose de folículos pilosos nas mães e nos filhotes. Mas a morfometria somente revelou diferença significativa no número de folículos pilosos das mães, bem como redução significativa da proliferação celular dos folículos pilosos do grupo tratado com 50mg/kg de cafeína. Os níveis de cortisol materno somente foram significativamente elevados no grupo tratado com 100mg/kg de cafeína. Conclui-se que a cafeína ingerida pelas ratas gestantes e lactantes pode causar lesões cutâneas tanto nas mães quanto nos filhotes, caracterizadas por hipotricose e/ou alopecia, independentemente dos níveis séricos do cortisol materno.
rata; cafeína; pele; alopecia; cortisol
The aim of this study was to investigate the effects of maternal caffeine intake during pregnancy and lactation on the skin of rats and their offspring, as well as their relationship to maternal serum levels of cortisol. 24 adult Wistar rats were equally divided into four groups represented by the control and treated with caffeine at doses of 25, 50 and 100mg/kg. The groups received caffeine by orogastric tube during the entire pregnancy and lactation. The control received distilled water as placebo. Different types of hair follicles and the thickness of the epidermis were assessed and quantified. Immunohystochemistry technique using antibody anti-CDC47 was used to evaluate cellular proliferation of the epidermis and hair follicles of the mothers. Also in the mothers, serum levels of cortisol were measured by the chemiluminescence technique. Data were submitted to analysis of variance comparing mediums by Kruskall Wallis Test and SNK. In groups treated with caffeine 25 and 50mg/kg, both mothers and their puppies had focal alopecia and/or hypotrichosis. Despite the higher frequency of macroscopic changes on the mothers, these lesions were diffuse when present on the puppies. Histological analysis showed calcinosis of hair follicles in the mothers and their puppies. But morphometry revealed significant difference in the number of hair follicles from mothers, as well as a significant reduction of cell proliferation of hair follicles in the group treated with 50mg/kg of caffeine. Maternal cortisol levels were significantly elevated in the group treated with 100mg/kg of caffeine. It is concluded that caffeine intake by pregnant and lactating rats can cause skin lesions in both the mothers and their offspring, characterized by alopecia and/or hypotrichosis, regardless of serum levels of maternal cortisol.
rat; caffeine; skin; alopecia; cortisol
INTRODUÇÃO
A cafeína é uma metilxantina encontrada em cafés, chás, refrigerantes, chocolates, e medicamentos, sendo amplamente consumida pela população humana (Souza e Sichieri, 2005SOUZA, R.A.G.; SICHIERI, R. Caffeine intake and food sources of caffeine and prematurity: a case-control study. Rev. Nutr., v.21, p.1919-1928, 2005.). Em ratas gestantes, a cafeína atravessa a barreira placentária, passa para o líquido amniótico e para o sangue do cordão umbilical e é encontrada no sangue fetal em concentrações similares às do sangue materno. Por isso, o consumo materno dessa xantina é considerado um fator de risco potencial para a saúde do feto (Souza e Sichieri, 2005SOUZA, R.A.G.; SICHIERI, R. Caffeine intake and food sources of caffeine and prematurity: a case-control study. Rev. Nutr., v.21, p.1919-1928, 2005.).
A cafeína é uma substância farmacologicamente ativa, que causa diversos efeitos no organismo. Pode se incorporar ao material genético da célula, aumentando a apoptose e induzindo mutagênese (Souza e Sichieri, 2005SOUZA, R.A.G.; SICHIERI, R. Caffeine intake and food sources of caffeine and prematurity: a case-control study. Rev. Nutr., v.21, p.1919-1928, 2005.). Estudos em células trofoblásticas, em osteoblastos e em células ovarianas confirmaram seu efeito apoptótico dose-dependente (Nomura et al., 2004NOMURA, K.; SAITO, S.; IDE, K. et al. Caffeine suppresses the expression of the Bcl-2 mRNA in BeWo cell culture and rat placenta. J. Nutr. Biochem., v.6, p.342-9, 2004.; Lu et al., 2008LU, P.Z.; LAI, C.Y.; CHAN, W.H. Caffeine induces cell death via activation of apoptotic signal and inactivation of survival signal in human osteoblasts. Int. J. Mol. Sci., v.9, p.698-718, 2008.; Fernández et al., 2003FERNÁNDEZ, M.J.; LÓPEZ, A.; SANTA-MARIA, A. Apoptosis induced by different doses of caffeine on Chinese hamster ovary cells. J. Appl. Toxicol., v.23, p.221-224, 2003.).
Apesar dos efeitos nocivos, alguns estudos têm comprovado que a cafeína também pode ter efeitos benéficos. Ensaios in vitro, por exemplo, sugerem que seu uso tópico pode inibir ou estimular o crescimento de folículos pilosos, dependendo da dose (Fischer et al., 2007FISCHER, T.W.; HIPLER, U.C.; ELSNER, P. Effect of caffeine and testosterone on the proliferation of human hair follicles in vitro. Int. J. Dermatol., v.46, p.27-35, 2007.).
A aplicação tópica de cafeína em ratos após submissão à irradiação UV indica que essa substância também é capaz de diminuir o dano ao DNA, a lesão de queratinócitos e a fotocarcinogênese (Koo et al., 2007KOO, S.W.; HIRAKAWA, S.; FUJJI, S. et al. Protection from photodamage by topical application of caffeine after ultraviolet irradiation. Br. J. Dermatol., v.156, p.957-964, 2007. ; Cooney et al., 2008COONEY, A.H.; KRAMATA, P.; LOU, Y.P. Effect of caffeine on UVB-induced carcinogenisis, apoptosis, and the elimination of UVB-induced patches of p53 mutant epidermal cells in SKH-1 mice. Photochem. Photobiol., v.84, p.330-338, 2008.; Lou et al., 2010LOU, Y.; PENG, Q.; NOLAN, B. et al. Oral administration of caffeine during voluntary exercise markedly decrease tissue fat and stimulates apoptosis and cyclin B1 in UVB treated skin of hairless p53- knockout mice. Carcinogenesis, v.31, p.671-678, 2010.; Szeto e Tong, 2010SZETO, Y.T.; TONG, H.H. Caffeine as a photoprotective agent for diminishing phototoxicity. Toxicol. Ind. Health., v.10, p.667-70, 2010.).
Entretanto, o consumo de cafeína materna pode causar defeitos congênitos nos fetos de ratas e reduzir o peso e o crescimento ósseo dos filhotes. Alopecia também pode ser observada nas mães e nos filhotes (Reis et al., 2012REIS, A.M.S.; BATISTA, A.C.M.; OCARINO, N.M.; SERAKIDES, R. Effects of caffeine intake in mothers on maternal cortisol levels and offspring endochondral ossification. Actual. Osteol., v. 10, p. 20-36, 2014.). Mas este parece ser o primeiro estudo que investiga as alterações cutâneas de ratas-mães e filhotes causadas pela ingestão materna de cafeína durante a gestação e a lactação e sua relação com os níveis séricos de cortisol materno.
MATERIAL E MÉTODOS
Os procedimentos técnicos experimentais deste estudo foram aprovados pelo Comitê de Ética em Experimentação Animal (CETEA) da Universidade Federal de Minas Gerais - UFMG - processo nº. 177/2010, e estão de acordo com os Princípios Éticos da Experimentação Animal adotados no Brasil pela Sociedade Brasileira de Ciência Animal (SBCAL).
Foram utilizadas 24 ratas Wistar com dois meses de idade, provenientes do Biotério do Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais (UFMG). Os animais foram alojados em caixas plásticas (seis animais/caixa), onde receberam a mesma ração comercial Nuvilab(r) (1,4% de cálcio, 0,60% de fósforo e 22% de proteína) e água ad libitum. As ratas foram mantidas em regime de 12 horas de luz e 12 horas de escuro, em temperatura ambiente média de 25ºC.
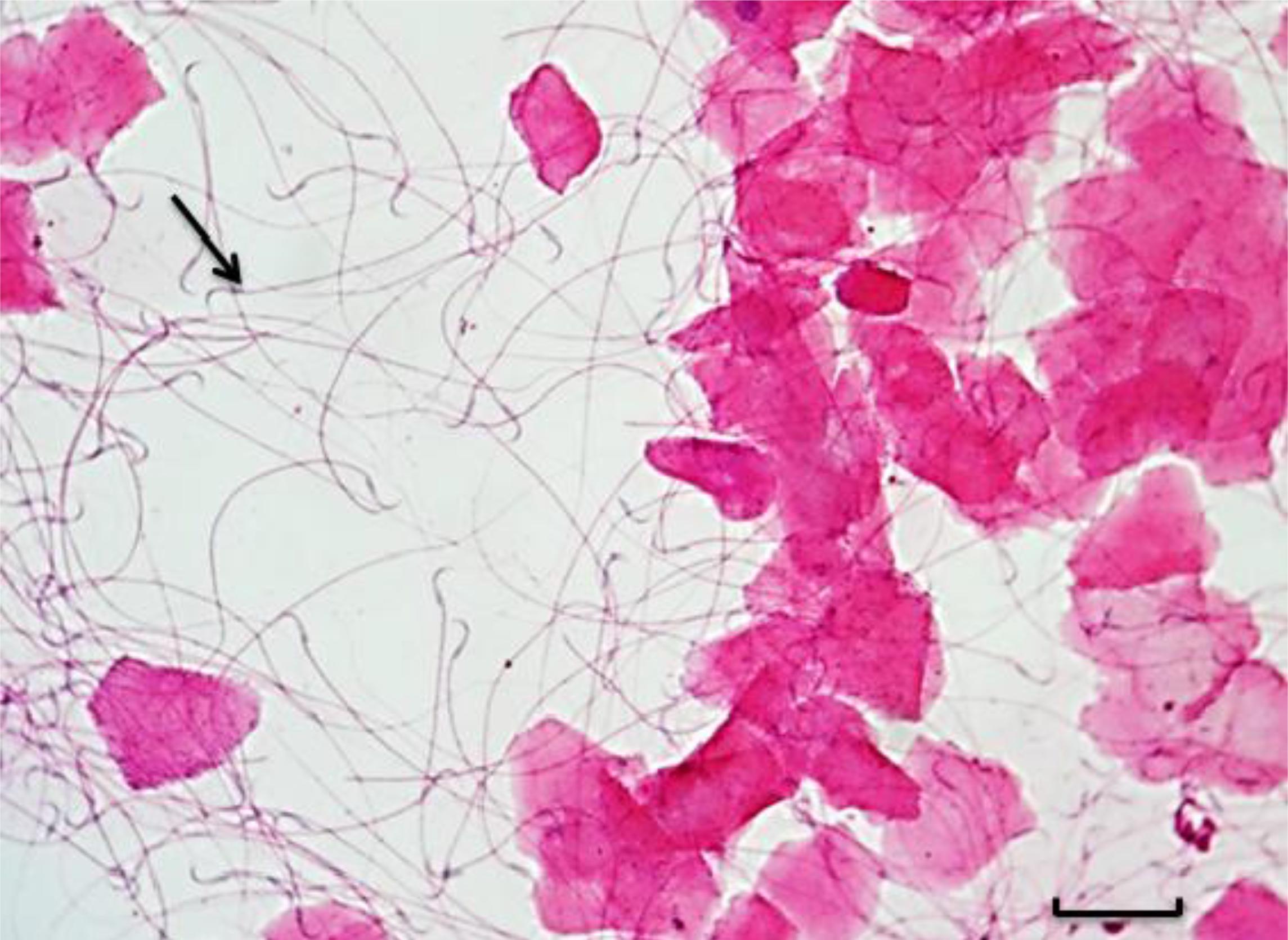
Após um período de 30 dias de adaptação, as ratas foram separadas ao acaso em quatro grupos, sendo três grupos tratados com diferentes doses de cafeína (Sigma-Aldrich, St. Louis, MO, USA) e um grupo controle (seis animais/grupo). Os grupos tratados receberam, por sonda orogástrica, cafeína diluída em 5mL de água destilada, nas doses de 25, 50 e 100mg/kg, que foram administradas uma vez ao dia, no mesmo horário, por todo o período experimental (gestação e lactação). Os animais do grupo controle receberam apenas 5mL de água destilada como placebo, também administrada por sonda orogástrica. Todas as fêmeas foram submetidas à citologia vaginal diária para determinação da fase do ciclo estral (Marcondes et al., 2002MARCONDES, F.K; BIANCHI, F.J.; TANNO, A.P. Determination of the estrous cycle phases of rats: some helpful considerations. Braz. J. Biol., v.62, p.609-614, 2002.). As ratas na fase de proestro foram alojadas em caixas plásticas com machos adultos por 12 horas, na proporção de quatro fêmeas para um macho. Após 12 horas, foram realizados esfregaços vaginais para visualização de espermatozoides e confirmação da cópula (Fig. 1). Após a cópula, as ratas foram alojadas separadamente em caixas plásticas. Os animais dos grupos tratado e controle continuaram recebendo cafeína e água destilada, respectivamente, por sonda orogástrica, durante a gestação e a lactação, por um período de 41 dias.
Fotomicroscopia da citologia vaginal das ratas na fase de estro. Seta indica a presença de espermatozoides, confirmando a cópula. Bar=9,45µm.
Transcorrido o período de lactação (21 dias), no período da manhã, as ratas foram anestesiadas com cloridrato de quetamina (40mg/kg) e cloridrato de xilazina (10mg/kg), para colheita de sangue por punção intracardíaca O sangue foi colhido em tubos para obtenção do soro e armazenado a -20ºC para a dosagem do cortisol (França et al., 2006FRANÇA, S.C.A.; NETO, T.L.B.; AGRESTA, M.C. et al. Resposta divergente da testosterona e do cortisol séricos em atletas masculinos após uma corrida de maratona. Arq. Bras. Endocrinol. Metab., v. 50, p. 1082-1087, 2006.). A dosagem de cortisol foi realizada pela técnica de quimioluminescência (Access Immunoassay System, Sanofi Diagnostics Pasteur Inc., Chaska, MN, USA), em sistema totalmente automático e de acordo com as recomendações do fabricante do kit.
Ao desmame, ou seja, 21 dias após o parto, as mães e os filhotes foram eutanasiados para colheita de fragmentos de pele para exame histopatológico. Foram avaliados três filhotes por mãe/grupo. A eutanásia foi realizada por punção cardíaca, precedida por sobredose dos anestésicos cloridrato de quetamina (40mg/kg) e cloridrato de xilazina (10mg/kg), ambos por via intraperitoneal (Reis et al., 2014REIS, A.M.S.; BATISTA, A.C.M.; OCARINO, N.M.; SERAKIDES, R. Effects of caffeine intake in mothers on maternal cortisol levels and offspring endochondral ossification. Actual. Osteol., v. 10, p. 20-36, 2014.).
Nas mães, foram colhidos dois fragmentos de pele, sendo um da região dorsocervical e o outro da região toracoventral. Nos filhotes, foi colhido um fragmento de pele da região dorsocervical. Os fragmentos foram fixados em formaldeído a 10% e submetidos ao processamento histológico de rotina com inclusão em parafina. Secções de 4µm foram coradas pela técnica de hematoxilina-eosina (HE) para análise histomorfométrica.
Foi quantificado o número de folículos pilosos com uma camada, duas camadas e com mais de duas camadas de células, de acordo com a classificação de Brenner (Brenner et al., 2006BRENNER, F.M.; SOUZA, F.M.; NETO, F. et al. Avaliação quantitativa em cortes histológicos transversais do couro cabeludo. An. Bras. Dermatol., v.81, p.227-232, 2006.). Essa quantificação foi realizada em 20 campos aleatórios, com objetiva de 40x. A espessura da epiderme foi determinada com auxílio de uma ocular micrométrica e objetiva de 40x, em 30 pontos aleatórios (Reis et al., 2014REIS, A.M.S.; BATISTA, A.C.M.; OCARINO, N.M.; SERAKIDES, R. Effects of caffeine intake in mothers on maternal cortisol levels and offspring endochondral ossification. Actual. Osteol., v. 10, p. 20-36, 2014.) (Fig. 2). Foi estabelecida uma média dos 30 pontos, e, em seguida, os valores foram convertidos para micrômetros, de acordo com a escala de uma lâmina micrométrica.
Fotomicroscopia da pele de ratas controle ilustrando a gratícula de 121 pontos usada para análise histomorfométrica. HE, Bar = 9,45µm.
Secções histológicas de 4μm de espessura em lâminas silanizadas foram submetidas à análise imunoistoquímica utilizando o anticorpo anti-CDC47 (47DC141, Neomarkers, Fremont, CA, USA) para estimativa da proliferação celular da epiderme e dos folículos pilosos. O anticorpo CDC-47 é capaz de marcar as células em desenvolvimento em diferentes órgãos, inclusive na pele, e a sua expressão determina a taxa de proliferação celular de um tecido (Leite et al., 2007LEITE, E.D.; FREITAS, E.S.; SOUZA, C..A. et al. Proliferação, apoptose e histomorfometria da glândula mamária de ratas tratadas com tiroxina na lactação e ao desmame e desenvolvimento dos filhotes. Arq. Bras. Endocrinol. Metab., v.51, p.1000-1006, 2007.). Foi utilizada a técnica da estreptavidina-biotina-peroxidase (Streptavidin Peroxidase, Lab Vision Corp., Fremont, CA, USA) e recuperação antigênica por calor em banho-maria a 98oC. As lâminas foram incubadas em câmara úmida overnight, com o anticorpo primário e por 30 minutos, nas etapas de bloqueio da peroxidase endógena, soro de bloqueio (Ultra vision Block, Lab Vision Corp., Fremont, CA. USA) e estreptavidina peroxidase. O anticorpo primário anti-CDC47 teve como diluição 1:50. A incubação com anticorpo secundário (Biotin Goat, Lab Vision Corp., Fremont, CA. USA) foi realizada por 45 minutos. O cromógeno utilizado foi a diaminobenzidina (DAB Substrate system, Lab Vision Corp., Fremont, CA. USA) com incubação por 30 minutos. As secções foram contracoradas com hematoxilina de Mayer. O controle negativo foi obtido pela substituição do anticorpo primário por IgG. Em um total de 100 células da epiderme e dos folículos pilosos de cada animal, foi determinada a porcentagem de células negativas e positivas quanto à expressão de CDC47. A expressão positiva era confirmada pela presença de células com núcleo de coloração acastanhada.
Os animais foram distribuídos de forma aleatória, em quatro grupos com seis repetições. Os dados foram submetidos à análise de variância e à verificação da distribuição de normalidade. Para cada variável estudada, foi determinada a média e o desvio-padrão. As médias do número de folículos pilosos, do número de células com expressão de CDC47 e das concentrações séricas do cortisol materno foram comparadas pelo teste Student-Newman-Keuls (SNK). As médias da espessura da epiderme foram avaliadas pelo teste Kruskal-Wallis (Sampaio, 1998SAMPAIO, I.B.B. Estatística aplicada à experimentação animal. Belo Horizonte: FEP/MVZ, 1998. 264p.). Diferenças foram consideradas significativas se P<0,05.
RESULTADOS
Os grupos tratados com doses menores de cafeína (25 e 50mg/kg) apresentaram níveis séricos de cortisol semelhantes ao do grupo controle (P≥0,05). Mas as concentrações de cortisol das mães do grupo tratado com cafeína na dose de 100mg/kg foram significativamente maiores do que as do controle (Tab. 1).
A pele das mães e a dos filhotes tratados com cafeína apresentaram alterações macroscópicas que variavam de alopecia parcial (hipotricose) focal, multifocal ou moderada a alopecia focal. A região mais afetada da pele foi o dorso do animal, e a frequência dessas alterações foi maior nas mães do que nos filhotes, apesar de as lesões terem sido mais localizadas nas mães e mais difusas nos filhotes (Fig. 3).
Fotomacroscopia dos animais afetados demonstrando áreas de alopecia periauricular focal e hipotricose focal nas mães dos grupos tratados com 25 e 50mg/kg de cafeína, respectivamente, em comparação com o grupo controle. Áreas de hipotricose dorsal difusa nos filhotes de 21 dias das ratas do grupo tratado com 25mg/kg de cafeína em comparação com o grupo controle.
A análise macroscópica da pele das mães demonstrou que o grupo mais afetado foi o tratado com 25mg/kg de cafeína, no qual 70% das ratas apresentaram lesões. Já no grupo tratado com cafeína 50mg/kg, somente 30% das mães apresentaram lesões cutâneas. As mães dos grupos controle e tratado com cafeína na dose de 100mg/kg não apresentaram alterações macro ou microscópica. As lesões das mães afetadas apresentaram diferentes localizações anatômicas (Tab. 2) e variaram quanto à intensidade (Tab. 3).
Distribuição das lesões cutâneas quanto à região afetada nas ratas-mães submetidas à administração de diferentes doses de cafeína
Distribuição das lesões cutâneas quanto à intensidade na pele das ratas-mães submetidas à administração de diferentes doses de cafeína
Considerando os filhotes, 6% foram afetados no grupo de mães tratadas com 25mg/kg, 3% foram acometidos no grupo tratado com 50mg/kg e 6% foram acometidos no grupo que recebeu 100mg/kg de cafeína, sendo as lesões dos filhotes do grupo tratado com cafeína na dose de 100mg/kg discretas e focais. Os filhotes das mães do grupo controle não apresentaram alterações macro ou microscópica. As lesões dos filhotes afetados variaram quanto à intensidade (Tab. 4) e apresentaram diferentes localizações anatômicas (Tab. 5).
Distribuição das lesões cutâneas quanto à intensidade na pele dos filhotes de ratas submetidas à administração de diferentes doses de cafeína
Distribuição das lesões quanto à área afetada na pele dos filhotes de ratas submetidas à administração de diferentes doses de cafeína
Assim como já havia sido observado à macroscopia, microscopicamente, a pele da região torácica ventral das mães não apresentou alterações significativas com relação à espessura da epiderme e ao número de folículos pilosos (Tab. 6 e Tab. 7). No entanto, na pele da região dorsal, as mães dos grupos tratados com 25 e 50mg/kg de cafeína apresentaram menor número de folículos pilosos devido a uma significativa redução do número de folículos com uma camada de células (Tab. 8). Alguns dos folículos presentes apresentavam-se sem pelos e obstruídos por ceratina, enquanto outros apresentavam placas de cálcio coradas pelo método de Von Kossa, caracterizando a calcinose (Fig. 4). Essa lesão foi observada em 20% das mães do grupo tratado com 25mg/kg e em 10% das ratas do grupo tratado com 50mg/kg. O grupo tratado com 100 mg/kg de cafeína nao apresentou calcinose de folículo piloso. A espessura da epiderme da região dorsocervical não diferiu significativamente entre grupos (Tab. 6).
A) Fotomicroscopia da pele dos filhotes de ratas do grupo tratado com 25mg/kg de cafeína, apresentando depósitos basofílicos no interior dos folículos pilosos (seta). HE, Bar=47,29µm. B) em detalhe, depósito de cálcio em negro, dentro do folículo piloso. Von Kossa, Bar=9,45µm. C, D, E e F) Fotomicroscopias da pele dorsocervical das ratas-mães demonstrando expressão nuclear de CDC47 (em marrom) semelhante na epiderme nos grupos controle e tratados com cafeína nas doses de 25, 50 e 100mg/kg, respectivamente. G, H, I e J) Fotomicroscopias da pele dorsocervical das ratas-mães demonstrando ausência de expressão de CDC47 nos folículos pilosos do grupo tratado com 50mg/kg de cafeína (C) em comparação ao controle (E) e aos demais grupos tratados com cafeína nas doses de 25mg/kg (F) e 100mg/kg (H). Imunistoquímica, contracoloração hematoxilina de Mayer. Bar = 9,45µm.
Com relação à análise microscópica da pele dos filhotes, pode-se verificar que, ao contrário do que foi observado nas mães, não houve diferença entre os grupos com relação ao número de folículos pilosos (Tab. 9). Também não houve diferença significativa com relação à espessura da epiderme (Tab. 6), mas vários animais apresentaram calcinose do folículo piloso. Essa lesão foi observada em 30% dos filhotes das mães tratadas com 25mg/kg e em 10% dos filhotes das ratas dos grupos tratados com 50mg/kg e 100mg/kg de cafeína.
Na análise imuno-histoquímica da proliferação celular, somente na pele das mães do grupo tratado com 50mg/kg de cafeina houve menor atividade proliferativa das células dos folículos pilosos, caracterizada por expressão de CDC47 significativamente menor em comparação à do grupo controle. Na epiderme, a taxa de proliferação celular foi semelhante entre os grupos (Fig.4, Tab.10).
DISCUSSÃO
As concentrações séricas de cortisol das mães aos 21 dias de lactação foram maiores somente no grupo tratado com cafeína na dose de 100mg/kg em comparação ao grupo controle. Outros estudos em ratos também revelaram que a cafeína em doses baixas ou moderadas não promove a elevação da corticosterona, ao contrário de doses elevadas (Tan et al., 2012TAN,Y.; LIU, J.; DENG, Y. et al. Caffeine-induced fetal rat over-exposure to maternal glucocorticoid and histone methylation of liver IGF-1 might cause skeletal growth retardation. Toxicol. Let., v.214, p.279-287, 2012.). A elevação do cortisol pode ser explicada pelo fato de a cafeína, por ser uma substância farmacologicamente ativa, atuar no sistema nervoso (Fisone et al., 2004FISONE, G.; BORGKVIST, A.; USIELLO, A. Caffeine as a psychomotor stimulant: mechanism of action. Cell Mol. Life Sci., v.61, p.857-872, 2004.) e, assim, ativar o eixo do estresse (Lovallo et al., 2005LOVALLO, W.R.; WHISETT, T.L.; ABSI, M. et al. Caffeine stimulation of cortisol secretion across de waking hours in relation to caffeine intake levels. Psychosom. Med., v.67, p.734-739, 2005.; Lovallo et al., 1989LOVALLO, W.R.; PINCOMB, G.A.; SUNG, B.H. et al. Caffeine may potentiate adrenocortical stress responses in hypertension-prone men. Hypertension, v.14, p.170-6, 1989.), elevar os níveis de catecolaminas e, consequentemente, dos glicocorticoides. Além disso, a cafeína aumenta tanto a secreção de ACTH quanto de cortisol por agir diretamente no hipotálamo ou por estimular indiretamente a liberação de epinefrina pela adrenal (Souza e Sichieri, 2005SOUZA, R.A.G.; SICHIERI, R. Caffeine intake and food sources of caffeine and prematurity: a case-control study. Rev. Nutr., v.21, p.1919-1928, 2005.; Kimyai-Asadi e Usman, 2001KIMYAI-ASADI, A.; USMAN, A. The role of psychological stress in skin disease. J. Cutan. Med. Surg., v.5, p.140-145, 2001.; Boghian et al., 2008BOGHIAN, V.; SOLCAN, G.; HRITCU, L.D. et al. Clinical and paraclinical aspects in Cushing syndrome (hyperadrenocorticism) in dogs. Lucrari Stiint., v.41, p.136-142, 2008.). Por extensão, as alterações no cortisol decorrentes do consumo diário de cafeína podem ter implicações na saúde (Fisone et al., 2004FISONE, G.; BORGKVIST, A.; USIELLO, A. Caffeine as a psychomotor stimulant: mechanism of action. Cell Mol. Life Sci., v.61, p.857-872, 2004.). Elevados níveis sanguíneos de cortisol podem causar ou exacerbar lesões cutâneas simétricas, como alopecia, ressecamento, adelgaçamento da pele, redução da elasticidade, ptose abdominal, hiperpigmentação e calcificação cutânea (Roy et al., 1996ROY, A.; LEBLANC, C.; PAQUETTE, L. et al. Skin bruising in asthmatic subjects treated with high doses of inhaled steroids: frequency and association with adrenal function. Eur. Respir J., v.9, p.226-231, 1996.; Kimyai-Asadi e Usman, 2001KIMYAI-ASADI, A.; USMAN, A. The role of psychological stress in skin disease. J. Cutan. Med. Surg., v.5, p.140-145, 2001.; Boghian et al., 2008BOGHIAN, V.; SOLCAN, G.; HRITCU, L.D. et al. Clinical and paraclinical aspects in Cushing syndrome (hyperadrenocorticism) in dogs. Lucrari Stiint., v.41, p.136-142, 2008.). Elevados níveis de cortisol nas mães podem, também, passar para os filhotes através da placenta e do leite (Swolin-eide et al., 2002SWOLIN-EIDE, D.; DAHLGREN, J.; NILSSON, C. et al. Affected skeletal growth but normal bone mineralization in rat offspring after prenatal dexametasone exposure. J. Endocrinol., v.174, p.411-418, 2002.; Glynn et al., 2007GLYNN, L.M.; DAVIS, E.P.; SHCETERR, C.D. et al. Postnatal maternal cortisol levels predict temperament in healthy breastfed infants. Early Hum. Develop., v.83, p.675-81, 2007.). O estresse experimental pode ser excluído como um possível interferente neste trabalho, pois os animais dos grupos tratados e controle foram submetidos aos mesmos procedimentos experimentais. No presente estudo, as lesões cutâneas das mães e dos filhotes, caracterizadas por hipotricose e/ou alopecia associadas à calcinose dos folículos pilosos, ocorreram independentemente dos níveis séricos do cortisol materno, uma vez que a maior ocorrência dessas lesões foi observada nos animais tratados com cafeína nas doses de 25 e 50mg/kg, cujas mães não apresentaram elevação dos níveis de cortisol.
Neste estudo, as ratas-mães do grupo tratado com 100mg/kg de cafeína não apresentaram lesões, ao contrário dos animais dos grupos tratados com 25 e 50mg/kg, o que pode ser justificado pela relação dose-dependente e pela tolerância à cafeína. Alguns estudos demonstram que o consumo diário dessa metilxantina pode causar tolerância (Kendler e Prescott, 1999KENDLER, K.S.; PRESCOTT, C.A. Caffeine intake, tolerance, and withdrawal in women: a population-base twin study. Am. J. Psychiatry., v.156, p.223-228, 1999.), que é rapidamente instalada após o acúmulo de cafeína e de seus metabólitos no organismo (Kendler e Prescott, 1999KENDLER, K.S.; PRESCOTT, C.A. Caffeine intake, tolerance, and withdrawal in women: a population-base twin study. Am. J. Psychiatry., v.156, p.223-228, 1999.).
Há pesquisadores que relatam que os níveis sanguíneos materno e fetal de cafeína são semelhantes durante a gestação (Souza e Sichieri, 2005SOUZA, R.A.G.; SICHIERI, R. Caffeine intake and food sources of caffeine and prematurity: a case-control study. Rev. Nutr., v.21, p.1919-1928, 2005.) e que, nos recém-nascidos, o metabolismo e a taxa de depuração da cafeína estão diminuídos (Tavares e Sakata, 2012TAVARES, C.; SAKATA, R.K. Cafeína para o tratamento de dor. Rev. Bras. Anestesiol., v.62, p.387-401, 2012.). No entanto, a despeito disso, a frequência das alterações foi maior nas mães quando comparadas aos filhotes. No entanto, quando presentes nos filhotes, as lesões foram mais intensas. Uma hipótese que poderia justificar o fato de as lesões serem menos frequentes nos filhotes após o desmame seria a de que, após esse período, os filhotes são menos susceptíveis à ação da cafeína, uma vez que já estão formados os sistemas microssomais do fígado envolvendo o citocromo P450. Outra hipótese poderia ser a de que a cafeína consumida pela mãe possa ser menos veiculada pelo leite do que pela placenta (Neal e Benowitz, 1990NEAL, L; BENOWITZ, M.D. Clinical pharmacology of caffeine. Ann. Rev. Med., v.41, p.277-288, 1990.).
Macroscopicamente, as lesões apresentaram localizações variadas. Porém, a região dorsal foi mais afetada em comparação às regiões ventrais. Uma explicação para esse resultado poderia ser o maior número de folículos pilosos nesta região, uma vez que a alopecia foi a lesão macroscópica evidenciada.
A calcificação dos folículos pilosos observada nos filhotes e nas mães, especialmente nos grupos tratados com 25 e 50mg/kg de cafeína, caracteriza a calcinosis cutis, que pode ser distrófica, metastática ou iatrogênica (Boghian et al., 2008BOGHIAN, V.; SOLCAN, G.; HRITCU, L.D. et al. Clinical and paraclinical aspects in Cushing syndrome (hyperadrenocorticism) in dogs. Lucrari Stiint., v.41, p.136-142, 2008.). O excesso de cortisol pode causar depósito ectópico de cálcio em tecidos moles, como a pele e a adrenal. Na pele, esse mineral causa efeitos maléficos, diminuindo a proliferação, a diferenciação e a adesão celular (Aszterbaum, 1993ASZTERBAUM, M.; FEINGOLD, K.R.; MENON, G.K.; WILLIAMS, M.L. Glucocorticoids accelerate fetal maturation of the epidermal permeability barrier in the rat. J. Clin. Invest., v.91, p.2703-08, 1993.). O hiperadrenocorticismo é uma das causas de calcinose cútis (Boghian et al., 2008BOGHIAN, V.; SOLCAN, G.; HRITCU, L.D. et al. Clinical and paraclinical aspects in Cushing syndrome (hyperadrenocorticism) in dogs. Lucrari Stiint., v.41, p.136-142, 2008.). Entretanto, neste estudo, os grupos de animais com calcinose não apresentaram elevação do cortisol, o que afasta a possibilidade de esse hormônio estar envolvido na gênese da calcificação dos folículos pilosos decorrente do consumo de cafeína. Neste caso, a calcificação pode ser explicada pelo fato de a cafeína ativar os canais de cálcio sensíveis à rianodina, levando à liberação de cálcio intracelular (Tavares e Sakata, 2012TAVARES, C.; SAKATA, R.K. Cafeína para o tratamento de dor. Rev. Bras. Anestesiol., v.62, p.387-401, 2012.).
As diferenças encontradas na proliferação celular dos folículos pilosos das mães tratadas e controle podem ser explicadas pelos efeitos da cafeína no material genético. A possível incorporação de cafeína no material genético pode alterar a replicação celular, diminuindo a fase G2 da mitose (Souza e Sichieri, 2005SOUZA, R.A.G.; SICHIERI, R. Caffeine intake and food sources of caffeine and prematurity: a case-control study. Rev. Nutr., v.21, p.1919-1928, 2005.), o que resultaria em menor proliferação celular. Além disso, a cafeína pode aumentar o metabolismo do folículo piloso, resultando em consumo excessivo de reservas energéticas, com exaustão da capacidade proliferativa (Fischer et al., 2007FISCHER, T.W.; HIPLER, U.C.; ELSNER, P. Effect of caffeine and testosterone on the proliferation of human hair follicles in vitro. Int. J. Dermatol., v.46, p.27-35, 2007.). Experimentos in vitro com baixas concentrações de cafeína (0,05% e 0,01%) demonstraram supressão do crescimento folicular, enquanto concentrações superiores, de 0,15%, mostraram-se estimulatórias e têm sido adicionadas ao tratamento local da calvície em humanos (Fischer et al., 2007FISCHER, T.W.; HIPLER, U.C.; ELSNER, P. Effect of caffeine and testosterone on the proliferation of human hair follicles in vitro. Int. J. Dermatol., v.46, p.27-35, 2007.). Embora existam estudos que analisaram os efeitos in vitro da cafeína sobre folículos pilosos (Fischer et al., 2007FISCHER, T.W.; HIPLER, U.C.; ELSNER, P. Effect of caffeine and testosterone on the proliferation of human hair follicles in vitro. Int. J. Dermatol., v.46, p.27-35, 2007.), o efeito da cafeína in vivo no modelo utilizado neste estudo parece ter sido a primeira tentativa de se estudarem os efeitos da cafeína na pele de ratas gestantes e de sua prole.
CONCLUSÃO
A cafeína ingerida pelas ratas gestantes e lactantes pode causar lesões cutâneas tanto nas mães quanto nos filhotes, caracterizadas por hipotricose e alopecia associadas à calcinose dos folículos pilosos, independentemente dos níveis séricos do cortisol materno.
AGRADECIMENTOS
Este trabalho foi financiado por recursos da Fundação de Amparo à Pesquisa de Minas Gerais (Fapemig) e do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
REFERÊNCIAS
- ASZTERBAUM, M.; FEINGOLD, K.R.; MENON, G.K.; WILLIAMS, M.L. Glucocorticoids accelerate fetal maturation of the epidermal permeability barrier in the rat. J. Clin. Invest., v.91, p.2703-08, 1993.
- BOGHIAN, V.; SOLCAN, G.; HRITCU, L.D. et al. Clinical and paraclinical aspects in Cushing syndrome (hyperadrenocorticism) in dogs. Lucrari Stiint., v.41, p.136-142, 2008.
- BRENNER, F.M.; SOUZA, F.M.; NETO, F. et al. Avaliação quantitativa em cortes histológicos transversais do couro cabeludo. An. Bras. Dermatol., v.81, p.227-232, 2006.
- COONEY, A.H.; KRAMATA, P.; LOU, Y.P. Effect of caffeine on UVB-induced carcinogenisis, apoptosis, and the elimination of UVB-induced patches of p53 mutant epidermal cells in SKH-1 mice. Photochem. Photobiol., v.84, p.330-338, 2008.
- FERNÁNDEZ, M.J.; LÓPEZ, A.; SANTA-MARIA, A. Apoptosis induced by different doses of caffeine on Chinese hamster ovary cells. J. Appl. Toxicol., v.23, p.221-224, 2003.
- FISCHER, T.W.; HIPLER, U.C.; ELSNER, P. Effect of caffeine and testosterone on the proliferation of human hair follicles in vitro. Int. J. Dermatol., v.46, p.27-35, 2007.
- FISONE, G.; BORGKVIST, A.; USIELLO, A. Caffeine as a psychomotor stimulant: mechanism of action. Cell Mol. Life Sci., v.61, p.857-872, 2004.
- FRANÇA, S.C.A.; NETO, T.L.B.; AGRESTA, M.C. et al. Resposta divergente da testosterona e do cortisol séricos em atletas masculinos após uma corrida de maratona. Arq. Bras. Endocrinol. Metab., v. 50, p. 1082-1087, 2006.
- GLYNN, L.M.; DAVIS, E.P.; SHCETERR, C.D. et al. Postnatal maternal cortisol levels predict temperament in healthy breastfed infants. Early Hum. Develop., v.83, p.675-81, 2007.
- KENDLER, K.S.; PRESCOTT, C.A. Caffeine intake, tolerance, and withdrawal in women: a population-base twin study. Am. J. Psychiatry., v.156, p.223-228, 1999.
- KIMYAI-ASADI, A.; USMAN, A. The role of psychological stress in skin disease. J. Cutan. Med. Surg., v.5, p.140-145, 2001.
- KOO, S.W.; HIRAKAWA, S.; FUJJI, S. et al. Protection from photodamage by topical application of caffeine after ultraviolet irradiation. Br. J. Dermatol., v.156, p.957-964, 2007.
- LEITE, E.D.; FREITAS, E.S.; SOUZA, C..A. et al. Proliferação, apoptose e histomorfometria da glândula mamária de ratas tratadas com tiroxina na lactação e ao desmame e desenvolvimento dos filhotes. Arq. Bras. Endocrinol. Metab., v.51, p.1000-1006, 2007.
- LU, P.Z.; LAI, C.Y.; CHAN, W.H. Caffeine induces cell death via activation of apoptotic signal and inactivation of survival signal in human osteoblasts. Int. J. Mol. Sci., v.9, p.698-718, 2008.
- LOU, Y.; PENG, Q.; NOLAN, B. et al. Oral administration of caffeine during voluntary exercise markedly decrease tissue fat and stimulates apoptosis and cyclin B1 in UVB treated skin of hairless p53- knockout mice. Carcinogenesis, v.31, p.671-678, 2010.
- LOVALLO, W.R.; PINCOMB, G.A.; SUNG, B.H. et al. Caffeine may potentiate adrenocortical stress responses in hypertension-prone men. Hypertension, v.14, p.170-6, 1989.
- LOVALLO, W.R.; WHISETT, T.L.; ABSI, M. et al. Caffeine stimulation of cortisol secretion across de waking hours in relation to caffeine intake levels. Psychosom. Med., v.67, p.734-739, 2005.
- MARCONDES, F.K; BIANCHI, F.J.; TANNO, A.P. Determination of the estrous cycle phases of rats: some helpful considerations. Braz. J. Biol., v.62, p.609-614, 2002.
- NEAL, L; BENOWITZ, M.D. Clinical pharmacology of caffeine. Ann. Rev. Med., v.41, p.277-288, 1990.
- NOMURA, K.; SAITO, S.; IDE, K. et al. Caffeine suppresses the expression of the Bcl-2 mRNA in BeWo cell culture and rat placenta. J. Nutr. Biochem., v.6, p.342-9, 2004.
- REIS, A.M.S.; BATISTA, A.C.M.; OCARINO, N.M.; SERAKIDES, R. Effects of caffeine intake in mothers on maternal cortisol levels and offspring endochondral ossification. Actual. Osteol., v. 10, p. 20-36, 2014.
- ROY, A.; LEBLANC, C.; PAQUETTE, L. et al. Skin bruising in asthmatic subjects treated with high doses of inhaled steroids: frequency and association with adrenal function. Eur. Respir J., v.9, p.226-231, 1996.
- SAMPAIO, I.B.B. Estatística aplicada à experimentação animal. Belo Horizonte: FEP/MVZ, 1998. 264p.
- SOUZA, R.A.G.; SICHIERI, R. Caffeine intake and food sources of caffeine and prematurity: a case-control study. Rev. Nutr., v.21, p.1919-1928, 2005.
- SWOLIN-EIDE, D.; DAHLGREN, J.; NILSSON, C. et al. Affected skeletal growth but normal bone mineralization in rat offspring after prenatal dexametasone exposure. J. Endocrinol., v.174, p.411-418, 2002.
- SZETO, Y.T.; TONG, H.H. Caffeine as a photoprotective agent for diminishing phototoxicity. Toxicol. Ind. Health., v.10, p.667-70, 2010.
- TAN,Y.; LIU, J.; DENG, Y. et al. Caffeine-induced fetal rat over-exposure to maternal glucocorticoid and histone methylation of liver IGF-1 might cause skeletal growth retardation. Toxicol. Let., v.214, p.279-287, 2012.
- TAVARES, C.; SAKATA, R.K. Cafeína para o tratamento de dor. Rev. Bras. Anestesiol., v.62, p.387-401, 2012.
Datas de Publicação
-
Publicação nesta coleção
Jul-Aug 2015
Histórico
-
Recebido
20 Ago 2013 -
Aceito
13 Mar 2015