RESUMO
Este estudo teve por objetivo estimar os parâmetros genéticos de características de interesse econômico, mensuradas em populações F2 desenvolvidas pela Embrapa Suínos e Aves utilizando cruzamento recíproco entre linhagens de corte e de postura. Ainda, foram avaliados os efeitos de sexo e de cruzamento recíproco sobre as características em estudo. Os pesos com um, 35 e 42 dias de idade; ganho de peso; consumo de ração e conversão alimentar entre 35 e 41 dias de idade; pesos dos pulmões, fígado, coração, moela, peito, pernas, carcaça, dorso, asas, cabeça, pés e gordura abdominal, além do comprimento do intestino, foram os fenótipos estudados. Foram estimados os componentes de variâncias genética aditiva e residual, além dos coeficientes de herdabilidade e das correlações genética e fenotípica. Os machos apresentaram maior peso para todas as características estudadas, nos dois cruzamentos recíprocos, exceto para gordura abdominal na população oriunda do cruzamento de machos de postura com fêmeas de corte. Os animais oriundos do cruzamento de machos de postura com fêmeas de corte foram mais pesados que os recíprocos, para todas as idades, além de apresentarem maior comprimento de intestino e maiores pesos de moela, carcaça, dorso, peito e cabeça. Os coeficientes de herdabilidade foram altos para consumo de ração e para os pesos ao nascimento, da moela e da gordura abdominal. As correlações fenotípicas estimadas foram, em sua maioria, baixas ou moderadas, contudo muitas correlações genéticas altas foram observadas. Ressalta-se que houve expressiva diferença nos coeficientes de herdabilidade de algumas características em função do cruzamento recíproco estudado, o que pode ser devido a efeitos materno, citoplasmático, ligados ao sexo ou imprinting.
Palavras-chave:
avicultura; componentes de variância; correlação genética; herdabilidade
ABSTRACT
This study aimed to estimate genetic parameters for several economic traits in F2 populations developed by Embrapa Swine and Poultry National Research Center, through reciprocal crosses of broilers and layer lines. Furthermore, sex and reciprocal cross effects were evaluated. Weights at 1, 35 and 42 days of age; weight gain, feed intake and feed conversion between 35 and 41 days of age; weights of the lungs, heart, liver, gizzard, breast, thighs, carcass, back, wings, head, legs and abdominal fat; and intestine length were studied. Residual and genetic additive variance components, heritability coefficients, and genetic and phenotypic correlations were estimated. Males were heavier than females for all studied traits in both reciprocal crosses, except for abdominal fat weight in chickens from the male layer x female broiler cross. Chickens from male layer x female broiler cross were heavier than those from its reciprocal cross, also having larger intestine length and gizzard, carcass, back, breast and head weights. The heritability coefficients were high for feed intake and body weight at birth, gizzard, and abdominal fat. The estimated phenotypic correlations were mostly lower or moderate, however, most high genetic correlations were observed. We found significant differences in heritability coefficients for some traits due to the reciprocal cross, which may be due maternal, cytoplasmic, sex-linked or imprinting effects.
Keywords:
variance components; correlation; heritability; poultry
INTRODUÇÃO
O desenvolvimento proporcional dos órgãos e a redução da gordura abdominal são alguns dos objetivos da avicultura moderna. Contudo, a seleção para essas características é difícil de ser conduzida, por elas terem herança genética complexa e serem de difícil avaliação fenotípica, em alguns casos. Assim, têm se buscado informações de marcadores moleculares para auxiliar o processo de seleção dessas características. Hocking (2005HOCKING, P. Review on QTL mapping in chickens. Poult. Sci. J. , v.61, p.215-226, 2005.) e Abasht et al. (2006ABASHT, B.; DEKKERS J.C.M.; LAMONT S.J. Review of quantitative trait loci identified in the chicken. Poult. Sci., v.85, p.2079-2096, 2006.) fizeram revisões que ilustram a importância dos estudos de mapeamento de QTLs em frangos no final do século XX e início do XXI. Muitos estudos de mapeamento de QTL foram conduzidos com populações F2 por meio de cruzamentos recíprocos, a fim de maximizar a variabilidade (Knott e Haley, 1992KNOTT, S.A.; HALEY, C.S. A simple regression method for mapping quantitative trait loci in line crosses using flanking markers. Heredity, v.69, p.315-324, 1992.). Porém, pouco se conhece sobre os parâmetros genéticos (herdabilidade e correlações genéticas) nessas populações F2. Sabe-se, contudo, que, para características de alta herdabilidade, os QTLs são identificados com mais facilidade que para aquelas de baixa herdabilidade, devido ao fato de a maior parte da variação fenotípica ser de natureza genética aditiva. Assim, conhecer a herdabilidade é o primeiro passo para saber o potencial de mapeamento de QTL para uma característica em uma determinada população. Já as correlações genéticas podem sugerir se QTLs mapeados muito próximos são múltiplos QTLs ligados (baixa correlação genética entre as características) ou se é apenas um QTL com efeito pleiotrópico sobre múltiplas características (alta correlação genética entre as características). Assim, o presente estudo teve por objetivo estimar os parâmetros genéticos para características de interesse mensuradas nas populações F2 desenvolvidas pela Embrapa, utilizando cruzamento recíproco entre linhagens de corte e de postura.
MATERIAL E MÉTODOS
Para a realização de estudos genômicos, duas linhagens desenvolvidas pela Embrapa Suínos e Aves foram utilizadas para a formação de duas populações referência. A linhagem de corte (TT) foi selecionada com o objetivo de melhorar o peso corporal, a conversão alimentar, o rendimento de carcaça e partes, a viabilidade, a fertilidade e a porcentagem de eclosão. A linhagem de postura (CC) foi selecionada para melhorar as características de produção de ovos, peso do ovo, conversão alimentar, viabilidade, maturidade sexual, fertilidade, porcentagem de eclosão, qualidade do ovo, bem como para reduzir o peso corporal. Com essas linhagens, foram realizados cruzamentos recíprocos entre machos de corte (TT) e fêmeas de postura (CC) para formar a população F1 TC e machos de postura (CC) e fêmeas de corte (TT) para formar a população F1 CT. A geração F1 constou de seis famílias de irmãos completos, cada uma contendo três machos e seis fêmeas escolhidas ao acaso. Machos e fêmeas em cada uma das populações F1 foram acasalados entre si, tendo o cuidado de se evitar o acasalamento entre indivíduos com laços de parentesco, para, assim, formar as duas gerações F2 (TCTC e CTCT). Cada população F2 foi constituída de aproximadamente 100 pintos por família de mãe, em 17 incubações, totalizando registros de 3823 animais, sendo 1760 CTCT e 2063 TCTC com dados de produção e desempenho. Foi utilizado o modelo linecross como descrito por Knott e Haley (1992KNOTT, S.A.; HALEY, C.S. A simple regression method for mapping quantitative trait loci in line crosses using flanking markers. Heredity, v.69, p.315-324, 1992.).
As características de desempenho mensuradas foram os pesos com um, 35 e 42 dias de idade, além do ganho de peso, do consumo de ração e da conversão alimentar, mensurados entre 35 e 41 dias de idade. O peso vivo aos 42 dias de idade (PV42) foi mensurado após seis horas de jejum e transporte para o abate. As características de carcaça, avaliadas após quatro horas de resfriamento, foram os pesos dos pulmões, fígado, coração, moela, peito, pernas (peso de coxas e sobrecoxas), carcaça (sem vísceras, pés e cabeça), dorso (peso da carcaça sem peito, asas e pernas), asas, cabeça, pés e gordura abdominal, além de comprimento do intestino.
O arquivo de dados de produção e desempenho incluiu registros de 3823 animais, sendo 1760 oriundos do acasalamento de machos de postura com fêmeas de corte (população CTCT) e 2063 animais oriundos do acasalamento de machos de corte com fêmeas de postura (população TCTC). Para a estimação dos parâmetros genéticos, foram realizadas análises multicaracterísticas separadas entre as populações. Para se estimarem os parâmetros genéticos e fenotípicos, pelo método REML, foi utilizado o programa VCE-6 (Groeneveld, 2008GROENEVELD, E.; KOVAC, M.; MIELENZ, N. VCE user's guide and reference manual. Version 6.0. Urbana, IL: University of Illinois/ Department of Animal Science, 2008.), em que as variáveis consideradas foram divididas em quatro grupos funcionalmente similares, da seguinte forma: grupo 1 - desempenho (ganho de peso, consumo de ração e conversão alimentar); grupo 2 - pesos (peso ao nascimento, peso vivo aos 35 dias e peso vivo aos 42 dias); grupo 3 - órgãos (pesos do fígado, coração, moela, pulmões, gordura abdominal e comprimento do intestino); e grupo 4 - cortes (pesos das asas, pernas, peito, pés, cabeça, dorso e carcaça).
Para todas as características, foram incluídos no modelo estatístico os efeitos fixos de sexo e incubação. Nas variáveis de consumo, eficiência e ganho de peso entre 35 e 41 dias, foi inserida no modelo a covariável peso vivo aos 35 dias de idade e, para as características analisadas pós-abate, foi inserido no modelo a covariável peso vivo aos 42 dias de idade. Os efeitos aleatórios de animal e de mãe também foram incluídos no modelo. Em termos matriciais, o modelo pode ser descrito como segue:
y = μ + bX + aZ+ mW + e,
em que: y é o vetor das n observações de uma determinada característica; μ é o vetor de médias das características em estudo; b é o vetor dos efeitos fixos de sexo (macho e fêmea), população (TCTC e CTCT) e incubação (17 níveis); X é uma matriz quadrada, de ordem N, que relaciona as observações de y com os respectivos efeitos fixos; a é o vetor dos efeitos genéticos aditivos diretos preditos; Z é uma matriz quadrada, de ordem N, que relaciona as observações de y com os respectivos efeitos genéticos aditivos dos animais, contidos em a; m é o vetor dos efeitos genéticos aditivos maternos preditos; W é uma matriz quadrada, de ordem N, que relaciona as observações de y com os respectivos efeitos genéticos aditivos maternos, contidos em a; e é o vetor de efeitos residuais aleatórios.
A distribuição conjunta dos efeitos aleatórios pode ser descrita pela seguinte relação:

em que: 2
m = variância genética aditiva materna; σ2
e = variância residual; I = matriz identidade de ordem N; N = número de matrizes com observações. = variância genética aditiva direta; A = matriz dos coeficientes de parentesco, de Wright, entre os animais; σ
= variância genética aditiva direta; A = matriz dos coeficientes de parentesco, de Wright, entre os animais; σ
RESULTADOS E DISCUSSÕES
Foram observadas diferenças entre os sexos (P<0,001) para todas as características estudadas, nos dois cruzamentos avaliados. Os machos foram mais pesados que as fêmeas e também apresentaram melhor conversão alimentar e maior comprimento de intestino. O efeito de cruzamento recíproco também foi observado para algumas características. Para peso vivo, os animais da população CTCT foram mais pesados que TCTC, tanto ao nascimento quanto aos 35 e 42 dias (Tab. 1). Assim, utilizar os cruzamentos envolvendo machos da linha CC com fêmeas da linha TT parece resultar em maior desenvolvimento dos animais da geração F2. Ledur et al. (2000LEDUR, M.C.; ZANELLA, E.L.; SCHMIDT, G.S. et al. Peso e características de carcaça em linhagens utilizadas no desenvolvimento de populações referência para detecção de QTLS em aves. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 2000, Campinas. Anais... Campinas: Fundação Apinco de Ciência e Tecnologia Avícolas, 2000. 1v. CD-ROM. ) já haviam observado esse efeito sobre a geração F1 desses cruzamentos e concluíram que a fêmea teve maior impacto que os machos na formação do peso corporal das progênies, possivelmente devido ao maior peso do ovo da linhagem TT. Contudo, para maiores explicações, fazem-se necessários estudos de fatores, como os efeitos de imprinting, citoplasmático, ligados ao sexo e materno. O efeito de imprinting genômico ainda não tem sua função totalmente elucidada, mas já se sabe que a expressão de um gene ou região no cromossomo pode variar em função da origem materna ou paterna. Ambos alelos, materno ou paterno, estão presentes, mas um grupo torna-se ativo e outro é silenciado (Hitchins E Moore, 2002HITCHINS, M.P.; MOORE, G.E. Genomic imprinting in fetal growth and development. Expert. Rev. Mol. Med., v.9, p.1-19, 2002.). Contudo, imprinting ainda não é um efeito comprovado em aves, e as populações desenvolvidas pela Embrapa podem ser boas opções para o aprofundamento da avaliação desse efeito em aves.
As aves do cruzamento TCTC apresentaram fígado e coração mais pesados que as aves CTCT. Essas duas características são de grande importância para o sistema de produção, pois aves com coração e fígado maiores podem ser mais resistentes aos estresses fisiológicos e de natureza ambiental.
O fato de utilizar pais da linha TT com mães da linha CC promoveu um aumento de 3,42% no peso do coração e de 2,08% no peso do fígado. Ressalta-se que, nesse caso, utilizar machos da linhagem de corte foi o que promoveu aumento nos pesos dessas características, contrastando com o que foi observado para peso vivo.
Outra diferença importante entre os recíprocos está no peso da moela e no comprimento do intestino. Aves CTCT apresentaram moela 3,02% mais pesada e intestino 2,96% maior que animais TCTC. Esses resultados podem ser a explicação para as aves CTCT terem peso vivo superior às TCTC, pois moelas mais pesadas e intestinos maiores podem contribuir para melhor aproveitamento digestivo dos nutrientes, garantindo maiores ganhos de peso.
O cruzamento CTCT também apresentou maiores pesos de dorso, peito e cabeça, promovendo ganhos extras em carcaça (0,82%), dorso (1,16%), peito (1,77%), cabeça (1,57%) e gordura abdominal (7,50%). Além disso, Ledur et al. (2000LEDUR, M.C.; ZANELLA, E.L.; SCHMIDT, G.S. et al. Peso e características de carcaça em linhagens utilizadas no desenvolvimento de populações referência para detecção de QTLS em aves. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 2000, Campinas. Anais... Campinas: Fundação Apinco de Ciência e Tecnologia Avícolas, 2000. 1v. CD-ROM. ) também detectaram maior porcentagem de gordura abdominal na população F1 CT, com 1,92%, comparada com a população F1 TC, com 1,51% de gordura.
O peso ao nascimento foi maior em aves CTCT, ou seja, aquelas que tiveram origem no acasalamento de machos de postura com fêmeas de corte. Esse peso tem relação direta com o peso do ovo, e essa influência pode se estender durante todo o período de criação (Traldi, 2009TRALDI, A.B.; MENTEN, J.F.M.; RACANICCI, A.M.C. et al. Estudo dos fatores que influenciam o peso de pintos de um dia: idade da matriz e peso do ovo. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 2009, Porto Alegre. Anais... Porto Alegre: Fundação Apinco de Ciência e Tecnologia Avícolas, 2009. 1v. CD-ROM. ). Um dos fatores que influenciam o peso do ovo é o peso das matrizes, pois fêmeas menores tendem a produzir ovos menores e, consequentemente, pintos mais leves (Danalezi et al., 2004DANALEZI, J.A.; MENDES, A.A.; GARCIA, E.A. et al. Efeito da idade da matriz sobre o rendimento e qualidade da carne de frangos de corte. Ciênc. Tecnol. Aliment., v.24, p.685-690, 2004.). Isso pode explicar a diferença entre as populações F2 para peso ao nascimento, pois, como observado em Ledur et al. (2000LEDUR, M.C.; ZANELLA, E.L.; SCHMIDT, G.S. et al. Peso e características de carcaça em linhagens utilizadas no desenvolvimento de populações referência para detecção de QTLS em aves. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 2000, Campinas. Anais... Campinas: Fundação Apinco de Ciência e Tecnologia Avícolas, 2000. 1v. CD-ROM. ), as aves F1 CT foram mais pesadas que as TC. As aves F1 CT tiveram como progenitoras aves de corte da linhagem TT, as quais produziram ovos maiores que as fêmeas da linhagem CC.
Apesar da diferença em peso corporal, não foram observadas diferenças significativas para peso de cortes cárneos comercialmente importantes, como asas e pernas, bem como para ganho de peso, conversão alimentar e consumo de ração avaliados entre o 35o e o 41o dia de idade. É possível que o período de teste de desempenho de apenas sete dias não tenha sido suficiente para detectar diferenças entre os cruzamentos TCTC e CTCT para conversão e consumo de ração entre 35 e 41 dias. Também é preciso considerar que houve elevado coeficiente de variação das características consumo de ração e conversão alimentar, o que é resultado de uma variância residual elevada. Logo, para essas duas características, pode ter havido erro tipo II no teste F da análise de variância para o efeito de cruzamento.
Foi possível observar algumas importantes diferenças entre os coeficientes de herdabilidade (Tab. 2 e 3) em função do cruzamento recíproco estudado. Na população CTCT, foram observados maiores valores de h2 que na população TCTC para o peso vivo ao nascer, os pesos do fígado, cabeça e pés e o comprimento do intestino. Já a população TCTC teve maiores valores de herdabilidade para os pesos da moela, pulmões, pernas, peito e asas. Para as demais características, não houve diferenças expressivas, provavelmente devido aos elevados erros-padrão das estimativas, como, por exemplo, para peso de gordura abdominal, que teve erros-padrão maiores que 0,15. Erros-padrão elevados para essas estimativas já eram esperados, visto que a amostra estudada é pequena.
A população TCTC foi estudada para fins de mapeamento de QTL por Nones et al. (2006NONES, K.; LEDUR, M.C.; RUY, D.C. et al. Mapping QTLs on chicken chromosome 1 for performance and carcass traits in a broiler x layer cross. Anim. Genet., v.37, p.95-100, 2006.), ao longo do cromossomo 1. Para as sete características que apresentaram herdabilidade maior que 0,40 (Tab. 3), esses autores mapearam nove QTLs. Para as seis características que apresentaram herdabilidade entre 0,20 e 0,40, foram mapeados 12 QTLs e para as seis características que apresentaram herdabilidade menor que 0,20, mapearam-se sete QTLs. Logo, parece não haver grande relação entre a magnitude dos coeficientes de herdabilidade aqui encontrados e o número de QTLs mapeados no estudo de Nones et al. (2006). Ressalta-se, porém, que muitos dos QTLs mapeados para características de moderada herdabilidade (PV35, PV42, pesos de pernas, carcaça e pés, e GP35-41) estão em posições bastante próximas, podendo ser QTLs pleiotrópicos, e, nesse caso, o número de QTLs seria menor do que os 12 relatados por Nones et al. (2006). Ainda, muitos dos altos coeficientes de herdabilidade do presente estudo também possuem altos erros-padrão, restando dúvidas quanto à acurácia desses altos valores de herdabilidade aqui encontrados.
Melhorar características de baixa herdabilidade, como o peso do coração, das asas, das pernas, do dorso e da conversão alimentar da população CTCT, ou os pesos do fígado, do coração, da cabeça e do dorso, a conversão alimentar e o comprimento do intestino da população TCTC, é difícil, pois apenas uma pequena parte da variação dessas características é devido às diferenças genéticas aditivas entre as aves. Assim, a seleção dos melhores animais não é uma garantia de que haverá ganhos expressivos nas gerações seguintes. Logo, para essas características, quanto mais informações mais preciso será o processo de seleção, e é nesse contexto que a seleção assistida por marcadores moleculares e a identificação das regiões cromossômicas responsáveis pela expressão dessas características pode contribuir.
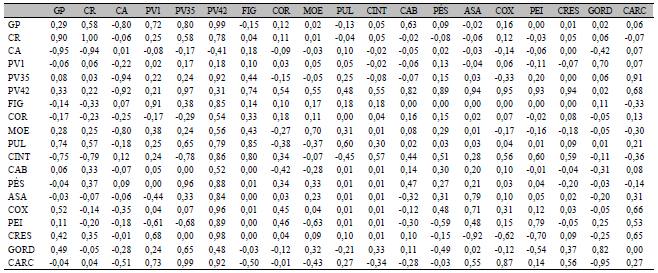
Das correlações genéticas calculadas com a população CTCT, 67,25% foram consideradas altas, 11,70% médias e 21,05% baixas, enquanto para as correlações fenotípicas, 61,40% foram altas, 19,30% moderadas e 19,30% baixas (Tab. 4). Por outro lado, a população TCTC apresentou 34,50% das correlações genéticas altas, 26,32% moderadas e 39,18% baixas, enquanto as correlações fenotípicas foram 18,71% altas, 10,53% moderadas e 70,76% baixas (Tab. 5). Existem várias razões para as diferenças entre as populações TCTC e CTCT quanto às estimativas de correlações. Uma é a alta variabilidade dentro dessas populações, pois ambas são populações F2 experimentais. Contudo, essas diferenças também podem ser devido a efeitos recíprocos, como pode ser observado em Tuiskula-Haavisto e Vilkki (2007). Causas para efeitos recíprocos podem ter origem em efeitos citoplasmáticos, em fatores genéticos ligados ao sexo, no efeito materno ou no efeito imprinting, embora este último ainda não tenha sido comprovado em aves, apesar de alguns estudos de mapeamento de QTL terem revelado regiões em cromossomos não sexuais com efeito diferenciado em função da origem do alelo (paterna ou materna), como pode ser observado na revisão de Tuiskula-Haavisto e Vilkki (2007).
As correlações genéticas podem ser úteis na avaliação da hipótese de pleiotropia versus ligação gênica, apesar de não ser um teste para esse fim. Se as correlações genéticas forem altas, tem-se um indicativo de QTL pleiotrópico, enquanto baixas correlações genéticas seriam um forte indício de QTLs ligados. No estudo de Nones et al. (2006NONES, K.; LEDUR, M.C.; RUY, D.C. et al. Mapping QTLs on chicken chromosome 1 for performance and carcass traits in a broiler x layer cross. Anim. Genet., v.37, p.95-100, 2006.), foram mapeados QTLs para várias das características aqui estudadas, na população TCTC. Muitos dos QTLs mapeados para essas características foram identificados em regiões próximas, no cromossomo 1, levantando a suspeita de QTLs pleiotrópicos. Por exemplo, na posição 332 cM esses autores encontraram QTL para PV35, PV42, pesos da carcaça, moela e pés.
Pode-se observar que existem altas correlações genéticas entre algumas características desse grupo, como entre PV42 e peso da carcaça (0,92) e PV35 e peso da carcaça (0,99) (Tab. 5), mas, em alguns casos, ela é praticamente nula. Assim, é difícil fazer uma avaliação mais objetiva sobre a existência de QTL pleiotrópico ou de QTLs ligados nessa população TCTC partindo dos valores de correlações aqui encontradas. Ressalta-se ainda que Nones et al. (2006NONES, K.; LEDUR, M.C.; RUY, D.C. et al. Mapping QTLs on chicken chromosome 1 for performance and carcass traits in a broiler x layer cross. Anim. Genet., v.37, p.95-100, 2006.) avaliaram apenas uma amostra da população TCTC aqui estudada, pois muitas aves não foram genotipadas.
CONCLUSÃO
A população F2 originada do cruzamento de machos de linhagem de postura e fêmeas de linhagem de corte (CTCT) originou animais mais pesados, com melhor conversão alimentar e, em média, maiores pesos de órgãos do sistema do trato gastrointestinal, porém o peso da carcaça eviscerada, do fígado e do coração da população TCTC (machos da linhagem de corte e fêmeas da linhagem de postura) foi maior em todas as idades. As variações de magnitude das herdabilidades e as correlações tanto genéticas quanto fenotípicas na população TCTC foram, em sua maioria, mais baixas que na população CTCT. Diante do exposto, pode-se concluir que há maior facilidade de detecção de QTLs na população CTCT.
REFERÊNCIAS
- ABASHT, B.; DEKKERS J.C.M.; LAMONT S.J. Review of quantitative trait loci identified in the chicken. Poult. Sci., v.85, p.2079-2096, 2006.
- DANALEZI, J.A.; MENDES, A.A.; GARCIA, E.A. et al. Efeito da idade da matriz sobre o rendimento e qualidade da carne de frangos de corte. Ciênc. Tecnol. Aliment., v.24, p.685-690, 2004.
- GAYA, L.G.; MOURÃO, G.B.; REZENDE, F.M. et al. Genetic trends of abdominal fat content in a male broiler chicken line. Genet. Mol. Res., v.4, p.760-764, 2005.
- GROENEVELD, E.; KOVAC, M.; MIELENZ, N. VCE user's guide and reference manual. Version 6.0. Urbana, IL: University of Illinois/ Department of Animal Science, 2008.
- HITCHINS, M.P.; MOORE, G.E. Genomic imprinting in fetal growth and development. Expert. Rev. Mol. Med., v.9, p.1-19, 2002.
- HOCKING, P. Review on QTL mapping in chickens. Poult. Sci. J. , v.61, p.215-226, 2005.
- KNOTT, S.A.; HALEY, C.S. A simple regression method for mapping quantitative trait loci in line crosses using flanking markers. Heredity, v.69, p.315-324, 1992.
- LEDUR, M.C.; ZANELLA, E.L.; SCHMIDT, G.S. et al. Peso e características de carcaça em linhagens utilizadas no desenvolvimento de populações referência para detecção de QTLS em aves. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 2000, Campinas. Anais... Campinas: Fundação Apinco de Ciência e Tecnologia Avícolas, 2000. 1v. CD-ROM.
- NONES, K.; LEDUR, M.C.; RUY, D.C. et al. Mapping QTLs on chicken chromosome 1 for performance and carcass traits in a broiler x layer cross. Anim. Genet., v.37, p.95-100, 2006.
- TRALDI, A.B.; MENTEN, J.F.M.; RACANICCI, A.M.C. et al. Estudo dos fatores que influenciam o peso de pintos de um dia: idade da matriz e peso do ovo. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 2009, Porto Alegre. Anais... Porto Alegre: Fundação Apinco de Ciência e Tecnologia Avícolas, 2009. 1v. CD-ROM.
- TUISKULA-HAAVISTO, M.; VILKKI, J. Parent-of-origin specific QTL - a possibility towards understanding reciprocal effects in chicken and the origin of imprinting. Cytog. Gen. Res., v.117, p.305-312, 2007.
Datas de Publicação
-
Publicação nesta coleção
May-Jun 2016
Histórico
-
Recebido
17 Maio 2015 -
Aceito
15 Nov 2015