ABSTRACT
This work aimed to describe the origin, distribution, and ramifications of the ischiadicus nerve in the giant anteater and to provide anatomical data which could explain not only the evolutionary aspects but also provide important information for other related works. For the present study, four specimens were used, prepared by perfusion of 10% formaldehyde solution via the femoral artery, for conservation and dissection. The origin of the right and left ischiadicus nerves in the giant anteater from the ventral ramification of the third lumbar (L3) and the first (S1), second (S2), and third (S3) sacral spinal nerves. These nerves were symmetrical in all animals studied. The distribution and ramification occurred to the superficial, middle, and deep gluteal, gemelli, piriform, quadratus femoris, tensor fasciae latae, caudal crural abductor, cranial and caudal parts of the biceps femoris, adductor, semitendinous, and cranial and caudal parts of the semimembranous muscles. Based on the origins of the ischiadicus nerves, there is a caudal migration in the nerve location in animals in a more recent position on the evolutionary scale due to reconfiguration of the lumbosacral plexus, resulting from the increase in a number of lumbar vertebrae. There is no complete homology of the muscle innervation.
Keywords:
myrmecophagidae; pilosa; evolution; innervations
RESUMO
Objetivou-se descrever as origens, distribuições e ramificações dos nervos isquiáticos no tamanduá-bandeira, disponibilizando, assim, dados anatômicos que possam não só elucidar os aspectos evolutivos como também fornecer informações importantes para áreas afins. Foram utilizados quatro espécimes preparados por meio da perfusão de formaldeído 10% via artéria femoral, para conservação e dissecação. As origens dos nervos isquiáticos direito e esquerdo no tamanduá-bandeira foram provenientes dos ramos ventrais dos nervos espinhais lombares três e sacrais um, dois e três, sendo simétricos em todos os animais estudados. As distribuições e ramificações ocorreram nos músculos glúteos superficial, médio e profundo; gêmeo; piriforme; quadrado femoral; tensor da fáscia lata; abdutor crural caudal; bíceps femoral parte cranial; bíceps femoral parte caudal; adutor; semitendíneo; semimembranáceo parte cranial e semimembranáceo parte caudal. Notou-se que houve uma migração caudal na localização deste nervo nos animais mais recentes na escala evolutiva, devido a uma reconfiguração do plexo lombossacral decorrente do aumento no número de vértebras lombares, não havendo uma homologia total quanto à inervação dos músculos.
Palavras-chave:
Myrmecophagidae; ordem pilosa; evolução; inervação
INTRODUCTION
The giant anteater, Myrmecophaga tridactyla, is a member of the placental superordem Xenarthra, representing the order Pilosa and belonging to the family Mymercophagidae (Wilson and Reeder, 2005WILSON, D.E.; REEDER´S, D.A. Mammal species of the world: a taxonomic and geographic reference. Baltimore: Johns Hopkins University Press, 2005. v.48.). The Xenarthra are restricted to the New World in a determined geographic area and are morphologically isolated (Engelmann, 1985ENGELMANN, G.F. The phylogeny of the xenarthra. In: MONTGOMERY, G.G. (Ed.) The evolution and ecology of armadillo, sloths, and vermilinguas. [s.l.]: Institution Press, 1985. p.51-64.) from the rest of the placental mammals, which likely occurred during the Cretaceous period, as long as 106 million years ago (Delsuc et al., 2001DELSUC, F.; CATZEFLIS, F.M.; STANHOPE, M.J.; DOUZERY, E.J.P. The evolution of armadillos, anteaters, and sloths depicted by nuclear and mitochondrial phylogenies: implications for the status of the enigmatic fossil Eurotamandua. Proc. Royal Soc. Biol. Sci., v.268, p.1605-1615, 2001.). A series of derived characters were developed throughout the Xenarthra evolution due to this isolation. Amongst the morphologic singularities, one can quote as example the number of cervical vertebrae that varies from six to nine, depending on the species, while most mammals have seven cervical vertebrae; the urinary tract and the female genitalia and/or the male testicles share the same duct (Nowak, 1999NOWAK, R.M. Walker’s mammals of the world. v.1. Baltimore: Johns Hopkins University Press, 1999.).
According to Carvalho-Barros et al. (2003), the evolutionary aspects of the posture and locomotion are understood through the study of the neural plexus, with the lumbosacral plexus being of great importance as it is the representative of the origin of the pelvic member nerves. It is extremely important to know the origin, distribution, and ramification of the ischiadicus nerve because it is considered vulnerable to several lesions, especially a few centimeters caudal to the femur, between the biceps femoris and semimembranous muscles (Dyce et al., 2004DYCE, K.M.; SACK, W.O.; WENSING, C.J.G. Tratado de anatomia veterinária. Guanabara Koogan, Rio de Janeiro, 2004. 856p.). Symptoms from ischiadicus nerve lesions include insensitivity and motor dysfunction on the gluteus area, thigh and leg of the affected limb (Guimarães et al., 2005GUIMARÃES, G.C.; MACHADO, M.R.F.; SANTOS, A.L.Q. et al. Origin and distribution of the sciatic nerve in the domestic cat (Felis catus domesticus Linnaeus, 1758). Biosci. J., v.21, p.189-195, 2005.).
The objective of this study was to describe the origin, distribution and ramifications of the ischiadicus nerve of the giant anteater in order to make available anatomy data that can elucidate not only the evolutionary aspects but also provide important information for other related works.
MATERIAL AND METHODS
Four adult male giant anteater specimens, with a body mass of approximately 40kg were used for this study. The specimens were fixed with an injection of a liquid solution of 10% formaldehyde through femoral artery perfusion and conserved in the same solution. All the specimen preparations were done according to routine macroscopic dissection procedures (Rodrigues, 2005RODRIGUES, H. Técnicas anatômicas. Vitória: Arte Visual, 2005.). A longitudinal incision was made along the ventral midline the xiphoid cartilage of the xiphoid process of the sternum to the caudal border of the pelvic symphysis. Two other transversal incisions were made in parallel with the cranial border of each antimere to the dorsal midline. In order to visualize the origin, distribution, and ramifications of the ventral branches of the lumbar and sacral spinal nerves from both antimeres, the pelvic symphysis was disarticulated through a longitudinal incision, and the abdominal and pelvic organs and adjacent fat tissue were removed. After the right and left ventral branches of the ischiadicus nerves were identified, the skin and subcutaneous fascia from the median and lateral gluteal regions of the thighs were folded so that the distribution and ramification of the nerve could be analyzed.
To confirm the number of lumbar and sacral vertebrae, each animal underwent radiographic examination in ventral-dorsal and latero-lateral projections. The examinations were performed in the radiology department of the Veterinary Hospital of the Faculty of Veterinary Medicine and Animal Science, UFU. Because of the lack of works in this area, studies on the origin and distribution of the ischiadicus nerves of two members of the Xenarthra superorder, such as M. tridactyla (Cruz et al., 2013CRUZ, V.S.; CARDOSO, J.R.; ARAÚJO, L.B.M. et al. Aspectos Anatômicos dos Nervos da Coxa de Tamanduá-bandeira (Myrmecophaga tridactyla, Linnaeus, 1758). Biosci. J., v.29, p.1275-1283, 2013. and 2014) and Tamandua tetradactyla (Cardoso et al., 2013CARDOSO, J.R.; SOUZA, P.R.; CRUZ, V.S. et al. Estudo anatômico do plexo lombossacral de Tamandua tetradactyla. Arq. Bras. Med. Vet. Zootec., v.65, p.1720-1728, 2013.) were analyzed. In addition, another study performed by our research group involving the bone anatomy of the pelvic girdle, the thigh, and the leg of M tridactyla (Ribeiro et al., 2013RIBEIRO, P.R.Q.; SANTOS, A.L.Q.; SOUZA, R.R. et al. Anatomia óssea do cíngulo pélvico, da coxa e da perna do tamanduá bandeira Myrmecophaga tridactyla (Myrmecophagidae: pilosa). Biotemas, v.26, p.153-160, 2013.) was also considered.
The anatomic nomenclature used to designate the identified structures was in accordance with the International Committee on Veterinary Gross Anatomical Nomenclature (Nomina..., 2017). The study was approved by the Animal Use Committee of the Federal University of Uberlandia, protocol nº 039/11.
RESULTS
The presence of three lumbar and four sacral vertebrae were recognized on the four specimens of M. tridactyla. The right and left ischiadicus nerves originated from the ventral branch of the third lumbar (L3) spinal nerve and the ventral branch of the first (S1), second (S2), and third (S3) sacral spinal nerves, demonstrating symmetry in all of the studied animals (Figure 1A and B).
Ventral macrophotography of the origin of the ischiadicus nerve (I) on the left antimere in M. tridactyla. Ventral branch of the third lumbar (L3) spinal nerve; Ventral branch of the first sacral (S1) spinal nerve; Ventral branch of the second sacral (S2) spinal nerve; Ventral branch of the third sacral (S3) spinal nerve. B. Ventral macrophotography of the left antimere showing the course of the ischiadicus nerve (I) through the major ischiatic foramen, which is bounded by the internal obturator muscle (IO), deep gluteal muscle (DG), sacrotuberous ligament (ST), and the major ischiatic incisures (MI). Abbreviations: obturator nerve (Ob) and psoas minor muscle tendon (PM).
The ischiadicus nerve consisted of an ischiadicus plexus formed by a nerve trunk resulting from the union of the ventral branches of the spinal nerves above. Subsequent to the formation of the ischiadicus nerve, it emerged from the pelvic cavity through the major ischiadicus foramen, which is circumscribed by the internus obturator and deep gluteal muscles, the broad sacrotuberal ligament, and the greater ischiadicus notch (Figure 1A and B). A peculiar distribution of the ischiadicus nerve was noted because in a lateral view, it was divided at the level of the greater trochanter of the femur into two branches with different thickness, namely the thicker superficial branch of the ischiadicus nerve and the thinner deep branch of the ischiadicus nerve (Figure 2A and B).
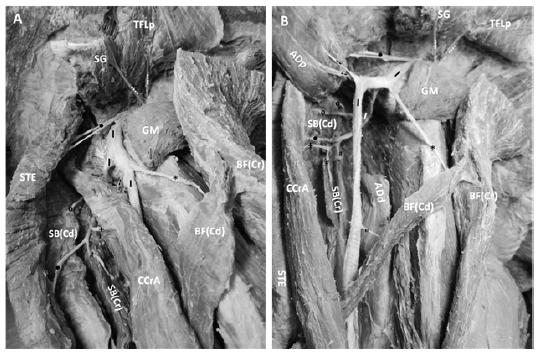
Macrophotography of the distribution and ramification of the ischiadicus nerve (I) to the muscles of the right lateral face of the pelvis and thigh in M. tridactyla. (⁞) ischiadicus nerve branches to the superficial gluteal muscle (SG) and proximal part of the tensor fasciae latea muscle (TFLp); (•) ischiadicus nerve branches to the semitendinous muscle (STE); (*) ischiadicus nerve branches to the cranial part of the biceps femoris muscle - BF(Cr); (") ischiadicus nerve branches to the caudal part of the biceps femoris muscle- BF(Cd); (⁰) ischiadicus nerve branches to the caudal part of the semimembranous muscle - SB(Cd); (#) ischiadicus nerve branches to the cranial part of the semimembranous muscle - SB(Cr); (-) ischiadicus nerve branches to the proximal part of the adductor muscle (ADP); (+) ischiadicus nerve branches to the caudal crural abductor muscle (CCrA). B. Macrophotography of the ischiadicus nerve (I) after section of the adductor (AD) and caudal crural abductor (CCrA) muscles, showing the subdivision of the ischiadicus nerve into a thicker superficial branch, and a thinner deep branch that extends at mediocaudal position, by passing the adductor (AD) and caudal crural abductor (CCrA) muscles as demonstrated in panel A. Abbreviation: ADd- distal part of the adductor muscle.
The superficial branch of the ischiadicus nerve extended on the lateral face of the thigh up to the level of the distal third, where it gave off branches to the cranial and caudal parts of the biceps femoris, semitendinous, caudal crural abductor, and adductor muscles. Next, it divided into the tibial, fibularis communis, and lateral cutaneous of the sura nerves (Figure 3). The deep branch of the ischiadicus nerve extended to a mediocaudal position, bypassing the adductor and caudal crural abductor muscles and turning laterally to be distributed to the caudal crural abductor and the cranial and caudal parts of the semimembranous and semitendinous muscles (Figure 2A and B).
Macrophotography of the distal third of the lateral face of the thigh on the right antimere in giant anteater. Distribution and ramifications of the ischiadicus nerve (I) to the following muscles: (*) Cranial part of the biceps femoris - BF(Cr); (") Caudal part of the biceps femoris- BF(Cd); (•) Semitendinous- STE; (+) Caudal crural abductor. Abbreviations: Fb -fibularis communis nerve; Tb - tibial nerve; S - Lateral cutaneous sural nerve.
Throughout its course, the ischiadicus nerve supplied branches to the superficial, middle, and deep gluteal, gemelli, piriform, quadratus femoris, tensor fasciae latae, caudal crural abductor, cranial and caudal parts of the biceps femoris, adductor, semitendinous, and cranial and caudal parts of the semimembranous muscles (Figure 4 and Figure 2A and B).
Macrophotography of the lateral face of the pelvis and thigh on the right antimere in giant anteater. Distribution and ramifications of the ischiadicus nerve (I) to the following muscles: (*) distal and proximal part of the tensor fasciae latea muscle (TFLd); (⁰) middle gluteal (MG); (⁞) superficial gluteal (SG); (•) semitendinous (STE); (+) part cranial of the biceps femoris muscle -BF(Cr); (#) Adductor (AD). Abbreviations: DG - deep gluteal muscle; PF - piriform muscle; GE - gemelli muscle; QF - quadradus femoris.
On the right antimere, 50% of the animals had the ischiadicus nerve supplying two branches to the superficial gluteal muscle and in the remaining 50%, three branches, while on the left antimere the same muscle received four branches in 25% of the animals and three branches in 75%. On both of the antimeres, the middle gluteal muscle received two branches in 75% and one branch in 25% of the specimens. The distribution and ramification of the ischiadicus nerve to the deep gluteal muscle were of two branches in 75% of the specimens on the right antimere, where as all (100%) of the animals had only one branch on the left. On the gemelli muscle, 67% of the animals had one branch on the right antimere and 100% had one branch on the left.
The piriform muscle received two branches of the ischiadicus nerve in 75% of the animals on the right antimere and only one branch (50% of cases) on the left antimere. The ischiadicus nerve provided one branch to the gemelli muscle (75% of cases) on the right antimere, while all (100%) of the specimens had only one branch on the left antimere. The distribution and ramification to the quadratus femoris muscle were of two branches (50% of cases) on the right antimere and only one branch (100% of cases) on the left antimere.
All of the studied specimens had two branches to the tensor fasciae latea muscle on the right antimere and one branch in 75% of cases on the left. The distribution and ramification to the adductor muscle were four branches (75% of cases) on the right antimere and two branches (50% of cases) on the left. The ischiadicus nerve provided four branches (75% of cases) to the caudal crural abductor muscle on the right antimere and on the left two branches in 25% and four in 75% of the animals.
The presence of four (75% of cases) and three (25% of cases) branches to the cranial part of the biceps femoris muscle on the right antimere were observed, while three (75% of cases) and two (25% of cases) branches were found on the left. The caudal part of the biceps femoris muscle received three (75% of cases) and two (25% of cases) branches on the right antimere and three (50% of cases) and two (50% of cases) on the left.
The semitendinous muscle had from three (25% of cases) to four (75% of cases) branches on the right antimere while on the left it had two branches in 25% and three in 75% of the animals. On the right antimere, the cranial part of the semimembranous muscle received three branches in 25% and four in 75% of the specimens, and on the left antimere two branches in 75% and three in 25% of cases. The caudal part of the semimembranous muscle had seven branches in 75% and eight in 25% of the specimenson the right antimere while on the left it had nine branches in 75% and ten in 25% of the studied animals (Table 1).
Muscle ramifications of the ischiadicus nerve on the right and left antimeres in M. tridactyla
DISCUSSION
The giant anteater (M. tridactyla) is reported to have two or three lumbar vertebrae. There may be a correlation between the number of thoracic and lumbar vertebrae, such as for 15 thoracic vertebrae present there are three lumbar vertebrae, and 16 thoracic vertebrae are commonly correlated with two lumbar vertebrae (Flower, 1885FLOWER, W.H. An introduction to the osteology of the mammalian. London: Macmillan, 1885.). This author also reported that there might be three to five sacral vertebrae in this animal species; however, in the present study, the four specimens of M. tridactyla had three lumbar and four sacral vertebrae. The cranial gluteal nerve was present in six specimens of giant anteater (Cruz et al., 2013CRUZ, V.S.; CARDOSO, J.R.; ARAÚJO, L.B.M. et al. Aspectos Anatômicos dos Nervos da Coxa de Tamanduá-bandeira (Myrmecophaga tridactyla, Linnaeus, 1758). Biosci. J., v.29, p.1275-1283, 2013.). However, in a more recent study, six specimens of this animal had the cranial and caudal gluteal nerves in the constitution of the lumbosacral plexus (Cruz et al., 2014).
Thus, there are several contradictory findings and variations in the number of lumbar and sacral vertebrae (Flower, 1885FLOWER, W.H. An introduction to the osteology of the mammalian. London: Macmillan, 1885.; Cruz et al., 2013CRUZ, V.S.; CARDOSO, J.R.; ARAÚJO, L.B.M. et al. Aspectos Anatômicos dos Nervos da Coxa de Tamanduá-bandeira (Myrmecophaga tridactyla, Linnaeus, 1758). Biosci. J., v.29, p.1275-1283, 2013.; Cruz et al., 2014). In our study we considered that the M. tridactyla specimens had no cranial and caudal gluteal nerves, but a nerve trunk consisting of an ischiadicus plexus, resulting from the union of the ventral branches of the third (L3) lumbar and first (S1), second (S2) and third (S3) sacral spinal nerves. Therefore, in this representative member of the Pilosa, the ischiadicus nerve innervates the muscles that are commonly innervated by the cranial and caudal gluteal nerves in animals with four or more lumbar vertebrae.
According to the classical literature on the anatomy of domestic animals, the origin of the ischiadicus nerve in ruminants was from the ventral branches of the fifth (L5) and sixth (L6) lumbar and the first (S1) and second (S2) sacral spinal nerves (Bruni and Zimmerl, 1977BRUNI, A.C.; ZIMMERL, U. Anatomia degli animali domestic - nervi spinali. [s.l.]: Dottor Francesco Vallardi, 1977. v.2, p.535-564.). In zebu-crossed bovine fetuses, the ischiadicus nerve had its origin from the ventral branches of the sixth (L6) lumbar and the first (S1) and second (S2) spinal sacral nerves (Ferraz et al., 2006FERRAZ, R.H.S.; LOPES, G.R.; MELO, A.P.F.; PRADA, I.L.S. Estudo anatômico da porção intrapélvica do nervo isquiático em fetos de bovinos azebuados. Braz. J. Vet. Res. Anim. Sci., v.43, p.302-308, 2006.). In zebu-crossed cattle (Ferraz et al., 2006), the ischiadicus nerve was also originated from the ventral branches of the sixth (L6) lumbar and first (S1) and second (S2) sacral spinal nerves, having in some cases the contribution from the third (S3) sacral spinal nerve. In dogs (Evans and De Lahunta, 2001EVANS, H.E.; DELAHUNTA, A. Guia para a dissecação do cão. Guanabara Koogan, Rio de Janeiro, 2001.) and domestic cats (Guimarães et al., 2005GUIMARÃES, G.C.; MACHADO, M.R.F.; SANTOS, A.L.Q. et al. Origin and distribution of the sciatic nerve in the domestic cat (Felis catus domesticus Linnaeus, 1758). Biosci. J., v.21, p.189-195, 2005.) the origin of the ischiadicus nerve was from the ventral branches of the sixth (L6) and seventh (L7) lumbar and the first (S1) and second (S2) sacral spinal nerves.
The origin of the ischiadicus nerve in gray brocket was derived from the ventral branches of the sixth (L6) lumbar and first (S1), second (S2), and third (S3) sacral spinal nerves (De Camargo et al., 2008). However, considering the same deer species, Martins et al. (2013MARTINS, T.M.M.; PEREIRA, K.F.; LIMA, F.C. et al. Origem e distribuição do nervo isquiático no Veado-catingueiro (Mazama gouazoubira). Braz. J. Vet. Res., v.33, p.273-278, 2013.) described the nerve origin predominantly from the ventral branches of the sixth (L6) lumbar spinal nerve and the first sacral spinal nerve (S1), having in some cases a contribution from the second pair of sacral (S2) spinal nerves.
In southern tamandua, the ischiadicus nerve originated from the ventral branches of the third (L3) lumbar and the first (S1), second (S2) and third (S3) sacral spinal nerves (Cardoso et al., 2013CARDOSO, J.R.; SOUZA, P.R.; CRUZ, V.S. et al. Estudo anatômico do plexo lombossacral de Tamandua tetradactyla. Arq. Bras. Med. Vet. Zootec., v.65, p.1720-1728, 2013.). This information about the origin of the ischiadius nerve in another member of the Pilosa confirms the morphological characteristics that recognize a phylogenetic homology between the two members of this order, Tamandua tetradactyla and M. tridactyla.
According to Cruz et al. (2013CRUZ, V.S.; CARDOSO, J.R.; ARAÚJO, L.B.M. et al. Aspectos Anatômicos dos Nervos da Coxa de Tamanduá-bandeira (Myrmecophaga tridactyla, Linnaeus, 1758). Biosci. J., v.29, p.1275-1283, 2013. and 2014), the origin of the ischiadicus nerve in giant anteater has been predominantly observed from the ventral branches of the third (L3) lumbar and the first (S1), second (S2), and third (S3) sacral spinal nerves. However, some variations on the number of lumbar vertebrae can be observed, thus characterizing an origin derived from the ventral branches of the second (L2) lumbar and the first (S1), second (S2), third (S3), and fourth (S4) sacral spinal nerves. In our findings, the origins of the ischiadicus nerve on both antimeres in four M. tridactyla specimens were from the ventral branches of the third (L3) lumbar and the first (S1), second (S2), and third (S3) sacral spinal nerves. This origin configuration suggests a plesiomorphic morphological condition of this placental mammal when compared to other more recent wild and domestic mammals on the phylogenetic scale.
Considering the origin of the ischiadicus nerve in all placental mammals quoted in this study, it was noted a similarity in the formation of that nerve involving the last ventral branches of the lumbar spinal nerves and the first ventral branches of the sacral spinal nerves. This inference is consistent with reports by Engelmann (1985ENGELMANN, G.F. The phylogeny of the xenarthra. In: MONTGOMERY, G.G. (Ed.) The evolution and ecology of armadillo, sloths, and vermilinguas. [s.l.]: Institution Press, 1985. p.51-64.), who consider the members of this Xenarthra superorder as the most primitive placental mammals.
In giant anteater, the ischiadicus nerve provided branches to the superficial, middle and deep gluteal, gemelli, piriform, quadratus femoris, tensor fasciae latae, caudal crural abductor, cranial and caudal parts of the biceps femoris, adductor, semitendinous, and cranial and caudal parts of the semimembranous muscles. No branches to the internus obturator muscle were detected in the four specimens studied. Regarding the distribution of the ischiadicus nerve, branches to the semitendinous, semimembranous and biceps femoris were found in domestic dogs (Evans and De Lahunta, 2001EVANS, H.E.; DELAHUNTA, A. Guia para a dissecação do cão. Guanabara Koogan, Rio de Janeiro, 2001.), zebu-crossed bovine fetuses (Campos et al., 2003CAMPOS, D.B.; SILVA, F.O.C.; SEVERINO, R.S. et al. Origem e distribuição dos nervos isquiáticos em fetos de bovinos azebuados. Ars. Vet., v.19, p.219-223, 2003.), and domestic cats (Guimarães et al., 2005GUIMARÃES, G.C.; MACHADO, M.R.F.; SANTOS, A.L.Q. et al. Origin and distribution of the sciatic nerve in the domestic cat (Felis catus domesticus Linnaeus, 1758). Biosci. J., v.21, p.189-195, 2005.). Branches of the ischiadicus nerve to the superficial gluteal muscle were found in domestic dogs (Evans and De Lahunta, 2001).
Branches of the referred nerve were found going to the deep gluteal muscle in zebu-crossed bovine fetuses (Campos et al., 2003CAMPOS, D.B.; SILVA, F.O.C.; SEVERINO, R.S. et al. Origem e distribuição dos nervos isquiáticos em fetos de bovinos azebuados. Ars. Vet., v.19, p.219-223, 2003.), and domestic cats (Guimarães et al., 2005GUIMARÃES, G.C.; MACHADO, M.R.F.; SANTOS, A.L.Q. et al. Origin and distribution of the sciatic nerve in the domestic cat (Felis catus domesticus Linnaeus, 1758). Biosci. J., v.21, p.189-195, 2005.). Branches to the middle gluteal muscle were present in zebu-crossed bovine foetuses (Campos et al., 2003). The ramification of the ischiadicus nerve for gemelli and quadratus femoris muscles was also found in domestic dogs (Evans and De Lahunta, 2001EVANS, H.E.; DELAHUNTA, A. Guia para a dissecação do cão. Guanabara Koogan, Rio de Janeiro, 2001.), and domestic cats (Guimarães et al., 2005).
The ischiadicus nerve provided branches to the internus obturator muscle in ruminants (Godinho et al., 1987GODINHO, H.P.; CARDOSO, F.M.; NASCIMENTO, J.F. Anatomia dos ruminantes domésticos. Belo Horizonte: Departamento de Morfologia, Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais, 1987.), and domestic dogs (Evans and De Lahunta, 2001EVANS, H.E.; DELAHUNTA, A. Guia para a dissecação do cão. Guanabara Koogan, Rio de Janeiro, 2001.). According to Godinho et al. (1987), the fibers derived from the ischiadicus nerve are distributed to the tensor fasciae latae muscle in ruminants. Also, branches of this nerve to the adductor muscle were described in ruminants (Godinho et al., 1987). In domestic dogs, the caudal crural abductor muscle also received branches of the ischiadicus nerve (Ghoshal, 1986GHOSHAL, N.G. Nervos espinhais. In: GETTY, R. (1986). Anatomia dos animais domésticos. Rio de Janeiro: Guanabara Koogan, 1986. v.2, p.1595-1617.).
In wild mammals, the ischiadicus nerve has distributed to the gluteobiceps muscles in gray brocket (De Camargo et al., 2008). Nerve fibers were also observed going to the superficial gluteal muscle in wild boar (Iglesias et al., 2011IGLESIAS, L.P.; SILVA, F.O.C.; BRITO, T.R. Origem e distribuição do nervo isquiático em fetos de javalis (Sus scrofa scrofa). Biotemas, v.24, p.141-145, 2011.). The middle gluteal muscle in wild boar (Iglesias et al., 2011), and chinchilla (Martinez-Pereira and Rickes, 2011) has also received branches from the referred nerve. The innervation for the deep gluteal muscle was observed in gray brocket (De Camargo et al., 2008). The semitendinous muscle has also been innervated by the ischiadicus nerve in gray brocket (De Camargo et al., 2008), wild boar (Iglesias et al., 2011), and giant anteater (Cruz et al., 2013CRUZ, V.S.; CARDOSO, J.R.; ARAÚJO, L.B.M. et al. Aspectos Anatômicos dos Nervos da Coxa de Tamanduá-bandeira (Myrmecophaga tridactyla, Linnaeus, 1758). Biosci. J., v.29, p.1275-1283, 2013.).
The distribution and ramification of the ischiadicus nerve to the semimembranous muscle were observed in gray brocket (De Camargo et al., 2008), wild boar (Iglesias et al., 2011IGLESIAS, L.P.; SILVA, F.O.C.; BRITO, T.R. Origem e distribuição do nervo isquiático em fetos de javalis (Sus scrofa scrofa). Biotemas, v.24, p.141-145, 2011.), and giant anteater (Cruz et al., 2013CRUZ, V.S.; CARDOSO, J.R.; ARAÚJO, L.B.M. et al. Aspectos Anatômicos dos Nervos da Coxa de Tamanduá-bandeira (Myrmecophaga tridactyla, Linnaeus, 1758). Biosci. J., v.29, p.1275-1283, 2013.). In addition, the distribution of the nerve was evidenced to the biceps femoris muscle in gray brocket (De Camargo et al., 2008; Martins et al., 2013MARTINS, T.M.M.; PEREIRA, K.F.; LIMA, F.C. et al. Origem e distribuição do nervo isquiático no Veado-catingueiro (Mazama gouazoubira). Braz. J. Vet. Res., v.33, p.273-278, 2013.), and giant anteater (Cruz et al., 2013). The gemelli muscle was innervated by the ischiadicus nerve in gray brocket (De Camargo et al., 2008). Branches to the adductor muscle were present in wild boar (Iglesias et al., 2011).
Studies in giant anteater (Cruz et al., 2013CRUZ, V.S.; CARDOSO, J.R.; ARAÚJO, L.B.M. et al. Aspectos Anatômicos dos Nervos da Coxa de Tamanduá-bandeira (Myrmecophaga tridactyla, Linnaeus, 1758). Biosci. J., v.29, p.1275-1283, 2013.) reported branches of the ischiadicus nerve to the quadratus femoris muscle. This nerve has also distributed to the piriform and tensor fasciae latae muscles in wild boar (Iglesias et al., 2011IGLESIAS, L.P.; SILVA, F.O.C.; BRITO, T.R. Origem e distribuição do nervo isquiático em fetos de javalis (Sus scrofa scrofa). Biotemas, v.24, p.141-145, 2011.). According to Gadow (1882GADOW, H. Beitragezur myologie der hinteren extremitat der reptilien. Morphologisches, v.2, p.382-466, 1882.), different nerve plexuses in several reptile species can innervate the same muscle; therefore the neuromuscular homology theory can not be used as main guide in establishing muscle homology for this group. Therefore, as in Gadow’s (1882) findings, the neuromuscular homology hypothesis does not seem to be applicable to M. tridactyla. This inference agrees with Haines (1935HAINES, R.W. A consideration of the constancy of muscular nerve supply. J. Anatomy, v.70, p.33-55, 1935.), who discredits the idea that the nervous supply represents an infallible guide for muscle homologies.
CONCLUSION
The origins of the right and left ischiadicus nerve are from the ventral branches of the third (L3) lumbar and the first (S1), second (S2), and third (S3) sacral spinal nerves in a symmetric way in all M. tridactyla specimens studied. The distribution and ramification of the ischiadicus nerve occurred to the superficial, middle and deep gluteal, gemelli, piriform, quadratus femoris, tensor fasciae latae, caudal crural abductor, cranial and caudal parts of the biceps femoris, adductor, semitendinous, and cranial and caudal parts of the semimembranous muscles. Based on the origins of the ischiadicus nerves, there is a caudal migration in the nerve location in animals in a more recent position on the evolutionary scale due to a reconfiguration of the lumbosacral plexus, resulting from the increase in a number of lumbar vertebrae. In addition, there is no complete homology of the muscle innervation, which is maintained phylogenetically on the different groups of animals considered in this study.
REFERENCES
- BRUNI, A.C.; ZIMMERL, U. Anatomia degli animali domestic - nervi spinali. [s.l.]: Dottor Francesco Vallardi, 1977. v.2, p.535-564.
- CAMPOS, D.B.; SILVA, F.O.C.; SEVERINO, R.S. et al. Origem e distribuição dos nervos isquiáticos em fetos de bovinos azebuados. Ars. Vet., v.19, p.219-223, 2003.
- CARDOSO, J.R.; SOUZA, P.R.; CRUZ, V.S. et al. Estudo anatômico do plexo lombossacral de Tamandua tetradactyla. Arq. Bras. Med. Vet. Zootec., v.65, p.1720-1728, 2013.
- CARVALHO-BARROS, R.A.; PRADA, I.L.S.; SILVA, Z. et al. Lumbar plexus formation of the Cebus apella monkey. Braz. J. Vet. Res. Anim. Sci., v.40, p.373-381, 2003.
- CRUZ, V.S.; CARDOSO, J.R.; ARAÚJO, L.B.M. et al. Aspectos Anatômicos dos Nervos da Coxa de Tamanduá-bandeira (Myrmecophaga tridactyla, Linnaeus, 1758). Biosci. J., v.29, p.1275-1283, 2013.
- CRUZ, V.S.; CARDOSO, J.R.; ARAÚJO, L.B.M. et al. Aspectos Anatômicos do Plexo Lombossacral de Myrmecophaga tridactyla (Linnaeus, 1758). Biosci. J., v.30, p.235-244, 2014.
- DE CAMARGO, V.M.F.; GUERRA, R.R.; TRANQUILIM, M.V.; CAMPOS, D.B. Origem e distribuição dos nervos isquiáticos no Veado-catingueiro (Mazama gouazoubira). In: CONGRESSO BRASILEIRO DE MEDICINA VETERINÁRIA, 35., 2008, Gramado, RS. Anais... Gramado: [s.n.], 2008. (Resumo).
- DELSUC, F.; CATZEFLIS, F.M.; STANHOPE, M.J.; DOUZERY, E.J.P. The evolution of armadillos, anteaters, and sloths depicted by nuclear and mitochondrial phylogenies: implications for the status of the enigmatic fossil Eurotamandua. Proc. Royal Soc. Biol. Sci., v.268, p.1605-1615, 2001.
- DYCE, K.M.; SACK, W.O.; WENSING, C.J.G. Tratado de anatomia veterinária. Guanabara Koogan, Rio de Janeiro, 2004. 856p.
- ENGELMANN, G.F. The phylogeny of the xenarthra. In: MONTGOMERY, G.G. (Ed.) The evolution and ecology of armadillo, sloths, and vermilinguas. [s.l.]: Institution Press, 1985. p.51-64.
- EVANS, H.E.; DELAHUNTA, A. Guia para a dissecação do cão. Guanabara Koogan, Rio de Janeiro, 2001.
- FERRAZ, R.H.S.; LOPES, G.R.; MELO, A.P.F.; PRADA, I.L.S. Estudo anatômico da porção intrapélvica do nervo isquiático em fetos de bovinos azebuados. Braz. J. Vet. Res. Anim. Sci., v.43, p.302-308, 2006.
- FLOWER, W.H. An introduction to the osteology of the mammalian. London: Macmillan, 1885.
- GADOW, H. Beitragezur myologie der hinteren extremitat der reptilien. Morphologisches, v.2, p.382-466, 1882.
- GHOSHAL, N.G. Nervos espinhais. In: GETTY, R. (1986). Anatomia dos animais domésticos. Rio de Janeiro: Guanabara Koogan, 1986. v.2, p.1595-1617.
- GODINHO, H.P.; CARDOSO, F.M.; NASCIMENTO, J.F. Anatomia dos ruminantes domésticos. Belo Horizonte: Departamento de Morfologia, Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais, 1987.
- GUIMARÃES, G.C.; MACHADO, M.R.F.; SANTOS, A.L.Q. et al. Origin and distribution of the sciatic nerve in the domestic cat (Felis catus domesticus Linnaeus, 1758). Biosci. J., v.21, p.189-195, 2005.
- HAINES, R.W. A consideration of the constancy of muscular nerve supply. J. Anatomy, v.70, p.33-55, 1935.
- IGLESIAS, L.P.; SILVA, F.O.C.; BRITO, T.R. Origem e distribuição do nervo isquiático em fetos de javalis (Sus scrofa scrofa). Biotemas, v.24, p.141-145, 2011.
- NOMINA anatomica veterinária. 6.ed. Hannover, Rio de Janeiro: ICVGAN, 2017. 178p.
- MARTINS, T.M.M.; PEREIRA, K.F.; LIMA, F.C. et al. Origem e distribuição do nervo isquiático no Veado-catingueiro (Mazama gouazoubira). Braz. J. Vet. Res., v.33, p.273-278, 2013.
- MARTINEZ-PEREIRA, M.A.; RICKES, E.M. The spinal nerves that constitute the lumbosacral plexus and their distribution in the chinchilla. J. S. Afr. Vet. Assoc., v.82, p.150-154, 2011.
- NOWAK, R.M. Walker’s mammals of the world. v.1. Baltimore: Johns Hopkins University Press, 1999.
- RIBEIRO, P.R.Q.; SANTOS, A.L.Q.; SOUZA, R.R. et al. Anatomia óssea do cíngulo pélvico, da coxa e da perna do tamanduá bandeira Myrmecophaga tridactyla (Myrmecophagidae: pilosa). Biotemas, v.26, p.153-160, 2013.
- RODRIGUES, H. Técnicas anatômicas. Vitória: Arte Visual, 2005.
- WILSON, D.E.; REEDER´S, D.A. Mammal species of the world: a taxonomic and geographic reference. Baltimore: Johns Hopkins University Press, 2005. v.48.
Publication Dates
-
Publication in this collection
10 Oct 2019 -
Date of issue
Jul-Aug 2019
History
-
Received
05 Mar 2018 -
Accepted
21 Jan 2019