RESUMO
Desde tempos remotos, a pelagem é valorizada no agronegócio equestre. Em animais Campolina, objetivou-se avaliar a ocorrência de pelagens e a distribuição entre sexos nos estados do Brasil, assim como verificar a frequência desse fenótipo nos acasalamentos e associá-lo com medidas lineares e qualidade da marcha. Foram selecionados 44.437 registros definitivos e utilizaram-se testes de qui-quadrado para avaliar a distribuição de pelagens entre sexos, estados e décadas. Por análise descritiva, foi obtida frequência de acasalamento dos fenótipos e proporções resultantes nos filhos. Foram contabilizados os indivíduos acasalados e os mais usados por décadas de nascimento e de registro. Pelagens baia, alazã e castanha predominaram, com 20.422; 11.941 e 5.256 animais, respectivamente. O fenótipo baio representou 45,21% em Minas Gerais; 46,98% no Rio de Janeiro e 48,98% em São Paulo. Para machos e fêmeas constatou-se maior frequência de pelagens baia, alazã e castanha. Os acasalamentos mais frequentes foram alazã x baia, baia x baia e baia x castanha. Este é o primeiro artigo a avaliar a ocorrência das pelagens em equinos da raça Campolina, sendo baia a mais prevalente em ambos os sexos. A qualidade da marcha ao registro não está associada à pelagem. Esse fenótipo deve ser usado com cautela na seleção.
Palavras-chave:
Equus caballus; raças equinas brasileiras; distribuição de frequência
ABSTRACT
Since ancient times, coat colors are valued in the equestrian business. The objective of this study was to evaluate the occurrence of coat colors and distribution among sexes in Brazil and also verify the frequency of this phenotype in reproduction to associate it with linear measurements and functionality. 44,437 definitive records were selected, and chi-square tests were used to evaluate coat colors and distribution between sexes, states and decades. By descriptive analysis the phenotypes breeding frequency and proportions in the offspring were obtained. Breeding and most used individuals were accounted by decade of birth and registration. Dun, chestnut and bay coat colors predominated, with 20,422; 11,941 and 5,256 animals, respectively. Dun represented 45.21% in Minas Gerais; 46.98% in Rio de Janeiro and 48.98% in São Paulo. For males and females there was a higher frequency of dun, chestnut and bay. The most frequent breeding colors were chestnut x dun, dun x dun and dun x bay. This is the first article to evaluate the coat colors in the Campolina population, dun being the most prevalent coat color in both sexes. The gait quality at the register is not associated with the coat color. This phenotype should be used with caution in selection.
Keywords:
Equus caballus; brazilian horse breeds; distribution of frequency
INTRODUÇÃO
A pelagem encontra-se entre os atributos fenotípicos equinos de interesse para criadores e pesquisadores (Castle, 1954CASTLE, W.E. Coat color inheritance in horses and in other mammals. Genetic, v.39, p.30, 1954. ; Thiruvenkadan et al., 2008THIRUVENKADAN, A.K.; KANDASAMY, N.; PANNERSELVAM, S. Coat colour inheritance in horses. Livest. Sci., v.117, p.109-129, 2008. ). Essa característica externa dos animais resulta do conjunto de pigmentação, despigmentação e reações oxidativas na pele, pelos, crina e cauda (Castle, 1954; Rezende e Costa, 2012REZENDE, A.S.C.; COSTA, M.D. Pelagem dos equinos: nomenclatura e genética. 2.ed. Belo Horizonte: FEPMVZ, 2012. 111p.). Apesar de não ser parâmetro estrito de desempenho, no âmbito comercial existe maior valor agregado para pelagens pouco comuns (Hennessy et al., 2008HENNESSY, K.D.; QUINN, K.M.; MURPHY, J. Producer or purchaser: different expectations may lead to equine wastage and welfare concerns. J. Appl. Anim. Welfare Sci., v.11, p.232-235, 2008. ). Tal modelo valorativo ocorreu também em equinos da raça Quarto de Milha na região Nordeste do Brasil (Maia et al., 2013MAIA, F.D.R.B.; SILVA, R.A.; MARACAJÁ, P.B. et al. Caracterização dos tipos de pelagens dos filhos de reprodutores machos “PO” da raça Quarto de Milha em microrregião do sertão Paraibano. Agropecu. Cient. Semiárido, v.9, p.124-131, 2013. ; Bastos et al., 2017BASTOS, M.S.; REZENDE, M.P.G.; SOUZA, J.C. et al. Levantamento da pelagem e idades de reprodutores Quarto de Milha utilizados na vaquejada em microrregiões do Nordeste do Brasil. Sci. Agrar. Paranaenses, v.16, p.62-68, 2017.).
De origem brasileira, o propósito inicial na raça Campolina foi o de obter animais de porte elevado, resistentes, ágeis e com andamento cômodo, sendo admitidas várias pelagens, conforme Associação Brasileira de Criadores do Cavalo Campolina (Origem, 2018). Pesquisas sobre pelagens em equinos Campolina exploraram poucos aspectos da relação pelagem com atributos de conformação e funcionais. Fontes (1957FONTES, L.R. Origem e características do cavalo campolina. 1957. 60f. Tese (Professor Catedrático em Zootecnia) - Universidade Federal de Minas Gerais. Belo Horizonte, MG.) e Procópio et al. (2003PROCÓPIO, A.M.; BERGMANN, J.A.G.; COSTA, M.D. Formation and demographic structure of the Campolina horse breed. Arq. Bras. Med. Vet., v.55, p.361-365, 2003. ) citaram as mais frequentes, enquanto Berbari Neto (2005) averiguou a distribuição das pelagens e a associação com medidas lineares, apenas para machos registrados em definitivo. Lucena et al. (2015LUCENA, J.E.C.; VIANNA, S.A.B.; BERBARI NETO, F. et al. Estudo comparativo das proporções morfométricas entre garanhões e castrados da raça Campolina. Semin. Ciênc. Agrár., v.36, p.353-366, 2015. ) descreveram a influência da pelagem sobre variáveis morfométricas de garanhões e machos castrados da raça Campolina.
No entanto, estudos sobre distribuição demográfica de pelagens, ocorrência de particularidades da pelagem, distribuição entre sexos e frequência de acasalamentos por pelagens, além de associações das pelagens com medidas corporais e funcionalidade, são ausentes na raça. Assim, em equinos da raça Campolina, objetivou-se avaliar a distribuição das pelagens por sexo, estados da federação, por acasalamentos e produtos, além de associar as pelagens com as medidas lineares e a pontuação do andamento por ocasião do registro definitivo.
MATERIAL E MÉTODOS
Foram utilizados os dados zootécnicos de animais registrados entre 1900 e 2016, disponibilizados pelo Serviço de Registro Genealógico da ABCCCampolina, contendo nome do animal, ano de nascimento e sexo; tipo, número e ano de registro; pelagem, particularidades e estados de criação; nome, registro e pelagem dos pais, anotados em livros aberto e fechado de machos e fêmeas, totalizando 106.452 animais. Apesar de fundada em 1951, ABCCCampolina mantém dados históricos dos primórdios da raça. Esses dados são de extrema importância para a avaliação da formação e do crescimento da população Campolina nacional. Realizaram-se análises de consistência, de modo a eliminar dados digitados erroneamente e/ou discrepantes, com os softwares Microsoft Access 2007® (Microsoft Corporation, 2007) e Statistical Analysis System ® - SAS 9.4 (Statistical..., 2014). Assim, foram obtidos 44.437 (41,74%) de dados de ambos os sexos, sendo 8.265 de machos e 36.172 de fêmeas, inseridos nos livros definitivos abertos e fechados, com descrição de pelagem no ato do registro. Animais inscritos em livros provisórios, n=58.299 (54,77%), não foram considerados nas análises em razão de a pelagem não estar totalmente definida.
Não foram consideradas nas análises as descrições das variedades das pelagens, utilizando-se, portanto, as nomenclaturas: alazã, baia, castanha, lobuna, pampa, preta, rosilha e tordilha (Castle, 1954CASTLE, W.E. Coat color inheritance in horses and in other mammals. Genetic, v.39, p.30, 1954. ; Rezende e Costa, 2012REZENDE, A.S.C.; COSTA, M.D. Pelagem dos equinos: nomenclatura e genética. 2.ed. Belo Horizonte: FEPMVZ, 2012. 111p.). Por análises exploratórias descritivas e de dispersão, foram determinadas a frequência das pelagens de acordo com o sexo, o estado, os anos de nascimento e de registro, e de acordo com os acasalamentos. Nos resultados das distribuições de frequência, utilizou-se o teste do qui-quadrado.
Apurações do uso dos indivíduos por pelagem foram feitas, sendo aferidos ascendentes mais utilizados por décadas de nascimento e de registro. Foram realizadas análises de correspondência para determinar associações entre pelagens e estados brasileiros em que ocorreram todas as pelagens: Bahia, Espírito Santo, Goiás, Minas Gerais, Pernambuco, Rio de Janeiro e São Paulo.
Para verificar o efeito de pelagens sobre medidas lineares de alturas de cernelha (ACER), dorso (ADOR), garupa (AGAR) e costado (ACOS); de comprimentos de cabeça (CCAB), pescoço (CPES), dorso (CDOR), garupa (CGAR), espádua (CESP) e corporal (CCORP); larguras de cabeça (LCAB), peito (LPEI) e garupa (LGAR); e perímetros torácico (PTOR) e de canela (PCAN), com bases anatômicas conforme descrito em Lucena et al. (2015LUCENA, J.E.C.; VIANNA, S.A.B.; BERBARI NETO, F. et al. Estudo comparativo das proporções morfométricas entre garanhões e castrados da raça Campolina. Semin. Ciênc. Agrár., v.36, p.353-366, 2015. ) e sobre as pontuações de andamento determinadas por técnico credenciado da ABCCCampolina: comodidade (PCOM), estilo (PEST), regularidade (PREG), desenvolvimento (PDES) e dissociação (PDIS), realizaram-se análises univariadas e teste T de Student (P<0,05) para comparar médias. Para avaliação de andamento, utilizaram-se informações de 5.196 animais, sendo pontuações PCOM, de um a 60; PDES, de um a 30; PDIS, de um a 40; PEST, de um a 40; e PREG, de um a 30, conforme ABCCCampolina (Regulamento..., 2006 e 2017).
O modelo misto geral utilizado para as análises estatísticas de mensurações lineares e pontuações de andamento foi descrito matricialmente, como: , em que: é o vetor correspondente às observações das características avaliadas; é a matriz correspondente aos efeitos fixos: pelagem, categoria animal (machos e fêmeas), ano de nascimento e idade, sendo esta entre três e quatro anos, entre quatro e cinco anos e animais acima de cinco anos para medidas lineares, acrescidos do efeito do técnico para as pontuações de andamento; é o vetor do efeito aleatório, correspondente à propriedade, o qual seguiu distribuição normal, com média igual a zero; e são as matrizes de incidência ligando observações aos efeitos fixos e aleatórios, respectivamente; é o vetor de resíduos.
Para análises de agrupamento hierárquico com o método de Ward, usaram-se médias de quadrados mínimos no PROC CLUSTER SAS 9.4 (Statistical..., 2014). Para agrupar as diferentes pelagens, realizaram-se procedimentos com medidas lineares ou pontuações de andamento e ambas as medidas. As análises estatísticas e o teste de comparação de médias foram feitos com PROC MIXED SAS 9.4 (Statistical..., 2014). Os componentes de variância foram obtidos por meio de máxima verossimilhança restrita.
RESULTADOS E DISCUSSÃO
A raça Campolina distribuiu-se em 21 estados, predominando em Minas Gerais, com 27.686 nascidos com pelagem descrita de 1885 a 2013, resultado justificado por ser o estado de origem da raça. Machos e fêmeas com pelagem conhecida a se reproduzirem foram 2.709 e 14.219, respectivamente. Do garanhão mais usado resultaram 976 filhos, e da matriz mais representativa procederam 132 progênies.
Em fêmeas foram mais frequentes as pelagens baia e castanha (Tab. 1), diferindo (P<0,05) dos machos. Essa diferença entre sexos pode associar-se com o maior número de registro de fêmeas em relação aos machos, justificado por custos de registro. Procópio et al. (2003PROCÓPIO, A.M.; BERGMANN, J.A.G.; COSTA, M.D. Formation and demographic structure of the Campolina horse breed. Arq. Bras. Med. Vet., v.55, p.361-365, 2003. ) descreveram registros de machos nos livros definitivos aberto e fechado, respectivamente, de 0,4 e 4,6% da população versus 22,4 e 16,6% nas fêmeas Campolina. A predominância de equinos baios (Tab. 1) pode estar vinculada à preferência por pelagens escuras e uniformes, como disposto no primeiro regulamento (Regulamento, 1938). Após o advento da associação em 1951, o critério de registro para pelagem passou a ser preferivelmente por animais baios, ainda sendo aceitos todos os outros fenótipos, exceto pseudoalbino (Serviço..., 1951). Para Fontes (1957FONTES, L.R. Origem e características do cavalo campolina. 1957. 60f. Tese (Professor Catedrático em Zootecnia) - Universidade Federal de Minas Gerais. Belo Horizonte, MG.), a maior frequência da pelagem baia pode estar relacionada à seleção de ancestrais da raça. Ademais, o uso dos reprodutores Golias e a sua progênie Otelo, durante a década de 30, contribuíram para dispersão desse fenótipo nas tropas da época (Berbari Neto, 2005). Consoante a isso, o alto número de indivíduos baios pode indicar maior probabilidade de ocorrência desses genes resultantes de acasalamentos entre animais baios com baio, alazão (e suas variedades), castanho, preto e rosilho (Rezende e Costa, 2012REZENDE, A.S.C.; COSTA, M.D. Pelagem dos equinos: nomenclatura e genética. 2.ed. Belo Horizonte: FEPMVZ, 2012. 111p.). É relevante considerar acasalamentos mais frequentes na população, os quais poderão determinar a variação das pelagens, em virtude da interação dos genes.
Da população estudada, 31.852 (71,67% da população) equinos Campolina apresentaram particularidades especiais de pelagem. Das especiais, foram mais frequentes as seguintes características: estrelas, calçamentos, listras de burro e beta (Tab. 2), as quais podem ser valorizadas por criadores e agregar valor comercial ao animal. Em virtude de 71,67% dessa ocorrência, infere-se ampla distribuição de genes Markings (M) na raça Campolina. Os alelos recessivos (mm) codificam presença de sinais na cabeça e nos membros (Negro et al., 2017NEGRO, S.; IMSLAND, M.; VALERA, M. et al. Association analysis of KIT, MITF, and PAX3 variants with white markings in Spanish horses. Anim. Genet., v48, p.349-352, 2017.; Rezende e Costa, 2012REZENDE, A.S.C.; COSTA, M.D. Pelagem dos equinos: nomenclatura e genética. 2.ed. Belo Horizonte: FEPMVZ, 2012. 111p.). Esse achado demonstra que, desde o início da formação da raça Campolina, houve seleção para essas particularidades especiais.
As frequências moderadas de listra de burro, faixa crucial e zebruras (Tab. 2) poderiam ser relacionadas aos 46,59% de indivíduos de pelagem baia na raça Campolina (Tab. 1). Rieder (2009RIEDER, S. Molecular tests for coat colours in horses. J. Anim. Breed. Genet., v.126, p.415-424, 2009.) relata que a pelagem baia e suas amplas variedades estão sempre adjuntas à presença dessas particularidades. Para Bailey e Brooks (2013BAILEY, E.; BROOKS, S.A. Horse genetics. 2.ed. London: CABI., 2013. p.42-64.), particularidades características desse fenótipo são associadas às marcas primitivas da pelagem equina.
Pelagens baia, alazã e castanha mostraram aumento de 1885 a 2013. Em 1885 registrou-se nascimento de animal alazão. Por ano de registro, a primeira inscrição de equino baio ocorreu em 1900. A partir de 1966, houve acréscimo de registro de baios (n=123). Máximo registro de fenótipos baios, alazões e castanhos incidiu em 1990, respectivamente com 1.338, 735 e 408 equinos, sendo em 1967 a ocorrência superior a 100 baios. O ápice de nascimentos de baios e alazões incidiu em 1988, com 838 e 485 potros. Maior ocorrência de castanhos foi em 1990, com 256 indivíduos.
Observou-se que, no decorrer das décadas de nascimento, houve seleção para pelagem castanha (P<0,05). Na década de 1940, a frequência observada de animais castanhos foi superior à esperada, sendo 40 e 25,44, respectivamente; enquanto em 2000 o nascimento de animais pampa foi duas vezes superior à frequência esperada de 444,25.
De forma similar às décadas de nascimento, verificou-se que as pelagens baias e castanhas (Fig. 1) apresentaram frequência acima do esperado (P<0,05), indicando seleção para essas na década de 1950, com 176 registros de baios e 58 de castanhos. O mesmo fato ocorreu em 2010 com as pelagens pampa, lobuna e preta (Fig. 1), as quais foram superiores ao esperado (P<0,05), respectivamente de 298,73; 269,44 e 151,37. Houve mudança da frequência de pelagens nas décadas e notou-se aumento da ocorrência de pampa, lobuna e preta, em detrimento da baia, alazã e castanha, apesar de essas serem as mais frequentes nas décadas avaliadas.
O declínio de registros de 1990 para 2000 pode associar-se à instabilidade econômica do Brasil entre 1990 e 1992, segundo Berbari Neto (2005). A redução de registros de machos inscritos em livro definitivo fechado foi constatada na raça até 2005 (Berbari Neto, 2005). A falta de apoio governamental após esse período pode ter resultado em vários entraves na equideocultura mineira, inviabilizando a retomada integral do setor (Vieira et al., 2015VIEIRA, E.R.; REZENDE, A.S.C.; LANA, A.M.Q. et al. Characterization of equidae breeding in Minas Gerais state. Arq. Bras. Med. Vet., v.67, p.319-323, 2015.).
Entre os fatores importantes que contribuíram para a expansão de indivíduos pampa (Fig. 1), inclui-se a criação da Associação Brasileira de Criadores do Cavalo Pampa (ABCCPampa) em 1993, na qual animais de diferentes raças são registrados por disporem dessa pelagem, ainda que possuam registro em outras associações. O aumento de pampas na raça pode relacionar-se também à mudança de perfil dos novos usuários do cavalo, ao visarem a práticas de lazer. Logo, o crescimento de fenótipos comerciais exóticos, muitas vezes resultantes da inserção de pessoas com conhecimento equestre restrito, pode contribuir para a dispersão de características indesejáveis, as quais precisam ser estudadas e os critérios de seleção, adotados de forma cautelosa.
A década de 2010, apesar de incompleta, indica o menor número de registros verificado desde 2000, o qual pode vincular-se ainda, conforme Berbari Neto (2005), ao critério e rigor de seleção de pelagem nos garanhões e ao advento de biotécnicas reprodutivas, que permitiram aumento do número de proles geradas por reprodutor. Porém, mesmo com a redução de animais inscritos em definitivo, houve aumento das proporções de pelagens inicialmente menos frequentes, como a pampa. Esse acréscimo pode justificar-se por possível valorização econômica em algumas regiões, com base nas tendências de compra e venda de equinos (Hennessy et al., 2008HENNESSY, K.D.; QUINN, K.M.; MURPHY, J. Producer or purchaser: different expectations may lead to equine wastage and welfare concerns. J. Appl. Anim. Welfare Sci., v.11, p.232-235, 2008. ).
Com 524 descendentes nas décadas de 1990 e 2000, Desacato da Maravilha foi o reprodutor preferido e exemplar, amplamente utilizado em razão das boas características de conformação, contribuindo para a maior frequência de animais baios. O animal pampa Garboso da Glória contribuiu com 131 filhos registrados na década de 2000, tendo instigado criadores e proprietários a buscar fenótipos menos comuns. Na década de 2010, Gavião do Barulho foi o garanhão mais utilizado e contribuiu para aumentar o número de filhos pampa, remetendo ao crescimento restrito, porém contínuo, de indivíduos com essa característica. A expansão verificada no número de animais pampa na raça (Fig. 1) coincide com a criação da Associação Brasileira de Criadores do Cavalo Pampa em 1993. A partir de 2011, julgamentos específicos do Campolina Pampa ratificaram a disseminação e a maior ocorrência na população e valorização comercial desse fenótipo.
Minas Gerais detém o maior número de registros, 65.114 animais, pois a raça originou-se no estado, está desenvolvida nacionalmente e encontra-se em sexta colocação no ranking nacional equestre, segundo Lima e Cintra (2016LIMA, R.A.S.; CINTRA, A.G. Revisão: estudo do complexo do agronegócio do cavalo. Brasília: Ministério da agricultura pecuária e abastecimento, 2016. p.56. ). A presente pesquisa revelou aumento no número de indivíduos e demonstrou a expansão nos estados, pois se registraram animais em 23 desses. Maior efetivo do Campolina em Minas Gerais foi descrito em 2000 (Procópio et al., 2003PROCÓPIO, A.M.; BERGMANN, J.A.G.; COSTA, M.D. Formation and demographic structure of the Campolina horse breed. Arq. Bras. Med. Vet., v.55, p.361-365, 2003. ) e 2011 (Vieira et al., 2015VIEIRA, E.R.; REZENDE, A.S.C.; LANA, A.M.Q. et al. Characterization of equidae breeding in Minas Gerais state. Arq. Bras. Med. Vet., v.67, p.319-323, 2015.).
A pelagem baia representou 45,21, 46,98 e 48,97% do observado, em Minas Gerais, Rio de Janeiro e São Paulo, respectivamente (Tab. 3). A alta incidência desse fenótipo nesses estados indica serem regiões tradicionais na criação do Campolina e de maiores plantéis, conforme Lima e Cintra (2016LIMA, R.A.S.; CINTRA, A.G. Revisão: estudo do complexo do agronegócio do cavalo. Brasília: Ministério da agricultura pecuária e abastecimento, 2016. p.56. ). Essa preferência pode fundamentar-se em parâmetros de seleção embasados em regulamento da raça e/ou por maior quantidade de equinos baios serem julgados e consagrados campeões, sendo, portanto, mais usados na reprodução. Observou-se diferença significativa na distribuição das frequências de pelagens entre estados (Tab. 3). Na Bahia, o número de 54 equinos registrados com pelagem pampa foi inferior (P<0,05) aos 170 esperados. No Rio de Janeiro, as frequências observadas para baia, lobuna e pampa foram, 46,98, 5,20 e 6,40%, respectivamente. Porém, foram inferiores (P<0,05) ao esperado da distribuição das pelagens alazã e tordilha, com 25,32 e 1,69%, nessa ordem.
Na análise de correspondência, verificou-se que os criadores de Minas Gerais escolheram mais a pelagem castanha, se comparados aos demais (Fig. 2). A preferência por animais castanhos e baios por estado foi observada quando essa é realizada de acordo com a primeira dimensão, a qual corresponde a 55,16% da inércia total.
Em Pernambuco e Goiás, demonstrou-se predileção por pelagem preta (Fig. 2). A seleção dos equinos por pelagem é comum e pode relacionar-se às prioridades do criador e aos títulos conquistados, possibilitando a maior utilização de animal específico em consequência de ser campeão, segundo Hennessy et al. (2008HENNESSY, K.D.; QUINN, K.M.; MURPHY, J. Producer or purchaser: different expectations may lead to equine wastage and welfare concerns. J. Appl. Anim. Welfare Sci., v.11, p.232-235, 2008. ) e Maia et al. (2013MAIA, F.D.R.B.; SILVA, R.A.; MARACAJÁ, P.B. et al. Caracterização dos tipos de pelagens dos filhos de reprodutores machos “PO” da raça Quarto de Milha em microrregião do sertão Paraibano. Agropecu. Cient. Semiárido, v.9, p.124-131, 2013. ). Apesar de a seleção na raça ser fundamentada nos aspectos morfológicos e funcionais conforme disposto no padrão racial vigente (Regulamento..., 2017), pode-se inferir a existência de proprietários e/ou criadores que escolhem reprodutores por predileções pessoais de pelagem, visando à facilidade de comercialização e ao maior valor atribuído às progênies com pelagens consideradas tendências ou raras, como a preta. Os animais castanhos, pampas, pretos e tordilhos apresentam médias de medidas lineares inferiores (P<0,05) às demais pelagens (Tab. 4). Isso pode relacionar-se ao critério de escolha de equinos somente por esse fenótipo.
Em relação às pelagens mais baixas, as mais altas foram superiores em um centímetro para ACER, ADOR e AGAR (Tab. 4). Berbari Neto (2005) verificou médias superiores de ACER, ADOR e AGAR em garanhões Campolina alazões, lobunos e rosilhos. Lucena et al. (2015LUCENA, J.E.C.; VIANNA, S.A.B.; BERBARI NETO, F. et al. Estudo comparativo das proporções morfométricas entre garanhões e castrados da raça Campolina. Semin. Ciênc. Agrár., v.36, p.353-366, 2015. ) descreveram efeito de pelagem sobre as medidas lineares obtidas no registro em animais da raça Campolina, exceto para CESP, LCAB e PCAN, sendo para castrados o efeito de pelagem verificado somente para CGAR.
Apesar dessa diferença em função da pelagem, isso pode justificar-se, em parte, por variações decorrentes do posicionamento inadequado do hipômetro a campo no momento da aferição das medidas, associado a fatores como índole individual, idade e tipo de manejo adotado. No entanto, em razão do tamanho amostral utilizado nesta pesquisa, é importante retratar o rigor dos processos de seleção dos animais na raça para que a escolha dos indivíduos não seja realizada apenas em função da pelagem, em detrimento das proporções de alturas.
No registro definitivo, houve similaridade entre pelagens (P>0,05) (Tab. 4) para andamento. Esse resultado confirma a atividade dos técnicos a campo e mostra que eles devem se concentrar em avaliar qualidades morfológicas e funcionais, sem influência quanto à pelagem dos animais por ocasião do registro. Todavia, qualidade, comodidade, estilo, regularidade e desenvolvimento podem ser melhorados por treinamento e condicionamento. Da mesma forma, o desempenho em corridas não foi correlacionado com pelagem em Puro Sangue Inglês e Árabe (Stachurska et al., 2007STACHURSKA, A.; PIETA, M.; LOJEK, J.; SZULOWSKA, B. Performance in racehorses of various colours. Livest. Sci., v.106, p.282-286, 2007.). Até o presente, nenhum estudo associou pelagem e características funcionais em equinos marchadores brasileiros.
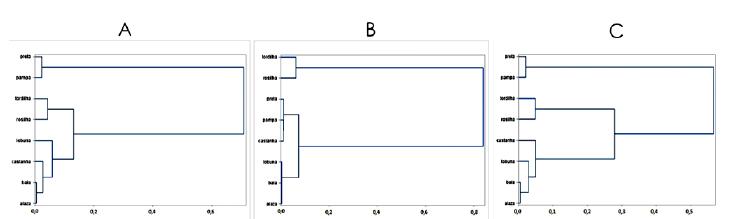
Ao considerar isoladamente medidas lineares, pontuação de andamento e ambas as medidas, foi verificada similaridade entre pelagens (Fig. 3). Para características lineares, formaram-se grupos: preta e pampa; tordilha e rosilha; lobuna, castanha, baia e alazã (Fig. 3a). Para pontuação de andamento, agruparam-se: tordilha e rosilha; preta, pampa e castanha; lobuna, baia e alazã (Fig. 3b). Apesar de pelagens distintas agruparem-se, quando avaliadas ambas as pelagens, os agrupamentos convergem, exceto para castanha. Animais com morfologia similar tendem à funcionalidade semelhante. Para caracteristicas lineares e pontuação, formaram-se grupos iguais aos de morfometria (Fig. 3c). O consenso nos grupos relaciona-se aos valores próximos das medidas. Para pampa e preta, a maioria das variáveis morfométricas foram próximas por médias de quadrados mínimos (Tab. 4) e justifica-se o agrupamento de alazões, baios e lobunos para funcionalidade (Fig. 3b).
Dendograma de agrupamento das diferentes pelagens. A= em função das medidas lineares, B= em função das pontuações de andamento, C= em função das características lineares e das pontuações de andamento.
Dos acasalamentos entre oito pelagens mais frequentes (Tabela 5), resultaram 10.200 fenótipos, sendo 18,62% de baia x alazã; 13,01% de baia x baia; 11,45% de baia x castanha; 8,62% de alazã x castanha; 2,92% de castanha x castanha e 2,51% de alazã x alazã. Os acasalamentos mais comuns por pelagem seguem a ocorrência no Campolina. Apesar de 45 machos rosilhos serem registrados, não aconteceram acasalamentos rosilha x preta e rosilha x rosilha (Tabela 5). Berbari Neto (2005) relatou 33 machos rosilhos inscritos até 2002. Assim, 50% dos produtos destes resultariam em fenótipo igual e há pouca chance de aumentá-lo (Bailey e Brooks, 2013BAILEY, E.; BROOKS, S.A. Horse genetics. 2.ed. London: CABI., 2013. p.42-64.).
Resultados fenotípicos dos acasalamentos observados em equinos da raça Campolina com diferentes pelagens
Produziram-se fenótipos diferentes do esperado de progenitores alazões (Tabela 5), e esses gerariam apenas alazões, segundo Rezende e Costa (2012REZENDE, A.S.C.; COSTA, M.D. Pelagem dos equinos: nomenclatura e genética. 2.ed. Belo Horizonte: FEPMVZ, 2012. 111p.). Esse resultado pode ser decorrente de erros de identificação ao registro. As pelagens equinas derivam de dois pigmentos, eumelanina (gene E_), que origina a tonalidade preta, e feomelanina (ee), que resulta em pelos vermelhos (Rieder, 2009RIEDER, S. Molecular tests for coat colours in horses. J. Anim. Breed. Genet., v.126, p.415-424, 2009.). Poderiam ocorrer filhos de diversas pelagens se um dos progenitores fosse baio, preto ou castanho e classificado erroneamente como alazão. A pelagem lobuna ainda não foi elucidada geneticamente (Rezende e Costa, 2012). Para produtos rosilhos e tordilhos, pode-se inferir negligência em anotar pedigrees e/ou diagnóstico incorreto das pelagens. Quando um dos progenitores era pampa, todos os fenótipos foram possíveis por causa das pelagens de fundo. Equinos pampas filhos de pais não pampas possivelmente ocorreram por falta de identificação de malhas de visualização difícil (Rezende e Costa, 2012). Acasalamentos de tordilhos resultaram em alazã, baia, rosilha e tordilha (Tabela 5) , pois o gene Gray (G) é epistático aos demais (Gremmell, 1939GREMMEL, F. Coat colors in horses. J. Herd, v.30, p.437-445, 1939.; Castle, 1954CASTLE, W.E. Coat color inheritance in horses and in other mammals. Genetic, v.39, p.30, 1954. ).
Essa distribuição de acasalamentos, ocorrência e evolução das pelagens, sendo mais comum baia, alazã e castanha, indicam, conforme Castle (1954CASTLE, W.E. Coat color inheritance in horses and in other mammals. Genetic, v.39, p.30, 1954. ), dispersão dos genes Black (B), Agouti (A) e Dilution (D), presentes desde os primórdios do Campolina. Fenótipos diversos dos padrões de herança genética de pelagens equinas podem associar-se a falhas no pedigree, identificação e/ou transcrição errôneas, além do fator regionalização para denominação das pelagens equinas no país. Para Castle (1954) e Nésio et al. (2004NÉSIO, R.S.; RODRIGUES, C.V.; REZENDE, A.S.C. et al. Análise genética das pelagens dos cavalos registrados na Associação Brasileira dos Criadores do Cavalo Pampa. In: SIMPÓSIO DA SOCIEDADE BRASILEIRA DE MELHORAMENTO ANIMAL, 5., 2004, Pirassununga. Anais... Pirassununga: [s.n], 2004. (Resumo). ), é necessário treinar técnicos de registro e padronizar a classificação de pelagens. Recomenda-se identificação objetiva de pelagens em sistema fenotípico simplificado, para aumentar a acurácia do arquivo genealógico.
CONCLUSÃO
Na raça Campolina, a pelagem baia é prevalente em ambos os sexos. A qualidade da marcha dos indivíduos ao registro não está associada à pelagem, portanto esse fenótipo não deve ser utilizado como principal critério de seleção. Para evitar subjetividades, é necessário padronizar a classificação de pelagens e ajustar as escalas de pontuações de andamento.
AGRADECIMENTOS
À Associação Brasileira dos Criadores do Cavalo Campolina, à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Fundação Capes) e à Fundação de Amparo à Pesquisa de MG (Fapemig) e à Universidade Federal de Minas Gerais.
REFERÊNCIAS
- BAILEY, E.; BROOKS, S.A. Horse genetics. 2.ed. London: CABI., 2013. p.42-64.
- BASTOS, M.S.; REZENDE, M.P.G.; SOUZA, J.C. et al. Levantamento da pelagem e idades de reprodutores Quarto de Milha utilizados na vaquejada em microrregiões do Nordeste do Brasil. Sci. Agrar. Paranaenses, v.16, p.62-68, 2017.
- BERBARI NETO, F. Evolução de medidas lineares e avaliação de índices morfométricos em garanhões da raça Campolina. 2005. 107f. Dissertação (Mestrado em Produção Animal). - Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense, Campos dos Goytacazes, RJ.
- CASTLE, W.E. Coat color inheritance in horses and in other mammals. Genetic, v.39, p.30, 1954.
- FONTES, L.R. Origem e características do cavalo campolina. 1957. 60f. Tese (Professor Catedrático em Zootecnia) - Universidade Federal de Minas Gerais. Belo Horizonte, MG.
- GREMMEL, F. Coat colors in horses. J. Herd, v.30, p.437-445, 1939.
- HENNESSY, K.D.; QUINN, K.M.; MURPHY, J. Producer or purchaser: different expectations may lead to equine wastage and welfare concerns. J. Appl. Anim. Welfare Sci., v.11, p.232-235, 2008.
- LIMA, R.A.S.; CINTRA, A.G. Revisão: estudo do complexo do agronegócio do cavalo. Brasília: Ministério da agricultura pecuária e abastecimento, 2016. p.56.
- LUCENA, J.E.C.; VIANNA, S.A.B.; BERBARI NETO, F. et al. Estudo comparativo das proporções morfométricas entre garanhões e castrados da raça Campolina. Semin. Ciênc. Agrár., v.36, p.353-366, 2015.
- MAIA, F.D.R.B.; SILVA, R.A.; MARACAJÁ, P.B. et al. Caracterização dos tipos de pelagens dos filhos de reprodutores machos “PO” da raça Quarto de Milha em microrregião do sertão Paraibano. Agropecu. Cient. Semiárido, v.9, p.124-131, 2013.
- MICROSOFT access, sistema de gerenciamento de banco de dados relacional para Windows. Guia do access 2007: guia interativo de referências de comando do access 2003 para o access 2007. USA: Microsoft Corporation, 2007.
- NEGRO, S.; IMSLAND, M.; VALERA, M. et al. Association analysis of KIT, MITF, and PAX3 variants with white markings in Spanish horses. Anim. Genet., v48, p.349-352, 2017.
- NÉSIO, R.S.; RODRIGUES, C.V.; REZENDE, A.S.C. et al. Análise genética das pelagens dos cavalos registrados na Associação Brasileira dos Criadores do Cavalo Pampa. In: SIMPÓSIO DA SOCIEDADE BRASILEIRA DE MELHORAMENTO ANIMAL, 5., 2004, Pirassununga. Anais... Pirassununga: [s.n], 2004. (Resumo).
- ORIGEM. Belo Horizonte: ABCCCCampolina, 2018. Disponível em: <http://www.campolina.org.br/portal/>. Acessado em: 15 jan. 2018.
» http://www.campolina.org.br/portal - PROCÓPIO, A.M.; BERGMANN, J.A.G.; COSTA, M.D. Formation and demographic structure of the Campolina horse breed. Arq. Bras. Med. Vet., v.55, p.361-365, 2003.
- REGULAMENTO do serviço de registro genealógico da ABCCCampolina. Belo Horizonte: ABCCCCampolina, 2006. 18p.
- REGULAMENTO do serviço de registro genealógico da ABCCCampolina. Belo Horizonte: ABCCCCampolina, 2017. 18p.
- REGULAMENTO. Registro genealógico do cavalo campolina. Belo Horizonte: ABCCCCampolina, 1938. 10p.
- REZENDE, A.S.C.; COSTA, M.D. Pelagem dos equinos: nomenclatura e genética. 2.ed. Belo Horizonte: FEPMVZ, 2012. 111p.
- RIEDER, S. Molecular tests for coat colours in horses. J. Anim. Breed. Genet., v.126, p.415-424, 2009.
- SAS help and documentation. Version 9.1.3. Cary, NC: SAS Institute Inc., 2014.
- SERVIÇO de registro genealógico. Belo Horizonte: ABCCCCampolina,, 1951. 24p.
- STACHURSKA, A.; PIETA, M.; LOJEK, J.; SZULOWSKA, B. Performance in racehorses of various colours. Livest. Sci., v.106, p.282-286, 2007.
- STATISTICAL analysis system. Version 9.4. Cary: SAS Institute, 2014.
- THIRUVENKADAN, A.K.; KANDASAMY, N.; PANNERSELVAM, S. Coat colour inheritance in horses. Livest. Sci., v.117, p.109-129, 2008.
- VIEIRA, E.R.; REZENDE, A.S.C.; LANA, A.M.Q. et al. Characterization of equidae breeding in Minas Gerais state. Arq. Bras. Med. Vet., v.67, p.319-323, 2015.
Datas de Publicação
-
Publicação nesta coleção
10 Out 2019 -
Data do Fascículo
Jul-Aug 2019
Histórico
-
Recebido
03 Abr 2018 -
Aceito
07 Dez 2018