Resumos
Ordenar espécies em relação a um contínuo de variação de características foliares pode ser útil para inferir sobre a suscetibilidade de uma comunidade a alterações na disponibilidade de recursos e/ou condições ambientais ao longo do tempo. Dez espécies lenhosas de restinga foram acompanhadas quanto à variação temporal de atributos morfológicos foliares. Folhas de dez espécies de restinga foram coletadas nos meses de fevereiro, abril e agosto ao longo de três anos para caracterização da variação temporal da massa de folha por unidade de área (MFA), suculência (SUC), espessura (ESP) e densidade (DEN). As espécies apresentaram uma tendência de valores mais elevados para as características estudadas nos meses mais secos. A partir da comparação sazonal e interanual da MFA, SUC, ESP e DEN sugerimos que a natureza das respostas das plantas à heterogeneidade temporal na disponibilidade de água foi convergente, independentemente do padrão fenológico de produção foliar e filogenia. Portanto, a similaridade observada entre as espécies na dinâmica temporal dos atributos foliares indica a disponibilidade hídrica como um fator determinante para o sucesso na ocupação das planícies arenosas costeiras do norte fluminense.
características foliares; disponibilidade de água; restinga; suculência; variação sazonal
Ranking species in relation to a continuum of leaf-trait variation can be useful to infer community susceptibility to changes in resource availability and/or environmental conditions over time. Leaves of ten woody restinga species were sampled in February, April and August over a period of 3 years to characterize temporal variability of leaf mass per area (LMA), succulence (SUC), thickness (THI) and density (DEN). The species showed a trend toward higher leaf-trait values in dry months during the three-year study. Based on seasonal and interannual comparison of LMA, SUC, THI and DEN we suggest that plant responses to temporal heterogeneity in water availability were convergent, independent of leaf phenological patterns and phylogeny. Thus, the similarity observed among species in temporal dynamics of leaf traits points to water availability as a limiting factor for the success of sandy-coastal-plain occupation in northeastern Rio de Janeiro.
leaf traits; water availability; restinga; succulence; seasonal variation
Variação temporal de características morfológicas de folhas em dez espécies do Parque Nacional da Restinga de Jurubatiba, Macaé, RJ, Brasil
Temporal variation of morphological leaf traits in ten species from Restinga of Jurubatiba National Park, Macaé, Rio de Janeiro State, Brazil
Bruno Henrique Pimentel Rosado; Eduardo Arcoverde de Mattos1 1 Autor para correspondência: eamattos@biologia.ufrj.br
Universidade Federal do Rio de Janeiro, Instituto de Biologia, Departamento de Ecologia, Centro de Ciências da Saúde, C. Postal 68020, 21941-970 Rio de Janeiro, RJ, Brasil (brunorosado@gmail.com)
RESUMO
Ordenar espécies em relação a um contínuo de variação de características foliares pode ser útil para inferir sobre a suscetibilidade de uma comunidade a alterações na disponibilidade de recursos e/ou condições ambientais ao longo do tempo. Dez espécies lenhosas de restinga foram acompanhadas quanto à variação temporal de atributos morfológicos foliares. Folhas de dez espécies de restinga foram coletadas nos meses de fevereiro, abril e agosto ao longo de três anos para caracterização da variação temporal da massa de folha por unidade de área (MFA), suculência (SUC), espessura (ESP) e densidade (DEN). As espécies apresentaram uma tendência de valores mais elevados para as características estudadas nos meses mais secos. A partir da comparação sazonal e interanual da MFA, SUC, ESP e DEN sugerimos que a natureza das respostas das plantas à heterogeneidade temporal na disponibilidade de água foi convergente, independentemente do padrão fenológico de produção foliar e filogenia. Portanto, a similaridade observada entre as espécies na dinâmica temporal dos atributos foliares indica a disponibilidade hídrica como um fator determinante para o sucesso na ocupação das planícies arenosas costeiras do norte fluminense.
Palavras-chaves: características foliares, disponibilidade de água, restinga, suculência, variação sazonal
ABSTRACT
Ranking species in relation to a continuum of leaf-trait variation can be useful to infer community susceptibility to changes in resource availability and/or environmental conditions over time. Leaves of ten woody restinga species were sampled in February, April and August over a period of 3 years to characterize temporal variability of leaf mass per area (LMA), succulence (SUC), thickness (THI) and density (DEN). The species showed a trend toward higher leaf-trait values in dry months during the three-year study. Based on seasonal and interannual comparison of LMA, SUC, THI and DEN we suggest that plant responses to temporal heterogeneity in water availability were convergent, independent of leaf phenological patterns and phylogeny. Thus, the similarity observed among species in temporal dynamics of leaf traits points to water availability as a limiting factor for the success of sandy-coastal-plain occupation in northeastern Rio de Janeiro.
Key words: leaf traits, water availability, restinga, succulence, seasonal variation
Introdução
A natureza das respostas das plantas à variação na disponibilidade de recursos e condições ambientais é, em grande parte, modulada por características morfológicas e fisiológicas (de Mattos et al. 2004). Características foliares têm sido consideradas boas descritoras de processos em vários níveis de organização biológica, desde organismos até ecossistemas (Grime 1977, Reich et al. 1992; Wardle et al. 1998; Garnier et al. 2001; Eviner & Chapin III 2003; Díaz et al. 2004). Além disto, características foliares estão diretamente relacionadas a processos ecossistêmicos, como produtividade e decomposição, devido a sua capacidade de influenciar a intensidade dos fluxos de carbono e nutrientes e os estoques de biomassa nos ecossistemas (Reich et al. 1992; Díaz et al. 2004). Deste modo, a categorização de espécies a partir de características foliares pode fornecer indicativos das estratégias ecológicas apresentadas pelas diferentes espécies em uma dada comunidade (Garnier et al. 2001; Ackerly 2004; Diaz et al. 2004).
Devido ao reconhecimento do significado funcional de características foliares tais como: massa da folha por unidade de área (MFA) (Reich et al. 1992), suculência (SUC) (Kluge & Ting 1978), densidade (DEN) e espessura da lâmina foliar (ESP) (Witkowski & Lamont 1991), estas têm sido amplamente utilizadas em estudos que visam caracterizar o desempenho ecofisiológico de diferentes espécies vegetais ao longo de gradientes de disponibilidade de recursos e condições (Garnier et al. 2001; Wright et al. 2002; Roche et al. 2004). A MFA é um atributo que reflete o custo de construção da folha baseado principalmente em carbono (Eamus et al. 1999; Villar & Merino 2001). Espécies que ocorrem em ambientes pobres em nutrientes e/ou sujeitos a uma menor disponibilidade hídrica tendem a apresentar folhas com maiores valores para MFA (Medina et al. 1990; Wright & Cannon 2001; Niinemets & Kull 2003), longevidade foliar (Chabot & Hicks 1982; Reich et al. 1992) e eficiência no uso de nutrientes (Turner 1994; Wright & Cannon 2001). A SUC, por sua vez, reflete a capacidade de armazenamento de água na folha e altos valores geralmente estão associados a halófitas, xerófitas e plantas que apresentam o metabolismo ácido das crassuláceas (MAC) como Bromeliaceae, Cactaceae e Orchidaceae (Nobel 1988; Lamont & Lamont 2000; Schwinning & Ehleringer 2001). Variações na ESP estão fortemente associadas à SUC. Espécies com folhas mais espessas são comumente encontradas em ambientes com alta intensidade luminosa e/ou pobres em recursos (Niinemets 2001). Um maior número de camadas do parênquima paliçádico, epiderme pluriestratificada, parênquima aqüífero, entre outras características, freqüentemente ocorrem em folhas mais espessas (Lambers et al. 1998). Já uma maior DEN tende a estar relacionada a folhas com uma menor proporção de espaços intercelulares, células menores e em maior número, aumento de fibras e esclereídeos e paredes celulares mais espessas, características estas que reduzem a difusão do CO2 no interior da folha e, por conseguinte, a capacidade fotossintética (Witkowski & Lamont 1991; Niinemets 2001). Por outro lado, espécies com folhas de maior DEN tendem a apresentar uma menor pressão de herbivoria (Turner 1994). Características estruturais, no entanto, devem ser consideradas como uma defesa tardia, isto é, só estão plenamente desenvolvidas nas folhas completamente expandidas.
Com base na funcionalidade para o metabolismo vegetal e na história evolutiva associada a habitats específicos, é possível atribuir um significado adaptativo à posse de determinados estados de atributos morfológicos foliares. Desta maneira, ordenar espécies em relação a um contínuo de variação de características foliares pode ser útil para inferir a suscetibilidade de uma comunidade a alterações na disponibilidade de recursos e/ou condições, que ocorrem em diferentes escalas temporais. No entanto, uma classificação robusta das espécies em função da posse de atributos funcionais depende da amplitude de variação temporal das características em questão e da maneira como as diferentes espécies respondem à heterogeneidade espaço-temporal dos fatores ambientais (Garnier et al. 2001). Modificações na estrutura foliar, que podem ocorrer de maneira idiossincrática entre as espécies, são observadas ao longo da expansão e desenvolvimento das folhas em resposta à variação na disponibilidade de água e nutrientes (Mooney et al. 1978; Cunningham et al. 1999; Prior et al. 2003), intensidade luminosa (Williams et al. 1989; Meziane & Shipley 2001) e herbivoria (Turner 1994). Desta maneira, postulamos que diferenças nos períodos de brotamento e expansão foliar influenciam os padrões de variação temporal de características foliares em uma comunidade. Além disto, a estabilidade da ordenação de espécies ao longo de um contínuo de variação de atributos funcionais dependerá da plasticidade do atributo em questão.
Localizadas marginalmente à Mata Atlântica, as restingas formam um complexo de tipos vegetacionais que ocupam os solos arenosos de origem marinha ao longo do litoral brasileiro (Rizzini 1997; Scarano 2002). Através de uma série de regressões e transgressões marinhas durante o Quaternário, cordões arenosos foram depositados paralelos ao mar formando as planícies costeiras (Suguio & Tessler 1984). Estes ambientes são caracterizados por baixa disponibilidade de recursos, principalmente em função de seus solos arenosos que possuem baixa capacidade de retenção de água e de nutrientes (Henrique et al. 1986; Araujo et al. 2004). Assumimos como premissa que nas restingas maiores valores de MFA, SUC, ESP e DEN tenham um significado funcional, já que estes atributos estão normalmente associados a plantas que ocorrem em ambientes pobres em nutrientes e/ou sujeitos a flutuações na disponibilidade hídrica. Nosso objetivo foi avaliar variação temporal da MFA, SUC, ESP e DEN em uma escala de meses e anos. Além disto, tendo como base a manutenção ao longo do tempo na ordenação das espécies para as características foliares, avaliamos qual atributo foliar é mais adequado para a distinção de dez espécies lenhosas da restinga aberta do Parque Nacional da Restinga de Jurubatiba.
Material e métodos
Local de estudo e espécies vegetais O estudo foi realizado no Parque Nacional da Restinga de Jurubatiba que apresenta (14 140 ha) Macaé, Rio de Janeiro (22º00'- 22º23'S e 41º15'- 41º35'O). A precipitação anual é de 1164 mm com mínima mensal (41 mm) durante o inverno e máxima (189 mm) no verão. A temperatura média anual é de 22,6 ºC (Henriques et al. 1986). A Restinga de Jurubatiba é um mosaico de onze fisionomias vegetais com diferentes espécies e formas de vida e dentre estas fisionomias, a restinga arbustiva aberta de Clusia, onde Clusia hilariana Schltdl. (Clusiaceae) é a espécie mais abundante e a única dentre as demais espécies lenhosas que apresenta o metabolismo ácido das crassuláceas (MAC) (Franco et al. 1996). Esta é uma das fisionomias mais representativas do parque e cobre cerca de 45% da área (Araujo et al. 1998) sendo caracterizada por moitas intercaladas por áreas de areia nua e pelo substrato arenoso, com pouca matéria orgânica e baixa capacidade de retenção de água ou de nutrientes (Henriques et al. 1986).
As espécies estudadas nessa área incluindo C. hilariana foram P. icicariba (DC.) Marchand (Burseraceae), B. sericea DC. (Malpighiaceae), M. parvifolia A.DC (Myrsinaceae), M. subsericea (Mart.) Dubard (Sapotaceae), T. brasiliensis Cambess. (Theaceae), O. notata (Nees) Mez (Lauraceae) E. ovalifolium Peyr. (Erythroxylaceae), E. umbelliflora Berg (Myrtaceae) e M. obtusifolia Mart. (Celastraceae), que estão entre as espécies mais abundantes do parque (Araujo et al. 2004).
Amostragem e acompanhamento temporal dos atributos foliares Para cada espécie foram coletadas 10 folhas já completamente expandidas de 10 indivíduos localizados no segundo cordão entre as lagoas Cabiúnas e Comprida. As coletas de folhas foram realizadas nos mesmos indivíduos nos meses de fevereiro (meio da estação chuvosa), abril (final da estação chuvosa) e agosto (meio da estação seca) nos três anos de estudo. Além disso, cinco ramos de cada indivíduo foram marcados e acompanhados a cada dois meses para a identificação dos períodos de nascimento e morte das folhas entre outubro/2001 e abril/2005.
Com o auxílio de um cortador de metal foi retirado de cada uma das dez folhas um disco foliar de 1,13 cm2 que foi utilizado para determinar os atributos foliares. Os discos foram hidratados em água destilada por um período máximo de 24 horas. Após a hidratação a espessura (mm) foi medida com um paquímetro digital (± 0,01 mm), e a massa saturada com uma balança eletrônica digital (0,001g). Os discos hidratados foram colocados em estufa à 55 ºC por 72 horas para obtenção da massa seca. A partir destes valores, foram calculados a SUC (g m-2) que é o resultado da diferença entre a massa saturada e a massa seca dividida pela área dos discos utilizados (Kluge & Ting 1978), e a MFA (g m-2) que é a razão entre a massa seca dos discos por sua área. Os valores de DEN (mg mm-3) foram obtidos a partir da fórmula: DEN = MFA / ESP (Witkowski & Lamont 1991).
Análises Estatísticas Utilizamos a correlação de Sperman com nível de significância de p < 0,05 para identificar possíveis alterações na ordenação das espécies entre os meses de estudo em função da variação temporal em cada uma das características morfológicas analisadas. Consideramos que as características mais adequadas para representar a ordenação entre as espécies são aquelas que apresentam os maiores coeficientes de correlação entre os períodos de coleta. A grande maioria dos dados mensais de cada característica apresentou distribuição normal (Shapiro-Wilk, p < 0,05; dados não mostrados). No entanto, para uma melhor visualização da dispersão e assimetria dos dados apresentamos os resultados na forma de Box Plots.O programa estatístico utilizado em todas análises foi o Statistica 6.0.
Resultados
Foi possível classificar as espécies em três grupos de acordo com padrão de produção foliar: C. hilariana, B. sericea, M. parvifolia e M. subsericea com produção contínua de folhas ao longo do ano; T. brasiliensis, E. ovalifolium, P. icicariba, E. umbelliflora e O. notata com produção de folhas concentrada na estação chuvosa e M. obtusifolia com produção de folhas concentrada na estação seca (dados não mostrados).
Entre 2003 e 2005 observa-se que os períodos mais secos do ano ocorreram entre os meses de junho e setembro (Fig. 1), quando o total da precipitação mensal foi menor que as médias mensais da temperatura do ar. Os anos de 2003, 2004 e 2005 apresentaram diferenças tanto em relação à precipitação total anual (1.001, 1.319 e 1.388 mm, respectivamente), quanto à precipitação total durante os meses de março a agosto (328, 544 e 555 mm, respectivamente).
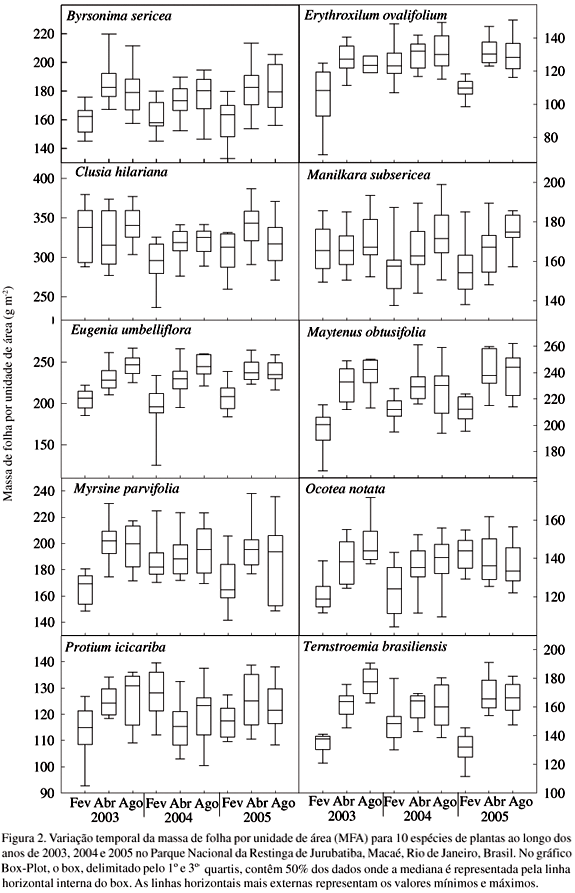
De modo geral, todas as espécies apresentaram uma tendência de valores mais elevados para todas as características foliares nos meses de abril e agosto durante os três anos de estudo (Fig. 2-5). Entre os anos, tanto os valores mínimos quanto os máximos para cada espécie em relação a cada uma das características tenderam a ser bastante similares. MFA e SUC apresentaram as maiores amplitudes de variação sazonal em todas as espécies (Fig 2, 3). Embora também tenham ocorrido tendências de aumentos de ESP e DEN nos meses mais secos, estes atributos não apresentaram grande variação sazonal (Fig. 4, 5). Na maioria dos casos, pode-se observar que de acordo com a amplitude dos valores máximos e mínimos e intervalo entre quartis a variação encontrada dentro de cada mês tendeu a ser maior do que a observada entre os três anos de estudo (Fig. 2, 3, 4, 5).
A ordenação das espécies em relação às características foliares foi bastante similar entre os meses de abril e agosto nos três anos, pois, além de valores iguais a um, foram observados os maiores coeficientes de correlação, na maior parte das comparações (Tab. 1). Dentre as variáveis estudadas, a DEN foi aquela que apresentou um maior número de comparações entre os meses com coeficientes de correlação inferiores a 0,90, o que indica alterações na ordem das espécies entre os meses. Enquanto a MFA e ESP apresentaram várias comparações com coeficientes de correlação menores que 0,96 a SUC só apresentou três comparações inferiores à 0,98, ou seja, uma maior similaridade na ordenação das espécies entre os meses.
Discussão
Diferenças de características associadas à obtenção e uso de recursos entre espécies de uma comunidade têm sido apontadas como um fator que propicia a coexistência de grande número de espécies (Amarasekare 2003). No entanto, baixa disponibilidade de recursos e condições ambientais extremas podem restringir o número de alternativas adaptativas possíveis, resultando em maior similaridade de respostas a estresses de diversas naturezas (Grime 1977). Os solos arenosos da restinga aberta de Clusia não favorecem a retenção de água e nutrientes, de modo que as plantas estão freqüentemente sujeitas a flutuações temporais na disponibilidade de água (de Mattos et al. 2004). Além disso, apesar do padrão de precipitação no Parque Nacional da Restinga de Jurubatiba não ser tão fortemente sazonal como o encontrado no Brasil central, são observados meses de menor precipitação e deficiência hídrica no solo ao longo do inverno (Henriques et al. 1986). Deste modo, apesar das diferenças nos valores absolutos entre as espécies, padrão fenológico de produção foliar e via fotossintética, a forte convergência na dinâmica sazonal observada em características morfológicas tais como: MFA, SUC, ESP e DEN aponta para a variação temporal na disponibilidade de água como um fator determinante para o desempenho ecofisiológico de plantas de restinga.
A tendência de aumento nos valores de cada variável para todas as espécies durante os meses de menor precipitação, indica a ocorrência de mecanismos ligados à resistência à deficiência hídrica nas espécies de restinga. Comparando espécies entre anos com diferentes disponibilidades hídricas, Abrams et al. (1994) verificaram que as espécies de maior MFA apresentaram potenciais hídricos foliares mais negativos, maiores condutâncias estomáticas e pressões osmóticas capazes de manter maiores taxas fotossintéticas em anos com menor precipitação. Além disso, maiores ESP e SUC foliares estão associadas à maior capacidade de estocagem de água o que propiciaria fontes alternativas de água durante períodos de menor disponibilidade hídrica em plantas que ocorrem em ambientes áridos (Lamont & Lamont 2000). Schwinning & Ehleringer (2001) propuseram que em ambientes onde a água é disponibilizada em pulsos, como é o caso das restingas em função principalmente do substrato arenoso, o fenótipo que garantiria maior eficiência entre a manutenção de um balanço hídrico positivo e a capacidade de aquisição de carbono é o de plantas que investem em folhas e ramos com alta capacitância. De maneira similar à SUC e ESP, a tendência de maiores valores de DEN durante os meses de abril e agosto, apesar de menores amplitudes de variação sazonal, pode também estar relacionada com mecanismos que afetam à estocagem de água. Aumentos na DEN podem estar diretamente ligados a: aumentos de fibras e esclereídeos que, por capilaridade, favorecem a retenção de água na folha (Salleo et al. 1997) e maior resistência celular ao murchamento (Oertli et al. 1990). Aumentos na rigidez da parede celular que podem favorecer a sobrevivência das plantas durante períodos secos através do aumento do gradiente entre o potencial hídrico das folhas e do solo, e conseqüente manutenção da absorção de água em situações de menor disponibilidade hídrica (Niinemets 2001). Clusia hilariana, a espécie com maior valor de importância na restinga aberta de Clusia (Araujo et al. 2004) apresenta o metabolismo ácido das crassuláceas (Franco et al. 1996), via fotossintética associada a um uso parsimonioso de água.
A plasticidade fenotípica está associada, entre outros fatores, à previsibilidade ambiental, à disponibilidade de recursos e ao tempo de resposta da planta (Alpert & Simms 2002). Apesar das diferenças de pluviosidade entre os anos a sazonalidade climática na restinga pode ser considerada previsível e gradual. O tipo de resposta observado se mostrou independente tanto da posse de um estado específico dos atributos morfológicos analisados, quanto do padrão fenológico de produção foliar. Portanto, os resultados indicam que, independentemente da filogenia, os maiores valores das características foliares ao longo dos meses mais secos do ano é o fator que define o tipo de resposta à heterogeneidade temporal na disponibilidade de água. Por outro lado, as diferenças observadas entre as espécies nas características foliares analisadas podem influenciar processos relevantes para a planta como o crescimento e a reprodução, bem como os padrões de dominância das espécies na comunidade. Assim, estudos futuros devem elucidar o papel da divergência entre as espécies em termos da capacidade de aquisição de recursos que resultam em um maior ganho de carbono e potencial de crescimento em situações de baixa disponibilidade de recursos e condições ambientais extremas.
Dentre as variáveis estudadas, a SUC foi a característica que apresentou a maior estabilidade temporal na ordenação das espécies, independentemente dos meses e anos de estudo. Quando as diferenças entre as espécies são suficientemente grandes em relação a uma dada característica, a manutenção na ordenação das espécies pode se dar independentemente do tipo de resposta da planta à variação nos fatores ambientais (Garnier et al. 2001). Por outro lado, alterações na ordenação das espécies ocorrem mais freqüentemente quando respostas antagônicas das plantas à heterogeneidade ambiental estão associadas a pequenas diferenças nos atributos entre as espécies (Garnier et al. 2001). Além das espécies terem apresentado a mesma tendência de aumento ao longo dos meses mais secos do ano, quando comparamos as quatro variáveis morfológicas estudadas a SUC foi o atributo que apresentou a maior amplitude de variação entre as espécies. Além disto, mudanças na SUC entre os meses ocorreram na mesma direção para as diferentes espécies, propiciando a maior similaridade na ordenação das espécies entre os meses. Esses resultados são relevantes já que analisamos espécies de diferentes famílias, padrões fenológicos de produção foliar e vias fotossintéticas. A proximidade do mar, que envolve deposição atmosférica de sais, especialmente NaCl, e a presença de solo arenoso podem explicar a relevância funcional da SUC em plantas de restinga.
Portanto, a partir de uma comparação sazonal e interanual da MFA, SUC, ESP e DEN em dez espécies de restinga, evidenciamos que a natureza das respostas das plantas à heterogeneidade temporal na disponibilidade de água foi convergente, independentemente da filogenia, da variação morfológica em atributos foliares, da via fotossintética e do padrão fenológico de produção foliar. A similaridade entre as espécies na dinâmica temporal dos atributos foliares indica que a disponibilidade hídrica é um fator determinante para a ocupação das planícies arenosas costeiras do norte fluminense.
Agradecimentos
A Pedro Cavalin, Leonardo Lima e Maria Isabel Braz, pela indispensável ajuda durante as coletas; à equipe do Núcleo de Pesquisas Ecológicas de Macaé, Universidade Federal do Rio de Janeiro (NUPEM-UFRJ), pelo apoio logístico; ao Programa de Pesquisas Ecológicas de Longa Duração (PELD), FAPERJ e CNPq, pelo financiamento e bolsas de pesquisa; aos dois revisores anônimos pelos comentários e correções do manuscrito. Este trabalho foi desenvolvido dentro do projeto "Seqüestro de Carbono em uma Restinga na Região de Influência da Bacia Petrolífera de Campos", uma parceria entre o Centro de Pesquisas da Petrobras (CENPES) e o Departamento de Ecologia da Universidade Federal do Rio de Janeiro.
Recebido em 25/05/2006. Aceito em 31/01/2007
- Abrams, M.D.; Kubiske, M.E. & Mostoller, S.A. 1994. Relating wet and dry year ecophysiology to leaf structure in contrasting temperate tree species. Ecology 75: 123-133.
- Ackerly, D. 2004. Functional strategies of chaparral shrubs in relation to seasonal water deficit and disturbance. Ecological Monographs 74: 25-44.
- Alpert, P. & Simms, E.L. 2002. The relative advantages of plasticity and fixity in different environments: when is it good for a plant to adjust? Evolutionary Ecology 16: 285-297.
- Amarasekare, P. 2003. Competitive coexistence in spatially structured environments: a synthesis. Ecology Letters 6: 1109-1122.
- Araujo, D.S.D.; Scarano, F.R.; Sá, C.F.C.; Kurtz, B.C.; Zaluar, H.L.T.; Montezuma, R.C.M. & Oliveira, R.C. 1998. Comunidades vegetais do Parque Nacional da Restinga de Jurubatiba. Pp. 39-62. In: F.A. Esteves (ed.). Ecologia das Lagoas Costeiras do Parque Nacional da Restinga de Jurubatiba e do Município de Macaé (RJ) Rio de Janeiro, NUPEM-UFRJ.
- Araujo, D.S.D.; Pereira, M.C.A. & Pimentel, M. 2004. Flora e estrutura de comunidades na Restinga de Jurubatiba - Síntese dos conhecimentos com enfoque especial para a Formação Aberta de Clusia Pp. 59-76. In: C.F.D. Rocha; F.A. Esteves & F.R. Scarano (eds.). Pesquisas de longa duração na restinga de Jurubatiba. Ecologia, história natural e conservação São Carlos, RiMa Editora.
- Chabot, B.F. & Hicks, D.J. 1982. The ecology of leaf life spans. Annual Review Ecology and Systematic 13: 229-259.
- Cunningham, S.A.; Summerhayes, B. & Westoby, M. 1999. Evolutionary divergences in leaf structure and chemistry, comparing rainfall and soil nutrient gradients. Ecological Monographs 69: 569-588.
- de Mattos, E.A.; Braz, M.I.G.; Cavalin, P.O.; Rosado, B.H.P.; Gomes, J.M.; Martins, L.S.T. & Arruda, R.C.O. 2004. Variação espacial e temporal em parâmetros fisioecológicos de plantas. Pp. 99-116. In: C.F.D. Rocha; F.A. Esteves & F.R. Scarano (eds.). Pesquisas de longa duração na restinga de Jurubatiba. Ecologia, história natural e conservação São Carlos, RiMa Editora.
- Díaz, S.; Hodson, J.G.; Thompson, K.; Cabido, M.; Cornelissen, J.H.C.; Jalili, A.; Montserrat-Martí, G.; Grime, J.P.; Zarrinkamar, F.; Asri, Y.; Band, S.R.; Basconcelo, S.; Castro-Díez, P.; Funes, G.; Hamzehee, B.; Khoshnevi, M.; Pérez-Harguindeguy, N.; Pérez-Rontomé, M.C.; Shirvany, F.A.; Vendramini, F.; Yazdani, S.; Abbas-Azimi, R.; Bogaard, A.; Boustami, S.; Charles, M.; Dehghan, M.; Torres-Espuny, L.D.; Falczuk, V.; Guerrero-Campo, J.; Hynd, A.; Jones, G.; Kowsary, E.; Kazemi-Saeed, F.; Maestro-Martínez, M.; Romo-Díez, A.; Shaw, S.; Siavash, B.; Villar-Salvador, P. & Zak, M.R. 2004. The plant traits that drive ecosystems: Evidence from three continents. Journal of Vegetation Science 15: 295-304.
- Eamus, D.; Myers, B.; Duff, G. & Williams, R. 1999. A cost-benefit analysis of leaves of eight Australian savanna species of differing leaf life-span. Photosynthetica 36: 575-586.
- Eviner, V.T. & Chapin III, F.S. 2003. Functional matrix: a conceptual framework for predicting multiple plant effects on ecosystems process. Annual Review Ecology and Systematic 34: 455-485.
- Franco, A.C.; Haag-Kerwer, A.; Herzog, B.; Grams, T.E.E.; Ball, E.; de Mattos, E.A.; Scarano, F.; Garcia, M.A.; Mantovani, A. & Lüttge, U. 1996. The effect of light levels on daily patterns of chlorophyll fluorescence and organic acid accumulation in the tropical CAM tree Clusia hilariana Trees: Structure and Function 10: 359-365.
- Garnier, E.; Laurent, G.; Bellman, A.; Debain, S.; Berthelier, P.; Ducout, B.; Roumet, C. & Navas, M.L. 2001. Consistency of species ordenação based on functional leaf traits. New Phytologist 152: 69-83.
- Grime, J.P. 1977. Evidence for the existence of three primary strategies n plants and its relevance to ecological and evolutionary theory. American Naturalist 111: 169-194.
- Henriques, R.P.B.; Araújo, D.S.D. & Hay, J.D. 1986. Descrição e classificação dos tipos de vegetação da restinga de Carapebús, Rio de Janeiro. Revista Brasileira de Botânica 9: 173-189.
- Kluge, M. & Ting, I.P. 1978. Crassulacean Acid Metabolism: Analysis of an Ecological Adaptation Berlin, Springer-Verlag.
- Lambers, H.; Chapin, F.S. & Pons, T.L. 1998. Plant Physiological Ecology Berlim, Springer-Verlag.
- Lamont, B. & Lamont, H. 2000. Utilizable water in leaves of 8 arid species as derived from pressure-volume curves and chlorophyll fluorescence. Physiologia Plantarum 110: 64-71.
- Medina, E.; Garcia, V. & Cuevas, E. 1990. Sclerophylly and oligotrophic environments: Relationships between leaf structure, mineral nutrient content, and drought resistance in tropical rain forests of upper Rio Negro region. Biotropica 22: 51-64.
- Meziane, D. & Shipley, B. 2001. Direct and indirect relationships between specific leaf area, leaf nitrogen and leaf gas exchange. Effects of irradiance and nutrient supply. Annals of Botany 88: 915-927.
- Mooney, H.A.; Ferrar, P.J. & Slatyer, R.O. 1978. Photosynthetic capacity and carbon allocation patterns in diverse growth forms of Eucalyptus Oecologia 36: 103-111.
- Niinemets, Ü. 2001. Global-scale climatic controls of leaf dry mass per area density, and thickness in trees and shrubs. Ecology 82: 453-469.
- Niinemets, Ü. & Kull, K. 2003. Leaf structure vs. nutrient relationship vary with soil conditions in temperate shrubs and trees. Acta Oecologica 24: 209-219.
- Nobel, P. 1988. Environmental Biology of Agaves and Cacti Cambridge, Cambridge University Press.
- Oertli, J.J.; Lips, S.H. & Agami, M. 1990. The strenght of sclerophyllous cells to resist collapse due to negative turgor pressure. Acta Oecologica 11: 281-289.
- Prior, L.; Eamus, D. & Bowman, D.M.J.S. 2003. Leaf attributes in the seasonally dry tropics: a comparison of four habitats in northern Australia. Functional Ecology 17: 504-515.
- Reich, P.B.; Walters, M.B. & Ellsworth, D.S. 1992. Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems. Ecological Monographs 62: 365-392.
- Rizzini, C.T. 1997. Tratado de Fitogeografia do Brasil Rio de Janeiro, Âmbito Cultural Edições.
- Roche, P.; Díaz-Burlinson & Gachet, S. 2004. Congruency analysis of species ordenação based on leaf traits: which traits are the more reliable. Plant Ecology 174: 37-48.
- Salleo, S.; Nardini, A. & Gullo, M.A.L. 1997. Is sclerophylly of Mediterranean evergreens an adaptation to drought? New Phytologist 135: 603-612.
- Scarano, F.R. 2002. Structure, function and floristic relationships of plant communities in stressful habitats marginal to the Brazilian Atlantic Rainforest. Annals of Botany 90: 517-524.
- Schwinning, S. & Ehleringer, J.R. 2001. Water use trade-offs and optimal adaptations to pulse-driven arid ecosystems. Journal of Ecology 89: 464-480.
- Suguio, K. & Tessler, M.G. 1984. Planícies de cordões litorâneos quaternários do Brasil: origem e nomenclatura. Pp. In: L.D.D. Lacerda; R. Cerqueira & B. Turcq (eds.). Restingas: origem, estrutura, processos Niterói, CEUFF.
- Turner, I.M. 1994. Sclerophylly: primarily protective? Functional Ecology 8: 669-675.
- Villar, R. & Merino, J. 2001. Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems. New Phytologist 151: 213-226.
- Wardle, D.A.; Barker, G.M.; Bonner, K.I. & Nicholson, K.S. 1998. Can comparative approaches based on plant ecophysiological traits predict the nature of biotic interactions and individual plant species effects in ecosystems? Journal of Ecology 86: 405-420.
- Williams, K.; Field, C.B. & Mooney, H.A. 1989. Relationships among leaf construction cost, leaf longevity, and light environment in rain-forest plants of the genus Piper. American Naturalist 133: 198-211.
- Witkowski, E.T.F. & Lamont, B.B. 1991. Leaf specific mass confounds leaf density and thickness. Oecologia 88: 486-493.
- Wright, I.J. & Cannon, K. 2001. Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora. Functional Ecology 15: 351-359.
- Wright, I.J.; Westoby, M. & Reich, P.B. 2002. Convergence towards higher leaf mass per area in dry and nutrient-poor has different consequences for leaf life span. Journal of Ecology 90: 534-453.
Datas de Publicação
-
Publicação nesta coleção
22 Nov 2007 -
Data do Fascículo
Set 2007
Histórico
-
Recebido
24 Maio 2006 -
Aceito
31 Jan 2007