ABSTRACT
Nectar production has an important role in pollinator attraction and successful fruit production in many self-incompatible angiosperm groups. The reproductive biology of Passiflora actinia was studied here and related to nectar dynamics. Passiflora actinia presented a temporal segregation of male and female functions at the beginning of anthesis. Due to the movements of floral verticils, the anthers were positioned in a way that favors pollination two hours before the stigmas reached the same position. The nectary consisted of an epidermis with stomata and a parenchyma rich in starch, which was hydrolyzed during anthesis. The nectary organization is probably associated with the continuous production of nectar during anthesis as well as with the high mean nectar concentration. Hand pollination tests indicated that Passiflora actinia is obligately xenogamous, depending on large bees for pollination, specifically the carpenter bee Xylocopa augusti. The continuous production of nectar may increase the number of bee visits, thus favoring pollen flow.
Keywords:
bees; nectar; nectary; pollination; Xylocopa
Introduction
Nectar production and secretion dynamics has an important role in pollinators attraction and successful reproduction of many allogamous angiosperm groups (Rathcke 1992Rathcke BJ. 1992. Nectar distribution, pollination behavior, and plant reproductive success. In: Hunter MD, Ohgushi T, Price PW. (eds.) Effect of resources distribution on animal-plant interactions. San Diego, Academy Press. p.113-138.; Pacini & Nepi 2007Pacini E, Nepi M. 2007. Nectar production and presentation. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar. Dordrecht, Springer . p. 167-214.). Passiflora is a mostly self-incompatible genus which depends on animal pollination for fruit production both in the wild and cultivated species (Sazima & Sazima 1978Sazima M, Sazima I. 1978. Bat-pollination of the passion flower, Passiflora mucronata, in Southeastern Brazil. Biotropica 10: 100-109.; Bruckner et al. 1995Bruckner CH, Casali VWD, Moraes CF, Redazzi AJ, Silva EAM. 1995. Self-incompatibility in passion fruit (Passiflora edulis Sims). Acta Hoticulturae 370: 45-57.; Koschnitzke & Sazima 1997Koschnitzke C, Sazima M. 1997. Biologia floral de cinco espécies de Passiflora L. (Passifloraceae) em mata semidecídua. Revista Brasileira de Botânica 20: 119-117.; Suassuna et al. 2003Suassuna TMF, Bruckner CH, Carvalho CR, Borém A. 2003. Self-incompatibility in passion fruit: evidence of gametophytic-sporophitic control. Theoretical and Applied Genetics 106: 298-302.; Cobra et al. 2015Cobra SSO, Silva CA, Krause W, Dias DC, Karsburg IV, Miranda AF. 2015. Características florais e polinizadores na qualidade de frutos de cultivares de maracujazeiro‑azedo. Pesquisa Agropecuaria Brasileira 50: 54-62 ). Passiflora exhibits a wide variety of floral forms (Muschner et al. 2003Muschner VC, Lorenz AP, Cervi AC, et al. 2003. A first molecular phylogenetic analysis of Passiflora (Passifloraceae). American Journal of Botany 90: 1229-1238.) and constitutes a monophyletic genus (Muschner et al. 2012Muschner VC, Zamberlan PM, Bonatto SL, Freitas LB. 2012. Phylogeny, biogeography and divergence times in Passiflora (Passifloraceae). Genetics and Molecular Biology 35: 1036-1043.). This variation in floral structures allows the genera to harbor a large number of pollinators that are associated with these differences (Benevides et al. 2013Benevides CR, Evans DM, Gaglianone MC. 2013. Comparing the structure and robustness of Passifloraceae - floral visitor and true pollinator networks in a lowland Atlantic Forest. Sociobiology 60: 295-305.). The genus is rich in bee-pollinated species, including crepuscular species (Janzen 1968Janzen DH. 1968. Reproductive behavior in the Passifloraceae and some of its pollinators in Central America. Behavior 32: 33-48.; Gottsberger et al. 1988Gottsberger G, Camargo JMF, Silberbauer-Gottsberger I. 1988. A bee-pollinated tropical community: the beach dune vegetation of Ilha de São Luís, Maranhão, Brazil. Botanische Jahrbücher Systematisches 109: 469-500.; Sazima & Sazima 1989Sazima I, Sazima M. 1989. Mamangavas e irapuás (Hymenoptera, Apoidea): visitas, interações e conseqüências para a polinização do maracujá (Passifloraceae). Revista Brasileira de Entomologia 33: 109-118.; Koschnitzke & Sazima 1997Koschnitzke C, Sazima M. 1997. Biologia floral de cinco espécies de Passiflora L. (Passifloraceae) em mata semidecídua. Revista Brasileira de Botânica 20: 119-117.; Camillo 2003Camillo E. 2003. A Polinização do Maracujá. Ribeirão Preto, Editora Holos.; Faria & Stehmann 2010Faria FS, Stehmann JR. 2010. Biologia reprodutiva de Passiflora capsularis L. e P. pohlii Mast. (Decaloba, Passifloraceae). Acta Botanica Brasilica 24: 262-269.), but there are also records of bat-pollinated species such as P. galbana and P. mucronata (Sazima & Sazima 1978Sazima M, Sazima I. 1978. Bat-pollination of the passion flower, Passiflora mucronata, in Southeastern Brazil. Biotropica 10: 100-109.; 1987Sazima M, Sazima I. 1987. Additional observations on Passiflora mucronata, the bat-pollinated passionflower. Ciência & Cultura 39: 310-312.; Varassin et al. 2001Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.), hummingbird-pollinated species, like P. speciosa (Varassin et al. 2001Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.) and P. vitifolia (Janzen 1968Janzen DH. 1968. Reproductive behavior in the Passifloraceae and some of its pollinators in Central America. Behavior 32: 33-48.), as well as wasp- and moth- pollinated species (Koschnitzke & Sazima 1997Koschnitzke C, Sazima M. 1997. Biologia floral de cinco espécies de Passiflora L. (Passifloraceae) em mata semidecídua. Revista Brasileira de Botânica 20: 119-117.). Due to this wide range of pollinators, Passiflora species may exhibit variable dynamics of nectar production associated to pollinator behavior and breeding strategies (Varassin et al. 2001Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.).
Among the various parameters related to the quality of nectar as a floral resource, nectar volume and concentration have been the most studied (Petanidou 2007Petanidou T. 2007. Ecological and evolutionary aspects of floral nectars in Mediterranean habitats. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar . Dordrecht, Springer . p. 343-375.). In fact, the dynamics of nectar production are ruled by temporal patterns of nectar secretion and resorption (Pacini & Nepi 2007Pacini E, Nepi M. 2007. Nectar production and presentation. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar. Dordrecht, Springer . p. 167-214.), which can vary the availability of nectar for animals. Nectar-feeding animals are subject to this variability, and the dynamics of nectar presentation may positively or negatively affect the number of visits (Thomson et al. 1989Thomson JD, Mckenna MA, Cruzan MC. 1989. Temporal patterns of nectar and pollen production in Aralia hispida: implications for reproductive success. Ecology 70: 1061-1068.; Rathcke 1992Rathcke BJ. 1992. Nectar distribution, pollination behavior, and plant reproductive success. In: Hunter MD, Ohgushi T, Price PW. (eds.) Effect of resources distribution on animal-plant interactions. San Diego, Academy Press. p.113-138.; Freitas & Sazima 2001Freitas L, Sazima M. 2001. Nectar features in Esterhazya macrodonta, a hummingbird pollinated Scrophulariaceae in Southeastern Brazil. Journal of Plant Research 144: 187-191.).
The patterns of nectar secretion in angiosperms is directly linked to the variety of floral nectary structures (Pacini & Nepi 2007Pacini E, Nepi M. 2007. Nectar production and presentation. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar. Dordrecht, Springer . p. 167-214.) which controls the quantity and quality of nectar production (Pacini & Nepi 2007Pacini E, Nepi M. 2007. Nectar production and presentation. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar. Dordrecht, Springer . p. 167-214.; Nepi & Stpiczyńska 2008Nepi M, Stpiczyńska M. 2008. The complexity of nectar: secretion and resorption dynamically regulate nectar features. Naturwissenschaften 95: 177-84.). The nectar can be exuded from the nectary by epidermal cells or trichomes, pores, rupture or permeability of the cuticle, or by modified stomata, which in the latter case is associated with a specialized parenchyma (Fahn 1979Fahn A. 1979. Ultrastructure of nectaries in relation to nectar secretion. American Journal of Botany 66: 977-985. ). The diversity of forms of nectar exudation is associated with distinct anatomical arrangements of the nectary itself (Nepi 2007Nepi M. 2007. Nectary structure and ultrastructure. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar. Dordrecht, Springer. p. 129-166.) that may be structured or not (Zimmerman 1932Zimmerman J. 1932. Über die extrafloralen Nektarien der Angiospermen. Beihefte zum Botanischen Centralblatt Abt I 49: 99-196). Plants pollinated by animals that require large amounts of nectar usually presents structured nectaries and starch storage in nectary parenchyma to supply nectar secretion at high rates (Pacini & Nepi 2007Pacini E, Nepi M. 2007. Nectar production and presentation. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar. Dordrecht, Springer . p. 167-214.).
We present here a study on the floral biology, nectar production and pollinators of Passiflora actinia seeking to test if nectar dynamics is adjusted to anthesis stages and bee behavioral patterns.
Materials and methods
The study was carried out from August 2007 to March 2008 in fragments of Araucaria Forest or Floresta Ombrofila Mista in the municipality of Curitiba, where the vine Passiflora actinia Hook. is common at the edge of the forest and gaps (Cervi 1997Cervi AC. 1997. Passifloraceae do Brasil. Estudo do gênero Passiflora L., subgênero Passiflora. Fontqueria 45: 1-92.). The climate of the region is classified as Cfb according to the classification of Köppen, which is characterized by temperate climate; average temperature in the coldest month below 18 ºC (mesothermic), with fresh summers, average temperature in the hottest month below 22 ºC and no dry season (IAPAR 2011IAPAR - Instituto Agronômico do Paraná 2011. Cartas climáticas do Paraná. http://www.iapar.br/modules/conteudo/conteudo.php?conteudo=597. 2 Dec. 2011.
http://www.iapar.br/modules/conteudo/con...
). The annual mean temperature is 16-17 °C, the average annual relative humidity varies from 80 to 85 % and the rainfall varies from 1,400 to 1,600 mm (IAPAR 2011IAPAR - Instituto Agronômico do Paraná 2011. Cartas climáticas do Paraná. http://www.iapar.br/modules/conteudo/conteudo.php?conteudo=597. 2 Dec. 2011.
http://www.iapar.br/modules/conteudo/con...
). Vouchers of the studied plants were destroyed during a fire but the plant is fairly common and have been studied before in the same region (Cervi 1997Cervi AC. 1997. Passifloraceae do Brasil. Estudo do gênero Passiflora L., subgênero Passiflora. Fontqueria 45: 1-92.).
The study of floral biology of Passiflora actinia was done during flowering period, which extended from early August to November and fruiting period from October to February, with two fruiting peaks, one in October and one in February. Floral verticils movement were recorded by direct observation during anthesis. Three stigmas of three flowers were collected every two hours to evaluate the onset of stigma receptivity by the catalase test, on which drops of 10 volumes hydrogen peroxide were applied directly on the stigma (Fleet 1952Fleet DS. 1952. Histochemical location of enzymes in vascular plants. Botanical Review 18: 354-398.). At the same time, the presence of pollen in the anthers and odor during the anthesis were recorded. The description of floral morphology followed the terminology adopted by Tillett (1988)Tillett SS. 1988. Passionis passifloris II. Terminologia. Ernestia 48: 1-40..
For the study of the rhythm of nectar production, flowers were bagged at pre-anthesis, and both the cumulative volume of nectar and the dynamic volume of nectar were sampled. To analyze the cumulative volume of nectar, nectar was collected using capillaries at regular intervals, using three to thirteen flowers per each hour interval. To measure the dynamic volume of nectar, four other flowers were used, where the nectar was removed by the same method, at regular intervals, then the flower was bagged again for the next collection. Nectar collections started at 6h15min AM and at each collection, we recorded the temperature and the relative humidity of the environment. In the field, samples were identified, stored at low temperature, taken to the laboratory, where they were frozen for later measurement of volume and concentration. The volume was measured using 50 μL syringes and the total solutes concentration, weight for weight (w.w-1) was measured using a pocket refractometer (ATAGO N-1(). Nectar concentration was calculated using a Conversion Table (ATAGO) for the temperature recorded in the laboratory. The influence of the abiotic variables, such as temperature, relative humidity, and time of day on nectar volume and concentration were analyzed by partial correlation (Zar 1996Zar JA. 1996. Biostatistical analysis. 3th. edn. New Jersey, Prentice-Hall International.). Besides that, relationship among cumulative volume and concentration of nectar along P. actinia anthesis was tested by linear or quadratic regressions (Kleenbaum et al. 1988Kleenbaum DG, Kupper LL, Muller KE. 1988. Applied by regression analysis and other multivariate methods. Boston, PWS-Kent Publishing Company.).
To investigate the structure and the presence of starch in the nectary, slides were mounted with hourly manual longitudinal sections of the nectar chamber, stained with Lugol (Jensen 1962Jensen WA. 1962. Carbohydrates and cell wall constituents. In: Jensen WA. (ed.) Botanical histochemistry: principles and practice. San Francisco, W.H. Freeman. p. 175-208.) from flowers and buds collected and fixed in 50 % alcohol in pre-anthesis, beginning of anthesis, end of anthesis and post-anthesis. In addition, the samples were dehydrated in an ethanol-acetone series, dried at the critical point, gold-coated and the nectary epithelium was observed in a scanning electron microscope, JEOL-6360LV.
The floral visitors were recorded through naked eye observations or with binoculars, throughout the anthesis period, in four nonconsecutive days in a total of 24 hours of observation. The animals that touched the reproductive structures were considered as pollinators. The floral visitors were collected with the aid of an insect net and sacrificed in a lethal chamber, for later identification by consulting biological collections. The relationship among pollinator activity (i.e. number of records) and P. actinia anthesis time was tested by quadratic regression (Kleenbaum et al. 1988Kleenbaum DG, Kupper LL, Muller KE. 1988. Applied by regression analysis and other multivariate methods. Boston, PWS-Kent Publishing Company.).
In order to determine the breeding system and its dependence on pollinators, four controlled pollination treatments (Radford et al. 1974Radford AE, Dickison WC, Massey JR, Bell RC, Seiler MS. 1974. Vascular plant systematics. New York, Harper and Row.) were carried out: 1) spontaneous self-pollination, 2) manual self-pollination, by isolating flowers at pre-anthesis and, during anthesis, by removing pollen from a flower of the same individual and depositing on the stigma of the previously isolated flower, 3) cross-pollination and 4) open pollination (control), only marking buds. Some 35 flowers were used per treatment, except to open-pollination treatments that used 50 flowers. The reproductive success in each treatment was estimated as the ratio between from the number of fruits formed and the number of flowers treated. Fruit-set was compared between pollination treatments using the G test.
Results
Passiflora actinia presented solitary flowers (Fig. 1A), with three purple bracts surrounding the bud. The sepals were white color on the adaxial face and green color on the abaxial face; the petals were white and the fimbriae were white with purple strips, arranged in four series, the outer one larger than the subsequent ones. The operculum (o) was denticulate and reddish purple (Fig. 1A-C). The limen (l), located at the basal portion of the androgynophore, formed along with the operculum a protection of the nectar chamber (Fig. 1B, C). In the nectar chamber, the nectary had an annulus (n), projected downward (Fig. 1C). The reproductive structures, supported by an androginophore (a), consisted of an androecium with five stamens, occasionally six, with rimosas anthers, and a gynoecium with three or occasionally four stigmas, and a superior ovary (Fig. 1A).

Flowers and pollinators of Passiflora actinia. A. flower, highlighting the androgynophore sustaining the reproductive organs; B. limen (1) and operculum (o); C. nectar chamber, with the annulus (n), limen (l) and operculum (o) at the base of the androgynophore (a); D. epidermis of the nectary in the region of the annulus, highlighting the stomata (arrow), MEV, 1310x; pollinators in contact with the reproductive structures: E. Xylocopa augusti and F. X. frontalis.
Anthesis was a gradual process, taking about four hours to reach full flower opening. Anthesis began between 5h00min and 6h00min AM with the upward movement of the sepals that moved independently of each other, followed by the movement of the petals. Sepals and petals deflected until they were parallel to the floral axis, while the fringes remained erect. The anthers presented, at the beginning of anthesis, an introrse position and pollen was already available. The filaments deflected 180 degrees during anthesis, until the anthers were disposed extrorse. This anther position favored the contact with floral visitors when collecting nectar. The gynoecium was the last structure to move. At the beginning of the anthesis the stigmas remained distant from the anthers but already receptive. The styles deflected until the stigmas were positioned between the anthers, about two hours after the anthesis onset (Fig. 1E, F). Due to the asynchronous movements of the filaments and styles, male and female functions of the flowers were separated in time, reducing the occurrence of deposition of self pollen on the stigma surface at the beginning of anthesis. Ten to thirteen hours after the beginning of anthesis, the anthers already showed signs of dehydration, presenting an orange color. Pollen had already been removed, i.e. the anthers were no longer pollen donors. In the same way, the stigma, was dehydrated with reduced volume. At the end of anthesis, the reproductive structures slowly returned to the initial vertical position and the elements of the perianth returned to the shape similar to the bud, but dehydrated. A sweet odor exhaled by flowers was noticed at all observation times.
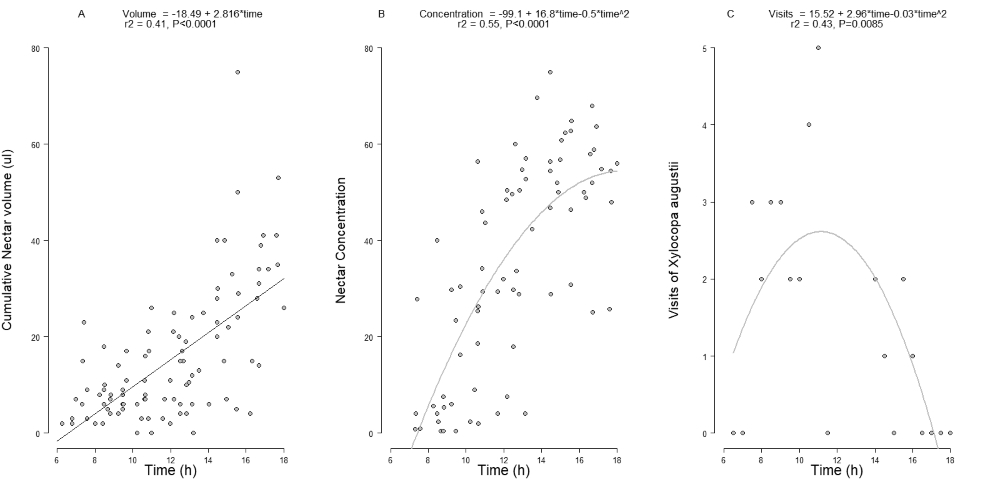
In the cumulative measurement of nectar production, the volume (15.3 ± 13.6 µL) increased with the rise in temperature and along the anthesis (Tab. 1, Fig. 2A) but did not change with the relative humidity (Tab. 1). During these experiments, the temperature was 21.6 ± 7.0 °C and the relative humidity was 57.6 ± 20.1 %. The solute concentration of nectar (35.5 ± 22.0 %) increased with the course of anthesis (Fig. 2B) but did not change with the temperature and relative humidity (Tab. 1). In the measurements of dynamic nectar production, there was no change in volume (4.9 ± 3.9 µL) or solute concentration (15.6 ± 7.9 %) in relation to temperature, relative humidity, nor along anthesis (Tab. 1). During these experiments, the temperature was 21.7 ± 6.9 °C and the relative humidity was 53.0 ± 19.6 %.
Partial correlation between abiotic variables and nectar production in Passiflora actinia. DF = Degrees of freedom; * P < 0.05.

Cumulative nectar volume (A), nectar concentration (B) and visits of Xylocopa augusti (C) along Passiflora actinia anthesis.
The nectary presented stomata in its epidermis (Fig. 1D). Below the epidermis, there was nectary parenchyma, containing many starch grains, followed by a parenchyma rich in intercellular spaces and with some vascular bundles. Buds in pre-anthesis showed large amounts of starch grains in the nectary parenchyma. During the anthesis, the nectary parenchyma showed a reduction in the reaction intensity to Lugol from 12:00 12:00h, and at 16:00h there was hydrolysis of a large part of the starch grains, with a new increase in reaction intensity at 18:00h. However, in post-anthesis, almost all the starch of the nectary parenchyma was hydrolyzed.
Nectar was the only resource used by floral visitors of P. actinia, which was visited by five species of bees, but only large bees were able to pollinate P. actinia (Tab. 2). Due to its abundance, Xylocopa augusti was considered the main pollinator of P. actinia. The foraging activity of X. augusti changed along anthesis, increasing until 10h00min to 11h00min AM and then decreasing (Fig. 2C). Males of X. augusti were observed foraging earlier on the day on P. actinia flowers, while females were observed along the whole anthesis. The large pollinators, Bombus pauloensis, Xylocopa augusti (Fig. 1E) and X. frontalis (Fig. 1F) landed on the inner surface of the fringes, moving to the center of the flower to reach the nectar chamber. When foraging, these bees contacted the anthers and then carried pollen grains on their body. To remove the nectar, the bees forced the androginophore with the head to increase the space between the operculum and the limen so as to allow the introduction of the glossa. In order to collect nectar from the entire chamber, bees moved laterally contacting the stigma or other anthers while circling the flower. Plebeia remota, which is a small species, acted as nectar robber. Apis mellifera when collecting nectar hardly touched the anthers, due to the size of their body, and acted as nectar robber in most interactions. Due to this, A. mellifera pollinated only occasionally, despite its frequency and abundance on P. actina flowers.
Number of records of floral visitors and their role in the pollination of Passiflora actinia.
Passiflora actinia did not set fruits by spontaneous self-pollination or manual self-pollination treatments. Fruit set among cross-pollination and open pollination treatments was similar, 91 % and 88 % fruits/flowers, respectively (G = 0.261, DF = 84, P> 0.05).
Discussion
Passiflora actinia is an outcrossing species pollinated mainly by large bees foraging for nectar. Pollinator foraging activity is probably related to nectar production dynamics, which is continuous during anthesis. The main pollinator, X. augusti, increases its visits to P. actinia flowers from early hours of anthesis, when flowers are mostly acting as pollen donors, until mid-anthesis, when flowers are able to receive pollen deposition. This temporal dicogamy contributes to increase cross-pollination events by long-distance foraging bees.
Passiflora actinia sets fruit only by cross pollination and open-pollination treatments and is thus an outcrossing species. Actually, except small-flowered species of Passiflora (such as P. capsularis (Koschnitzke & Sazima 1997Koschnitzke C, Sazima M. 1997. Biologia floral de cinco espécies de Passiflora L. (Passifloraceae) em mata semidecídua. Revista Brasileira de Botânica 20: 119-117.; Faria & Stehmann 2010Faria FS, Stehmann JR. 2010. Biologia reprodutiva de Passiflora capsularis L. e P. pohlii Mast. (Decaloba, Passifloraceae). Acta Botanica Brasilica 24: 262-269.; Amorim et al. 2011Amorim JS, Souza MM, Viana AJC, Freitas JCO. 2011. Self-, cross- and interspecific pollinations in Passiflora capsularis and P. rubra. Revista Brasileira de Botânica 34: 537-544.), P. rubra (Amorim et al. 2011Amorim JS, Souza MM, Viana AJC, Freitas JCO. 2011. Self-, cross- and interspecific pollinations in Passiflora capsularis and P. rubra. Revista Brasileira de Botânica 34: 537-544.) and P. suberosa (Koschnitzke & Sazima 1997Koschnitzke C, Sazima M. 1997. Biologia floral de cinco espécies de Passiflora L. (Passifloraceae) em mata semidecídua. Revista Brasileira de Botânica 20: 119-117.), most are outcrossers, such as P. alata (Varassin & Silva 1999Varassin IG, Silva AG. 1999. A melitofilia em Passiflora alata Dryander (Passifloraceae), em vegetação de Restinga. Rodriguésia 50: 5-18.), P. edulis (Bruckner et al. 1995Bruckner CH, Casali VWD, Moraes CF, Redazzi AJ, Silva EAM. 1995. Self-incompatibility in passion fruit (Passiflora edulis Sims). Acta Hoticulturae 370: 45-57.; Cobra et al. 2015Cobra SSO, Silva CA, Krause W, Dias DC, Karsburg IV, Miranda AF. 2015. Características florais e polinizadores na qualidade de frutos de cultivares de maracujazeiro‑azedo. Pesquisa Agropecuaria Brasileira 50: 54-62 ), P pohli (Faria & Stehmann 2010Faria FS, Stehmann JR. 2010. Biologia reprodutiva de Passiflora capsularis L. e P. pohlii Mast. (Decaloba, Passifloraceae). Acta Botanica Brasilica 24: 262-269.), P. galbana, P. mucronata, and P. speciosa (Varassin et al. 2001Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.). Indeed, strong genetically controlled self-incompatibility mechanisms have been described for Passiflora species (Bruckner et al. 1995Bruckner CH, Casali VWD, Moraes CF, Redazzi AJ, Silva EAM. 1995. Self-incompatibility in passion fruit (Passiflora edulis Sims). Acta Hoticulturae 370: 45-57.; Suassuna et al. 2003Suassuna TMF, Bruckner CH, Carvalho CR, Borém A. 2003. Self-incompatibility in passion fruit: evidence of gametophytic-sporophitic control. Theoretical and Applied Genetics 106: 298-302.). Besides outcrossing, pollination in P. actinia relies on large bees, which are able to long-distance pollination. This is expected for large-flowered species of Passiflora clade (Muschner et al. 2003Muschner VC, Lorenz AP, Cervi AC, et al. 2003. A first molecular phylogenetic analysis of Passiflora (Passifloraceae). American Journal of Botany 90: 1229-1238.), which tend to be pollinated by larger animals as large bees and vertebrates (Sazima & Sazima 1989Sazima I, Sazima M. 1989. Mamangavas e irapuás (Hymenoptera, Apoidea): visitas, interações e conseqüências para a polinização do maracujá (Passifloraceae). Revista Brasileira de Entomologia 33: 109-118.; Koschnitzke & Sazima 1997Koschnitzke C, Sazima M. 1997. Biologia floral de cinco espécies de Passiflora L. (Passifloraceae) em mata semidecídua. Revista Brasileira de Botânica 20: 119-117.; Varassin et al. 2001Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.; Longo & Fischer 2006Longo JM, Fischer E. 2006. Efeito da taxa de secreção de néctar sobre a polinização e a produção de sementes em flores de Passiflora speciosa Gardn. (Passifloraceae) no Pantanal. Revista Brasileira de Botânica 29: 481-488.; Benevides et al. 2009Benevides CR, Gaglianone MC, Hoffmann M. 2009. Visitantes florais do maracujá-amarelo (Passiflora edulis f. flavicarpa Deg. Passifloraceae) em áreas de cultivo com diferentes proximidades a fragmentos florestais na região Norte Fluminense, RJ. Revista Brasileira de Entomologia 53: 415-421.; Faria & Stehmann 2010Faria FS, Stehmann JR. 2010. Biologia reprodutiva de Passiflora capsularis L. e P. pohlii Mast. (Decaloba, Passifloraceae). Acta Botanica Brasilica 24: 262-269.). On the other hand, in the Decaloba clade, which is characterized by small flowers (Muschner et al. 2003Muschner VC, Lorenz AP, Cervi AC, et al. 2003. A first molecular phylogenetic analysis of Passiflora (Passifloraceae). American Journal of Botany 90: 1229-1238.), pollination by smaller insects predominates (wasps and small bees) (Koschnitzke & Sazima 1997Koschnitzke C, Sazima M. 1997. Biologia floral de cinco espécies de Passiflora L. (Passifloraceae) em mata semidecídua. Revista Brasileira de Botânica 20: 119-117.; Faria & Stehmann 2010Faria FS, Stehmann JR. 2010. Biologia reprodutiva de Passiflora capsularis L. e P. pohlii Mast. (Decaloba, Passifloraceae). Acta Botanica Brasilica 24: 262-269.). Passiflora actinia main pollinator is Xylocopa augusti. Such bees may have long-distance flight capacity, similar to Xylocopa flavorufa up to 6 km range (Pasquet et al. 2008Pasquet RS, Peltier A, Hufford MB, et al. 2008. Long-distance pollen flow assessment through evaluation of pollinator foraging range suggests transgene escape distances. Proceedings of the National Academy of Sciences. 105: 13456-13461.), and Bombus terrestris 15 km range (Goulson & Stout 2001Goulson D, Stout JC. 2001. Homing ability of the bumblebee Bombus terrestris. Apidologie 32: 105-112.), although most flights are on average shorter (Charman et al. 2010Charman TG, Sears J, Green RE, Bourke AFG. 2010. Conservation genetics, foraging distance and nest density of the scarce Great Yellow Bumblebee (Bombus distinguendus). Molecular Ecology 19: 2661-2674.). This long-distance pollen-flow may increase genetic diversity within and between populations through pollination (Ellstrand 1992Ellstrand NC. 1992. Gene flow by pollen: implications for plant conservation genetics. Oikos 63: 77-86.).
Pollinator foraging activity in P. actina flowers throughout the anthesis is probably related to nectar production dynamics, which is continuous during anthesis. This pattern of nectar production is common in Passiflora (Varassin et al. 2001Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.; Longo & Fischer 2006Longo JM, Fischer E. 2006. Efeito da taxa de secreção de néctar sobre a polinização e a produção de sementes em flores de Passiflora speciosa Gardn. (Passifloraceae) no Pantanal. Revista Brasileira de Botânica 29: 481-488.; Benevides et al. 2009Benevides CR, Gaglianone MC, Hoffmann M. 2009. Visitantes florais do maracujá-amarelo (Passiflora edulis f. flavicarpa Deg. Passifloraceae) em áreas de cultivo com diferentes proximidades a fragmentos florestais na região Norte Fluminense, RJ. Revista Brasileira de Entomologia 53: 415-421.; Varassin et al. 2012Varassin IG, Ximenes BMS, Melo GAR, et al. 2012. Produção de néctar e visitas por abelhas em duas espécies cultivadas de Passiflora L. (Passifloraceae). Acta Botanica Brasilica 26: 251-255. ), although species pollinated by bats, with nocturnal anthesis, might produce nectar either before (P. mucronata) or at the beginning of anthesis only (P. galbana) (Varassin et al. 2001Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.). This longer availability of nectar may result in more frequent visits (Rathcke 1992Rathcke BJ. 1992. Nectar distribution, pollination behavior, and plant reproductive success. In: Hunter MD, Ohgushi T, Price PW. (eds.) Effect of resources distribution on animal-plant interactions. San Diego, Academy Press. p.113-138.; Stout & Goulson 2002Stout JC, Goulson D. 2002. The influence of nectar secretion rates on the responses of bumblebees (Bombus spp.) to previously visited flowers. Behavioral Ecology and Sociobiology 52: 239-246.), which should result in increased pollen deposition and reproductive success (Real & Rathcke 1991Real LA, Rathcke BJ. 1991. Individual variation in nectar production and its effect on fitness in Kalmia latifolia. Ecology 72: 149-155.; Longo & Fischer 2006Longo JM, Fischer E. 2006. Efeito da taxa de secreção de néctar sobre a polinização e a produção de sementes em flores de Passiflora speciosa Gardn. (Passifloraceae) no Pantanal. Revista Brasileira de Botânica 29: 481-488.). Actually, Xylocopa augusti increases its visitation to P. actinia flowers along the morning, with few visits in the early morning, when small amounts of nectar are available. At the end of anthesis, X. augusti drops its visitation, probably due to nectar depletion by foraging bees, even if nectar production is still occurring. Since for bees of the genera Xylocopa and Bombus, the frequency of visits is positively related to the amount of available resource (Harder 1990Harder LD. 1990. Pollen removal by bumble bees and its implications for pollen dispersal. Ecology 71: 1110-1125.; Kawai & Kudo 2009Kawai Y, Kudo G. 2009. Effectiveness of buzz pollination in Pedicularis chamissonis: significance of multiple visits by bumblebees. Ecological Research 24: 215-223.), the dynamic of nectar production in P. actinia was expected to be related to pollinator foraging activity. Passiflora species with diurnal anthesis and pollinated by bees, such as P. edulis (Akamine & Girolami 1959Akamine EK, Girolami G. 1959. Pollination and fruit set in the yellow passion fruit. Hawaii Agricultural Experiment Station Technical Bulletin 39: 1-4.; Varassin et al. 2012Varassin IG, Ximenes BMS, Melo GAR, et al. 2012. Produção de néctar e visitas por abelhas em duas espécies cultivadas de Passiflora L. (Passifloraceae). Acta Botanica Brasilica 26: 251-255. ), P. alata (Varassin et al. 2012Varassin IG, Ximenes BMS, Melo GAR, et al. 2012. Produção de néctar e visitas por abelhas em duas espécies cultivadas de Passiflora L. (Passifloraceae). Acta Botanica Brasilica 26: 251-255. ), P. caerulea, P. foetida and P. misera (García & Gottsberger 2009García MTA, Gottsberger G. 2009. Composition of the floral nectar of different subgenera of Argentinian Passiflora species. Plant Systematics and Evolution 283: 133-147.), and hummingbirds, such as P. coccinea (Fischer & Leal 2006Fischer E, Leal IR. 2006. Effect of nectar secretion rate on pollination success of Passiflora coccinea (Passifloraceae) in the Central Amazon. Brazilian Journal of Biology 66: 747-754.) and P. speciosa (Longo & Fischer 2006Longo JM, Fischer E. 2006. Efeito da taxa de secreção de néctar sobre a polinização e a produção de sementes em flores de Passiflora speciosa Gardn. (Passifloraceae) no Pantanal. Revista Brasileira de Botânica 29: 481-488.), pollinator activity ends at the end of nectar production. However, for bat-pollinated species, P. galbana and P. mucronata, nectar production strategies may diverge (Varassin et al. 2001Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.). The observed foraging time differences for males and females of X. augusti is probably not associated to the dynamic of nectar production in P. actinia but to the different requirements associated to territory defense by males. Males of Xylocopa hirsutissima tend to defend territories later on the day, so would forage earlier (Velthius & Camargo 1975Velthius HHW, Camargo JMF. 1975. Further observations on the function of male territories in the carpenter bee Xylocopa (Neoxylocopa) hirsutissima Maidl (Anthophoridae), Hymenoptera. Netherlands Journal of Zoology 25: 616-528.).
The functional dicogamy reported in P. actinia would favor cross-pollination by preventing the pollen grains removed from the flower by X. augusti at the beginning of anthesis to be immediately deposited on their own stigmas (Janzen 1968Janzen DH. 1968. Reproductive behavior in the Passifloraceae and some of its pollinators in Central America. Behavior 32: 33-48.; Sazima & Sazima 1978Sazima M, Sazima I. 1978. Bat-pollination of the passion flower, Passiflora mucronata, in Southeastern Brazil. Biotropica 10: 100-109.; Koschnitzke & Sazima 1997Koschnitzke C, Sazima M. 1997. Biologia floral de cinco espécies de Passiflora L. (Passifloraceae) em mata semidecídua. Revista Brasileira de Botânica 20: 119-117.; Varassin et al. 2001Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.). In addition, this temporal segregation would reduce the deposition of an excess of incompatible pollen (pollen from the individual itself) on the stigmas that could block later fertilization by compatible pollen (pollen clogging), reducing the reproductive success (Webb & Lloyd 1986Webb CJ, Lloyd DG. 1986. The avoidance of interference between the presentation of pollen and stigmas in Angiosperms II. Hercogamy. New Zealand Journal of Botany 24: 163-178. ). The dicogamy is given by the differential movement of the filaments and styles (Endress 1994Endress PK. 1994. Diversity and evolutionary biology of tropical flowers. Cambridge, Cambridge University Press.; Varassin et al. 2001Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.), and is reported for all studied species of the genus (Sazima & Sazima 1989Sazima I, Sazima M. 1989. Mamangavas e irapuás (Hymenoptera, Apoidea): visitas, interações e conseqüências para a polinização do maracujá (Passifloraceae). Revista Brasileira de Entomologia 33: 109-118.; Koschnitzke & Sazima 1997Koschnitzke C, Sazima M. 1997. Biologia floral de cinco espécies de Passiflora L. (Passifloraceae) em mata semidecídua. Revista Brasileira de Botânica 20: 119-117.; Varassin et al. 2001Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.; Longo & Fischer 2006Longo JM, Fischer E. 2006. Efeito da taxa de secreção de néctar sobre a polinização e a produção de sementes em flores de Passiflora speciosa Gardn. (Passifloraceae) no Pantanal. Revista Brasileira de Botânica 29: 481-488.; Benevides et al. 2009Benevides CR, Gaglianone MC, Hoffmann M. 2009. Visitantes florais do maracujá-amarelo (Passiflora edulis f. flavicarpa Deg. Passifloraceae) em áreas de cultivo com diferentes proximidades a fragmentos florestais na região Norte Fluminense, RJ. Revista Brasileira de Entomologia 53: 415-421.; Faria & Stehmann 2010Faria FS, Stehmann JR. 2010. Biologia reprodutiva de Passiflora capsularis L. e P. pohlii Mast. (Decaloba, Passifloraceae). Acta Botanica Brasilica 24: 262-269.).
High rates of nectar production, as observed in P. actinia, are associated with structural characteristics of nectaries, such as the storage of starch in a parenchyma (Pacini & Nepi 2007Pacini E, Nepi M. 2007. Nectar production and presentation. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar. Dordrecht, Springer . p. 167-214.). In Passiflora, starch storage was first described for P. biflora by Durkee et al. (1983Durkee LT. 1983. The Ultrastructure of floral and extrafloral nectaries. In: Bentley B, Elias T. (eds.) The biology of nectaries. New York, Columbia University Press. p. 1-29.) and also observed in P. alata (Varassin & Silva 1999Varassin IG, Silva AG. 1999. A melitofilia em Passiflora alata Dryander (Passifloraceae), em vegetação de Restinga. Rodriguésia 50: 5-18.) and three other Passiflora species (Varassin 1996Varassin IG. 1996. Néctar e voláteis em quatro espécies de Passiflora (L.) (Passifloraceae). MSc Thesis, Universidade Estadual de Campinas, Campinas.). This kind of nectary is distinguished from those in which there is no starch storage and where there is little control over the production of nectar, since the nectar originates either directly from the phloem (Nepi et al. 2001Nepi M, Guarnieri M, Pacini E. 2001. Nectar secretion, reabsorption, and sugar composition in male and female flowers of Cucurbita pepo. International Journal of Plant Sciences 162: 353-358.; Nepi 2007Nepi M. 2007. Nectary structure and ultrastructure. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar. Dordrecht, Springer. p. 129-166.) or associated with chloro-amyloplasts (Nepi 2007Nepi M. 2007. Nectary structure and ultrastructure. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar. Dordrecht, Springer. p. 129-166.). In Passiflora species, nectar production is controlled by hydrolysis of starch stored in the nectary parenchyma (Durkee et al. 1983Durkee LT. 1983. The Ultrastructure of floral and extrafloral nectaries. In: Bentley B, Elias T. (eds.) The biology of nectaries. New York, Columbia University Press. p. 1-29.), a mechanism well described in Cucurbita pepo (Nepi et al. 1996Nepi M, Ciampolini F, Pacini E. 1996. Development and ultrastructure of Cucurbita pepo nectaries of male flowers. Annals of Botany 78: 95-104.). The new starch stock observed in the nectary parenchyma of P. actinia after the anthesis has already been described for C. pepo (Nepi et al. 1996Nepi M, Ciampolini F, Pacini E. 1996. Development and ultrastructure of Cucurbita pepo nectaries of male flowers. Annals of Botany 78: 95-104.). Although no clear pattern of nectar resorption might be deduced from the nectar dynamics in P. actinia, it is possible that the starch stored in plastids in post-anthesis flowers originates from the carbohydrates reabsorbed from previously secreted nectar and are temporarily stored in the parenchyma (Nepi et al. 1996Nepi M, Ciampolini F, Pacini E. 1996. Development and ultrastructure of Cucurbita pepo nectaries of male flowers. Annals of Botany 78: 95-104.). If resorption occurs, this would explain the decrease in nectar concentration at the end of anthesis.
Passiflora actinia is an outcrossing species visited by large bees that, while foraging for nectar, promotes its pollination. The continuous nectar production, that attracts pollinators during the whole anthesis, is regulated by starch hydrolysis in nectary parenchyma, a nectary organization that maintains high rates of nectar production along the anthesis. The nectar dynamics, functional dicogamy, and the temporal pattern of X. augusti visitation reduces self-pollen deposition and increases the chances of cross-pollination.
References
- Akamine EK, Girolami G. 1959. Pollination and fruit set in the yellow passion fruit. Hawaii Agricultural Experiment Station Technical Bulletin 39: 1-4.
- Amorim JS, Souza MM, Viana AJC, Freitas JCO. 2011. Self-, cross- and interspecific pollinations in Passiflora capsularis and P. rubra Revista Brasileira de Botânica 34: 537-544.
- Benevides CR, Gaglianone MC, Hoffmann M. 2009. Visitantes florais do maracujá-amarelo (Passiflora edulis f. flavicarpa Deg. Passifloraceae) em áreas de cultivo com diferentes proximidades a fragmentos florestais na região Norte Fluminense, RJ. Revista Brasileira de Entomologia 53: 415-421.
- Benevides CR, Evans DM, Gaglianone MC. 2013. Comparing the structure and robustness of Passifloraceae - floral visitor and true pollinator networks in a lowland Atlantic Forest. Sociobiology 60: 295-305.
- Bruckner CH, Casali VWD, Moraes CF, Redazzi AJ, Silva EAM. 1995. Self-incompatibility in passion fruit (Passiflora edulis Sims). Acta Hoticulturae 370: 45-57.
- Camillo E. 2003. A Polinização do Maracujá. Ribeirão Preto, Editora Holos.
- Cervi AC. 1997. Passifloraceae do Brasil. Estudo do gênero Passiflora L., subgênero Passiflora Fontqueria 45: 1-92.
- Charman TG, Sears J, Green RE, Bourke AFG. 2010. Conservation genetics, foraging distance and nest density of the scarce Great Yellow Bumblebee (Bombus distinguendus). Molecular Ecology 19: 2661-2674.
- Cobra SSO, Silva CA, Krause W, Dias DC, Karsburg IV, Miranda AF. 2015. Características florais e polinizadores na qualidade de frutos de cultivares de maracujazeiro‑azedo. Pesquisa Agropecuaria Brasileira 50: 54-62
- Durkee LT. 1983. The Ultrastructure of floral and extrafloral nectaries. In: Bentley B, Elias T. (eds.) The biology of nectaries. New York, Columbia University Press. p. 1-29.
- Ellstrand NC. 1992. Gene flow by pollen: implications for plant conservation genetics. Oikos 63: 77-86.
- Endress PK. 1994. Diversity and evolutionary biology of tropical flowers. Cambridge, Cambridge University Press.
- Fahn A. 1979. Ultrastructure of nectaries in relation to nectar secretion. American Journal of Botany 66: 977-985.
- Faria FS, Stehmann JR. 2010. Biologia reprodutiva de Passiflora capsularis L. e P. pohlii Mast. (Decaloba, Passifloraceae). Acta Botanica Brasilica 24: 262-269.
- Fischer E, Leal IR. 2006. Effect of nectar secretion rate on pollination success of Passiflora coccinea (Passifloraceae) in the Central Amazon. Brazilian Journal of Biology 66: 747-754.
- Fleet DS. 1952. Histochemical location of enzymes in vascular plants. Botanical Review 18: 354-398.
- Freitas L, Sazima M. 2001. Nectar features in Esterhazya macrodonta, a hummingbird pollinated Scrophulariaceae in Southeastern Brazil. Journal of Plant Research 144: 187-191.
- García MTA, Gottsberger G. 2009. Composition of the floral nectar of different subgenera of Argentinian Passiflora species. Plant Systematics and Evolution 283: 133-147.
- Gottsberger G, Camargo JMF, Silberbauer-Gottsberger I. 1988. A bee-pollinated tropical community: the beach dune vegetation of Ilha de São Luís, Maranhão, Brazil. Botanische Jahrbücher Systematisches 109: 469-500.
- Goulson D, Stout JC. 2001. Homing ability of the bumblebee Bombus terrestris Apidologie 32: 105-112.
- Harder LD. 1990. Pollen removal by bumble bees and its implications for pollen dispersal. Ecology 71: 1110-1125.
- IAPAR - Instituto Agronômico do Paraná 2011. Cartas climáticas do Paraná. http://www.iapar.br/modules/conteudo/conteudo.php?conteudo=597 2 Dec. 2011.
» http://www.iapar.br/modules/conteudo/conteudo.php?conteudo=597 - Janzen DH. 1968. Reproductive behavior in the Passifloraceae and some of its pollinators in Central America. Behavior 32: 33-48.
- Jensen WA. 1962. Carbohydrates and cell wall constituents. In: Jensen WA. (ed.) Botanical histochemistry: principles and practice. San Francisco, W.H. Freeman. p. 175-208.
- Kawai Y, Kudo G. 2009. Effectiveness of buzz pollination in Pedicularis chamissonis: significance of multiple visits by bumblebees. Ecological Research 24: 215-223.
- Kleenbaum DG, Kupper LL, Muller KE. 1988. Applied by regression analysis and other multivariate methods. Boston, PWS-Kent Publishing Company.
- Koschnitzke C, Sazima M. 1997. Biologia floral de cinco espécies de Passiflora L. (Passifloraceae) em mata semidecídua. Revista Brasileira de Botânica 20: 119-117.
- Longo JM, Fischer E. 2006. Efeito da taxa de secreção de néctar sobre a polinização e a produção de sementes em flores de Passiflora speciosa Gardn. (Passifloraceae) no Pantanal. Revista Brasileira de Botânica 29: 481-488.
- Muschner VC, Lorenz AP, Cervi AC, et al 2003. A first molecular phylogenetic analysis of Passiflora (Passifloraceae). American Journal of Botany 90: 1229-1238.
- Muschner VC, Zamberlan PM, Bonatto SL, Freitas LB. 2012. Phylogeny, biogeography and divergence times in Passiflora (Passifloraceae). Genetics and Molecular Biology 35: 1036-1043.
- Nepi M. 2007. Nectary structure and ultrastructure. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar. Dordrecht, Springer. p. 129-166.
- Nepi M, Ciampolini F, Pacini E. 1996. Development and ultrastructure of Cucurbita pepo nectaries of male flowers. Annals of Botany 78: 95-104.
- Nepi M, Guarnieri M, Pacini E. 2001. Nectar secretion, reabsorption, and sugar composition in male and female flowers of Cucurbita pepo International Journal of Plant Sciences 162: 353-358.
- Nepi M, Stpiczyńska M. 2008. The complexity of nectar: secretion and resorption dynamically regulate nectar features. Naturwissenschaften 95: 177-84.
- Pacini E, Nepi M. 2007. Nectar production and presentation. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar. Dordrecht, Springer . p. 167-214.
- Pasquet RS, Peltier A, Hufford MB, et al 2008. Long-distance pollen flow assessment through evaluation of pollinator foraging range suggests transgene escape distances. Proceedings of the National Academy of Sciences. 105: 13456-13461.
- Petanidou T. 2007. Ecological and evolutionary aspects of floral nectars in Mediterranean habitats. In: Nicolson SW, Nepi M, Pacini E. (eds.) Nectaries and nectar . Dordrecht, Springer . p. 343-375.
- Radford AE, Dickison WC, Massey JR, Bell RC, Seiler MS. 1974. Vascular plant systematics. New York, Harper and Row.
- Rathcke BJ. 1992. Nectar distribution, pollination behavior, and plant reproductive success. In: Hunter MD, Ohgushi T, Price PW. (eds.) Effect of resources distribution on animal-plant interactions. San Diego, Academy Press. p.113-138.
- Real LA, Rathcke BJ. 1991. Individual variation in nectar production and its effect on fitness in Kalmia latifolia Ecology 72: 149-155.
- Sazima I, Sazima M. 1989. Mamangavas e irapuás (Hymenoptera, Apoidea): visitas, interações e conseqüências para a polinização do maracujá (Passifloraceae). Revista Brasileira de Entomologia 33: 109-118.
- Sazima M, Sazima I. 1978. Bat-pollination of the passion flower, Passiflora mucronata, in Southeastern Brazil. Biotropica 10: 100-109.
- Sazima M, Sazima I. 1987. Additional observations on Passiflora mucronata, the bat-pollinated passionflower. Ciência & Cultura 39: 310-312.
- Stout JC, Goulson D. 2002. The influence of nectar secretion rates on the responses of bumblebees (Bombus spp.) to previously visited flowers. Behavioral Ecology and Sociobiology 52: 239-246.
- Suassuna TMF, Bruckner CH, Carvalho CR, Borém A. 2003. Self-incompatibility in passion fruit: evidence of gametophytic-sporophitic control. Theoretical and Applied Genetics 106: 298-302.
- Thomson JD, Mckenna MA, Cruzan MC. 1989. Temporal patterns of nectar and pollen production in Aralia hispida: implications for reproductive success. Ecology 70: 1061-1068.
- Tillett SS. 1988. Passionis passifloris II. Terminologia. Ernestia 48: 1-40.
- Varassin IG. 1996. Néctar e voláteis em quatro espécies de Passiflora (L.) (Passifloraceae). MSc Thesis, Universidade Estadual de Campinas, Campinas.
- Varassin IG, Silva AG. 1999. A melitofilia em Passiflora alata Dryander (Passifloraceae), em vegetação de Restinga. Rodriguésia 50: 5-18.
- Varassin IG, Trigo JR, Sazima M. 2001. The role of nectar production, flower pigments and odour in the pollination of four species of Passiflora (Passifloraceae) in south-eastern Brazil. Botanical Journal of the Linnean Society 136: 139-152.
- Varassin IG, Ximenes BMS, Melo GAR, et al 2012. Produção de néctar e visitas por abelhas em duas espécies cultivadas de Passiflora L. (Passifloraceae). Acta Botanica Brasilica 26: 251-255.
- Velthius HHW, Camargo JMF. 1975. Further observations on the function of male territories in the carpenter bee Xylocopa (Neoxylocopa) hirsutissima Maidl (Anthophoridae), Hymenoptera. Netherlands Journal of Zoology 25: 616-528.
- Webb CJ, Lloyd DG. 1986. The avoidance of interference between the presentation of pollen and stigmas in Angiosperms II. Hercogamy. New Zealand Journal of Botany 24: 163-178.
- Zar JA. 1996. Biostatistical analysis. 3th. edn. New Jersey, Prentice-Hall International.
- Zimmerman J. 1932. Über die extrafloralen Nektarien der Angiospermen. Beihefte zum Botanischen Centralblatt Abt I 49: 99-196
Publication Dates
-
Publication in this collection
02 July 2018 -
Date of issue
Jul-Sep 2018
History
-
Received
12 Apr 2018 -
Accepted
13 June 2018