Resumos
Obesidade hipotalâmica pode ocorrer em humanos e pode ser reproduzida, experimentalmente, por lesão do VMH em ratos. Esta obesidade pode ser revertida por vagotomia troncular (VT), devido à redução da ingestão alimentar e da insulinemia mediada pelo nervo vago. Experimentalmente, a injeção de MSG causa lesão em nível de ARC. O objetivo deste trabalho é avaliar os efeitos do MSG em ratos e se VT os altera. Estudou-se 52 ratos Wistar machos, divididos em dois grupos de 26 animais, um submetido à injeção de MSG na fase neonatal e outro à de solução salina. Aos 30 dias de vida, após nova divisão, obteve-se: grupo MSG, submetido à VT (VTMSG), e outro à laparotomia (LAPMSG); grupo SALINA, submetido à VT (VTSAL), e outro à laparotomia (LAPSAL). Obteve-se peso, CNA e índice de Lee. O consumo alimentar foi obtido dos 30 aos 90 dias de vida. Aos 90 dias, após eutanásia, mensurou-se peso, CNA, índice de Lee e gordura perigonadal. Análise estatística foi realizada pelo "t de Student". Constatou-se que o MSG provoca redução do CNA e aumento do índice de Lee aos 30 dias de vida, e provoca redução do peso e do CNA, aumento do índice de Lee e da gordura perigonadal aos 90 dias e aumento do consumo alimentar dos 30 aos 90 dias de vida. A VT provoca redução do peso, do índice de Lee e da gordura perigonadal, e tendência à redução do CNA no rato injetado com MSG. A VT provoca redução de consumo alimentar nos primeiros 30 dias de pós-operatório, mas com tendência a maior consumo nos 30 dias subseqüentes. Conclui-se que o MSG injetado na fase neonatal provoca aumento do consumo alimentar e da adiposidade e causa redução da estatura e do peso do animal dos 30 aos 90 dias de vida. E que a VT, realizada aos 30 dias de vida, provoca redução do consumo alimentar nos primeiros 30 dias de pós-operatório, da adiposidade e do peso.
Obesidade; Ratos; Glutamato monossódico; Vagotomia
The hypothalamus and autonomic nervous system exert strong influence in food intake. Hypothalamic lesion could cause neuroendocrine obesity. Hypothalamic obesity could be reproduced through ventromedial hypothalamic lesion in animals and attenuated that by subdiafragmatic vagotomy (VT). VT decreased food intake and insulinemia. Injection of monosodium glutamate (MSG) neonatally in rats produces adiposity, hyperphagia, without weight gain because it injures arcuate nucleus of the hypothalamus, site of growth hormone releasing hormone production. The purpose of this paper is to evaluate the MSG effect on weight, stature, Lee index (3 square root body weight/nasoanal length), gonadal fat (GF) and food intake in rats and the VT effect on these parameters. Fifty-two male Wistar rats were used, divided into two groups of twenty-six animals each. SALINA group was subcutaneously injected with saline solution 12.5% (1.25 mg/g body weight/dayly/5days) and MSG group was injected with MSG solution 24% (4 mg/g body weight/dayly/5 days), after the rats were born, in the posterior cervical region without adverse reaction. On the 30th day of life, the animals were divided into four groups and submitted to operation, sham operated groups (LAPSAL and LAPMSG) and vagotomized groups (VTSAL and VTMSG). On operation day, weight, nasoanal length (NAL) and Lee index were obtained. Food intake were obtained from the 30th day until 90th day. On 90th day, the animals were sacrificed and weight, NAL, Lee index and GF were obtained. Statistic analysis was performed by the Student’s t test. NAL was smaller and Lee index was greater in MSG group than in SALINA group on the 30th day. Weight gain and NAL were smaller and Lee index and GF were greater on the 90th day, and food intake from the 30th to 90th day period was greater in LAPMSG group than in LAPSAL group. Weight gain, Lee index and GF were smaller and NAL trended smaller in VTMSG group than in LAPMSG group on the 90th day. Food intake from the 30th to 60th day period was smaller and trended greater from the 60th to 90th day period in VTMSG group than in LAPMSG. Vagal regeneration could explain that food intake inversion. These results suggest that MSG injected neonatally increases food intake and adiposity, and decreases stature and weight gain from the 30th to 90th day period. VT performed on the 30th day decreases food intake in first 30 days and decreases weight gain, Lee index and GF in MSG injected rats.
Obesity; Rats; MSG; Vagotomy
6 ORIGINAL ARTICLE
EFEITO DA VAGOTOMIA TRONCULAR EM RATOS INJETADOS NA FASE NEONATAL COM GLUTAMATO MONOSSÓDICO: ESTUDO BIOMÉTRICO1 1 . Trabalho realizado no Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da Universidade Estadual de Maringá, e no Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da Universidade Federal do Paraná (UFPR). 2. Professor Assistente da Disciplina de Cirurgia do Aparelho Digestivo e Gastroenterologia do Departamento de Medicina da UEM. 3. Professor Adjunto da Disciplina de Cirurgia do Aparelho Digestivo do Departamento de Cirurgia da UFPR. 4. Professor Titular do Departamento de Cirurgia e Coordenador do Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da UFPR. 5. Professor Titular do Departamento de Cirurgia do Setor de Ciências da Saúde da UFPR. 6. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 7. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 8. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 9. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 10. Professor Auxiliar da Disciplina de Patologia do Departamento de Medicina da UEM. 11. Professor Titular do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da UEM.
Fernando de Souza2 1 . Trabalho realizado no Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da Universidade Estadual de Maringá, e no Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da Universidade Federal do Paraná (UFPR). 2. Professor Assistente da Disciplina de Cirurgia do Aparelho Digestivo e Gastroenterologia do Departamento de Medicina da UEM. 3. Professor Adjunto da Disciplina de Cirurgia do Aparelho Digestivo do Departamento de Cirurgia da UFPR. 4. Professor Titular do Departamento de Cirurgia e Coordenador do Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da UFPR. 5. Professor Titular do Departamento de Cirurgia do Setor de Ciências da Saúde da UFPR. 6. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 7. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 8. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 9. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 10. Professor Auxiliar da Disciplina de Patologia do Departamento de Medicina da UEM. 11. Professor Titular do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da UEM.
João Batista Marchesini3 1 . Trabalho realizado no Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da Universidade Estadual de Maringá, e no Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da Universidade Federal do Paraná (UFPR). 2. Professor Assistente da Disciplina de Cirurgia do Aparelho Digestivo e Gastroenterologia do Departamento de Medicina da UEM. 3. Professor Adjunto da Disciplina de Cirurgia do Aparelho Digestivo do Departamento de Cirurgia da UFPR. 4. Professor Titular do Departamento de Cirurgia e Coordenador do Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da UFPR. 5. Professor Titular do Departamento de Cirurgia do Setor de Ciências da Saúde da UFPR. 6. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 7. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 8. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 9. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 10. Professor Auxiliar da Disciplina de Patologia do Departamento de Medicina da UEM. 11. Professor Titular do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da UEM.
Antônio Carlos L. Campos4 1 . Trabalho realizado no Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da Universidade Estadual de Maringá, e no Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da Universidade Federal do Paraná (UFPR). 2. Professor Assistente da Disciplina de Cirurgia do Aparelho Digestivo e Gastroenterologia do Departamento de Medicina da UEM. 3. Professor Adjunto da Disciplina de Cirurgia do Aparelho Digestivo do Departamento de Cirurgia da UFPR. 4. Professor Titular do Departamento de Cirurgia e Coordenador do Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da UFPR. 5. Professor Titular do Departamento de Cirurgia do Setor de Ciências da Saúde da UFPR. 6. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 7. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 8. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 9. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 10. Professor Auxiliar da Disciplina de Patologia do Departamento de Medicina da UEM. 11. Professor Titular do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da UEM.
Osvaldo Malafaia5 1 . Trabalho realizado no Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da Universidade Estadual de Maringá, e no Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da Universidade Federal do Paraná (UFPR). 2. Professor Assistente da Disciplina de Cirurgia do Aparelho Digestivo e Gastroenterologia do Departamento de Medicina da UEM. 3. Professor Adjunto da Disciplina de Cirurgia do Aparelho Digestivo do Departamento de Cirurgia da UFPR. 4. Professor Titular do Departamento de Cirurgia e Coordenador do Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da UFPR. 5. Professor Titular do Departamento de Cirurgia do Setor de Ciências da Saúde da UFPR. 6. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 7. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 8. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 9. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 10. Professor Auxiliar da Disciplina de Patologia do Departamento de Medicina da UEM. 11. Professor Titular do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da UEM.
Orlando G. Monteiro6 1 . Trabalho realizado no Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da Universidade Estadual de Maringá, e no Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da Universidade Federal do Paraná (UFPR). 2. Professor Assistente da Disciplina de Cirurgia do Aparelho Digestivo e Gastroenterologia do Departamento de Medicina da UEM. 3. Professor Adjunto da Disciplina de Cirurgia do Aparelho Digestivo do Departamento de Cirurgia da UFPR. 4. Professor Titular do Departamento de Cirurgia e Coordenador do Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da UFPR. 5. Professor Titular do Departamento de Cirurgia do Setor de Ciências da Saúde da UFPR. 6. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 7. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 8. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 9. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 10. Professor Auxiliar da Disciplina de Patologia do Departamento de Medicina da UEM. 11. Professor Titular do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da UEM.
Fernando B. Ribeiro7 1 . Trabalho realizado no Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da Universidade Estadual de Maringá, e no Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da Universidade Federal do Paraná (UFPR). 2. Professor Assistente da Disciplina de Cirurgia do Aparelho Digestivo e Gastroenterologia do Departamento de Medicina da UEM. 3. Professor Adjunto da Disciplina de Cirurgia do Aparelho Digestivo do Departamento de Cirurgia da UFPR. 4. Professor Titular do Departamento de Cirurgia e Coordenador do Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da UFPR. 5. Professor Titular do Departamento de Cirurgia do Setor de Ciências da Saúde da UFPR. 6. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 7. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 8. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 9. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 10. Professor Auxiliar da Disciplina de Patologia do Departamento de Medicina da UEM. 11. Professor Titular do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da UEM.
Helvércio F. P. Alves8 1 . Trabalho realizado no Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da Universidade Estadual de Maringá, e no Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da Universidade Federal do Paraná (UFPR). 2. Professor Assistente da Disciplina de Cirurgia do Aparelho Digestivo e Gastroenterologia do Departamento de Medicina da UEM. 3. Professor Adjunto da Disciplina de Cirurgia do Aparelho Digestivo do Departamento de Cirurgia da UFPR. 4. Professor Titular do Departamento de Cirurgia e Coordenador do Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da UFPR. 5. Professor Titular do Departamento de Cirurgia do Setor de Ciências da Saúde da UFPR. 6. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 7. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 8. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 9. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 10. Professor Auxiliar da Disciplina de Patologia do Departamento de Medicina da UEM. 11. Professor Titular do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da UEM.
Fernando J. Siroti9 1 . Trabalho realizado no Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da Universidade Estadual de Maringá, e no Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da Universidade Federal do Paraná (UFPR). 2. Professor Assistente da Disciplina de Cirurgia do Aparelho Digestivo e Gastroenterologia do Departamento de Medicina da UEM. 3. Professor Adjunto da Disciplina de Cirurgia do Aparelho Digestivo do Departamento de Cirurgia da UFPR. 4. Professor Titular do Departamento de Cirurgia e Coordenador do Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da UFPR. 5. Professor Titular do Departamento de Cirurgia do Setor de Ciências da Saúde da UFPR. 6. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 7. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 8. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 9. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 10. Professor Auxiliar da Disciplina de Patologia do Departamento de Medicina da UEM. 11. Professor Titular do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da UEM.
Hugo Meister10 1 . Trabalho realizado no Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da Universidade Estadual de Maringá, e no Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da Universidade Federal do Paraná (UFPR). 2. Professor Assistente da Disciplina de Cirurgia do Aparelho Digestivo e Gastroenterologia do Departamento de Medicina da UEM. 3. Professor Adjunto da Disciplina de Cirurgia do Aparelho Digestivo do Departamento de Cirurgia da UFPR. 4. Professor Titular do Departamento de Cirurgia e Coordenador do Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da UFPR. 5. Professor Titular do Departamento de Cirurgia do Setor de Ciências da Saúde da UFPR. 6. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 7. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 8. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 9. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 10. Professor Auxiliar da Disciplina de Patologia do Departamento de Medicina da UEM. 11. Professor Titular do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da UEM.
Paulo C. F. Mathias11 1 . Trabalho realizado no Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da Universidade Estadual de Maringá, e no Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da Universidade Federal do Paraná (UFPR). 2. Professor Assistente da Disciplina de Cirurgia do Aparelho Digestivo e Gastroenterologia do Departamento de Medicina da UEM. 3. Professor Adjunto da Disciplina de Cirurgia do Aparelho Digestivo do Departamento de Cirurgia da UFPR. 4. Professor Titular do Departamento de Cirurgia e Coordenador do Programa de Pós-Graduação em Clínica Cirúrgica, do Setor de Ciências da Saúde da UFPR. 5. Professor Titular do Departamento de Cirurgia do Setor de Ciências da Saúde da UFPR. 6. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 7. Acadêmico do Curso de Medicina do Departamento de Medicina da UEM. 8. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 9. Médico graduado pelo Curso de Medicina do Departamento de Medicina da UEM. 10. Professor Auxiliar da Disciplina de Patologia do Departamento de Medicina da UEM. 11. Professor Titular do Departamento de Biologia Celular e Genética, do Centro de Ciências da Saúde da UEM.
Souza F, Marchesini JB, Campos ACL, Malafaia O, Monteiro OG, Ribeiro FB, Alves HFP, Siroti FJ, Meister H, Mathias PCF. Efeito da vagotomia troncular em ratos injetados na fase neonatal com glutamato monossódico: estudo biométrico. Acta Cir Bras [serial online] 2001 Jan-Mar;16(1). Available from: URL: http://www.scielo.br/acb.
RESUMO: Obesidade hipotalâmica pode ocorrer em humanos e pode ser reproduzida, experimentalmente, por lesão do VMH em ratos. Esta obesidade pode ser revertida por vagotomia troncular (VT), devido à redução da ingestão alimentar e da insulinemia mediada pelo nervo vago. Experimentalmente, a injeção de MSG causa lesão em nível de ARC. O objetivo deste trabalho é avaliar os efeitos do MSG em ratos e se VT os altera. Estudou-se 52 ratos Wistar machos, divididos em dois grupos de 26 animais, um submetido à injeção de MSG na fase neonatal e outro à de solução salina. Aos 30 dias de vida, após nova divisão, obteve-se: grupo MSG, submetido à VT (VTMSG), e outro à laparotomia (LAPMSG); grupo SALINA, submetido à VT (VTSAL), e outro à laparotomia (LAPSAL). Obteve-se peso, CNA e índice de Lee. O consumo alimentar foi obtido dos 30 aos 90 dias de vida. Aos 90 dias, após eutanásia, mensurou-se peso, CNA, índice de Lee e gordura perigonadal. Análise estatística foi realizada pelo "t de Student". Constatou-se que o MSG provoca redução do CNA e aumento do índice de Lee aos 30 dias de vida, e provoca redução do peso e do CNA, aumento do índice de Lee e da gordura perigonadal aos 90 dias e aumento do consumo alimentar dos 30 aos 90 dias de vida. A VT provoca redução do peso, do índice de Lee e da gordura perigonadal, e tendência à redução do CNA no rato injetado com MSG. A VT provoca redução de consumo alimentar nos primeiros 30 dias de pós-operatório, mas com tendência a maior consumo nos 30 dias subseqüentes. Conclui-se que o MSG injetado na fase neonatal provoca aumento do consumo alimentar e da adiposidade e causa redução da estatura e do peso do animal dos 30 aos 90 dias de vida. E que a VT, realizada aos 30 dias de vida, provoca redução do consumo alimentar nos primeiros 30 dias de pós-operatório, da adiposidade e do peso.
DESCRITORES: Obesidade. Ratos. Glutamato monossódico. Vagotomia.
INTRODUÇÃO
O hipotálamo está relacionado com a regulação da ingestão alimentar, balanço energético e o Sistema Nervoso Autônomo (SNA). O hipotálamo pode ser dividido em duas zonas diferentes, de acordo com suas conexões, sendo que a área lateral do hipotálamo (LHA) pertence à zona do SNA parassimpático e a área medial do hipotálamo, principalmente o núcleo ventromedial (VMH), pertence à zona do SNA simpático (1,2). A ingestão alimentar é controlada por estes centros. O VMH seria o centro da saciedade e a LHA o centro da fome. Estímulo elétrico na LHA aumenta a atividade vagal hepática e diminui a atividade da inervação esplâncnica, acompanhado de importante aumento da ingestão alimentar. Por outro lado, a estimulação do VMH aumenta a atividade simpática e reduz os efeitos vagais, provocando intensa hipofagia no animal. Mas, lesão da LHA causa diminuição da ingestão alimentar, do ganho de peso, e perda de tecido adiposo e protéico. O contrário é observado na lesão do VMH, que é acompanhada de hiperfagia em camundongos, ratos, cães, macacos, galinhas e até em humanos (1). Dentre as causas de obesidade neuroendócrina, existe a obesidade por lesão do hipotálamo (3), que pode ser reproduzida experimentalmente em animais. Em humanos, existem situações especiais em que se pode evidenciar a influência do hipotálamo na ingestão alimentar e composição corporal. Desde os relatos de BABINSKI em 1900 e FRÖLICH em 1901, foram descritos mais de cem casos de obesidade associada à lesão hipotalâmica. No homem, as principais causas de lesão hipotalâmica são o traumatismo craniano, neoplasias malignas (craniofaringeoma), meningites, leucemia, alterações anatômicas, alterações vasculares, radioterapia e outras. Nos casos de craniofaringeoma, os pacientes ganham mais de 20 Kg de peso. Acredita-se que o mecanismo comum do ganho de peso seja hiperfagia de início relativamente súbito. Este quadro é acompanhado por hipertrofia do tecido adiposo, hiperinsulinemia intratável e alteração da resposta da secreção do hormônio do crescimento (GH), sob estímulo de injeção de arginina e levodopa. Estes efeitos podem ser explicados pela destruição de áreas do hipotálamo, próximas ao VMH (3). A lesão hipotalâmica pode acompanhar-se de alterações no metabolismo do GH; com isto, pode-se observar as conseqüências deste distúrbio, como: redução da estatura, inadequado desenvolvimento do esqueleto e inadequada mobilização da gordura, que contribui para a adiposidade deste indivíduo. Este fato ocorre porque as células nervosas do núcleo arqueado do hipotálamo (ARC), que produzem hormônio liberador do hormônio do crescimento (GH-RH), são destruídas (4). A obesidade (do latim obesus), definida como aumento da quantidade de gordura corporal acompanhada de aumento de peso corporal, é considerada uma doença universal e de caráter epidêmico, podendo ser considerada problema de saúde pública (3). Para determinar se há obesidade, utilizam-se métodos para mensurar o percentual de gordura corporal. Pode-se utilizar métodos diretos de análise da composição corporal da carcassa, após o óbito do indivíduo, ou métodos indiretos. Dos métodos indiretos, pode-se utilizar os métodos biométricos, baseados em peso, altura e índice de massa corpórea (3,2). Não existe um método ideal para definir qual indivíduo é obeso. O acúmulo de gordura corporal com ou sem aumento de peso corporal é definido como adiposidade; com aumento de peso corporal, obesidade. Experimentalmente, pode-se reproduzir obesidade através de lesão bilateral do VMH, e adiposidade por lesão do ARC por injeção de glutamato monossódico (MSG), que é acompanhada de hipossomatotropismo. O ARC é conhecido como local de secreção de neurotransmissores, peptídeos e de hormônios liberadores hipofisiotróficos como GH-RH, hormônio liberador do hormônio luteinizante, NPY, corticotropina, somatostatina, substância P, prolactina, b-endorfina, dopamina, galamina, neurotensina, polipeptídeo pancreático e pró-encefalina, e a lesão desta região na fase neonatal implica importantes efeitos na função neuroendócrina (5,6,7). Desta forma, o MSG pode ser utilizado como instrumento de investigação do envolvimento do ARC, na função neuroendócrina. Quase todos os modelos de obesidade induzida por lesão hipotalâmica, apresentam hiperinsulinemia de jejum, em função da hiperatividade vagal, causando hipersecreção pancreática de insulina. A vagotomia troncular (VT) subdiafragmática reverte e impede a progressão da obesidade induzida em ratos com lesão do VMH, desde que realizada nas primeiras fases da vida do animal, provocando redução do consumo alimentar e interrupção do estímulo parassimpático para a secreção de insulina em nível pancreático (8). O ARC, principal alvo dos efeitos deletérios do MSG injetado, tem importância fundamental nos estudos de adiposidade e hiperfagia por lesão hipotalâmica. Da mesma forma, sabendo-se que a via comum das principais possíveis causas da obesidade por lesão hipotalâmica são hiperfagia e hiperinsulinemia e, também, sabendo-se que a VT pode reverter estes efeitos nos ratos com lesão do VMH, passamos a realizar a VT nos ratos que foram submetidos à injeção com MSG.
MÉTODOS
Amostra
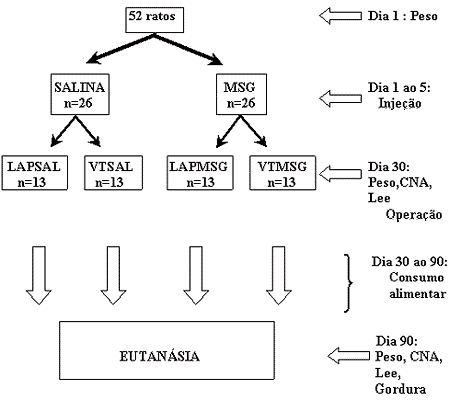
O número total foi de 52 animais da espécie Rattus norvegicus de linhagem Wistar, procedentes do próprio Biotério Central da Universidade Estadual de Maringá. Foram utilizados ratos machos, do 1° ao 5° dia de vida, para injeção de MSG e, a seguir, aos 30 dias de vida, para realização da VT subdiafragmática. O local da experimentação foi o Laboratório de Biologia Celular da Secreção, do Departamento de Biologia Celular e Genética, do Centro de Ciências Biológicas, do Centro de Ciências da Saúde, da Universidade Estadual de Maringá, na cidade de Maringá - PR. As condições de alojamento foram adequadas ao delineamento experimental, dentro do Biotério Setorial, do Departamento de Biologia Celular e Genética. Houve um período de adaptação, utilizando-se para experimentação ratos recém-nascidos de uma mesma linhagem. Os acasalamentos, na proporção de um macho para quatro fêmeas, foram realizados no próprio biotério, aguardando-se o período de gestação, que é de 21 a 25 dias. Após o nascimento, cada rata lactante permaneceu com apenas 6 filhotes, com o objetivo de garantir maior capacidade de lactação para a ninhada, por um período de 21 dias após o nascimento. Foram aproveitados na ninhada o maior número possível de filhotes machos. Os animais foram confinados em gaiolas no padrão do Biotério Setorial do Departamento de Biologia Celular e Genética, com apenas um animal por gaiola, submetidos a um período de luz cíclico de 12 horas (claridade das 7 h às 19 h), em um ambiente arejado, sob controle de temperatura de 23 ± 2 ° C, em ambiente sem ruídos e umidade relativa do ar nos limites padronizados pelo biotério. Os animais foram nutridos administrando-se ração produzida no próprio laboratório e água "ad libidum" (Tabela 1).
O experimento
Os animais obtidos, ratos machos, foram produtos dos acasalamentos no biotério. No primeiro dia de vida, os animais foram submetidos à pesagem, e o peso, entre 5 e 7 g no primeiro dia de vida, foi utilizado como fator de inclusão para o estudo. Ao nascimento, os ratos foram divididos aleatoriamente em dois grupos: SALINA - injeção de solução salina hiperosmótica (n=26); e MSG - injeção de solução de MSG (n=26).
Nos primeiros cinco dias de vida, os animais foram submetidos à injeção de solução de MSG (Aji no moto®) e de solução salina hiperosmótica, de acordo com a divisão dos respectivos grupos. Os ratos do grupo MSG foram submetidos a injeções subcutâneas de solução de MSG na concentração de 24%, com 240 mg por ml de solução, na região cervical posterior, uma vez ao dia, nos primeiros 5 dias de vida, na dose de 4 mg/g de peso corporal. Os ratos do grupo SALINA foram submetidos a injeções subcutâneas de solução salina na concentração de 12,5%, com 125 mg por ml de solução, nos mesmos locais e com a mesma freqüência, na dose de 1,25 mg/g de peso corporal. Os volumes de injeção de solução salina e de MSG foram diferentes para que se pudesse obter o mesmo número de partículas da substância por dose (1,42 Molar). Aos 30 dias de vida, imediatamente antes da operação, os ratos foram submetidos à mensuração do peso e da estatura (CNA). A partir destes dados, obteve-se um índice de massa corpórea, o índice de Lee, que é a razão da raiz cúbica do peso corporal (em gramas) sobre o CNA (em centímetros) multiplicado por 1000 (Figura 1). Neste mesmo dia, os animais foram novamente distribuídos aleatoriamente em quatro grupos: LAPSAL - injeção de solução salina hiperosmótica sem VT (n=13); VTSAL - injeção de solução salina hiperosmótica com VT (n=13); LAPMSG - injeção de solução MSG sem VT (n=13); e VTMSG - injeção de solução MSG com VT (n=13).
Os ratos foram então submetidos à operação, sendo que os ratos dos grupos VTSAL e VTMSG foram submetidos à VT subdiafragmática e os dos grupos LAPSAL e LAPMSG foram submetidos somente à laparotomia exploradora. A partir da operação, o peso corporal e o consumo alimentar dos animais foram obtidos desde os 30 dias até aos 90 dias de vida, individualmente, a cada dois dias. O consumo alimentar foi obtido através da razão da ingestão em gramas sobre o peso de cada animal em quilogramas. Aos 90 dias de vida, foram obtidos peso, CNA e índice de Lee dos animais. Neste mesmo dia, os animais foram submetidos à eutanásia, quando, então, realizou-se laparotomia e dissecou-se a gordura perigonadal para pesagem e obtenção da percentagem desta gordura em relação ao peso corporal do animal.
Técnica operatória Laparotomia exploradora e Vagotomia troncular
Os animais foram anestesiados com quetamina, na dose de 0,05 mg/g de peso corporal, e xilazina, na dose de 0,02 mg/g de peso corporal, por intermédio de injeção intramuscular, no membro inferior. Realizou-se depilação do abdome. O rato foi fixado em plataforma, em decúbito dorsal, com os quatro membros estendidos. Realizou-se anti-sepsia com solução de polivinil-pirrolidona-iodo. Procedeu-se à colocação de campos esterilizados, delimitando a área de trabalho. Utilizou-se material cirúrgico e luvas esterilizados. Realizou-se incisão mediana medindo 4 cm, inferiormente ao esterno. Nos ratos selecionados para realização da laparotomia exploradora exclusiva, após abertura da cavidade peritoneal, procedeu-se ao manuseio dos órgãos e explorou-se a cavidade sem execução de qualquer outro procedimento específico. A seguir, procedeu-se ao fechamento da cavidade abdominal com sutura do plano músculo-aponeurótico e pele com VicrylÒ 000. Nos ratos selecionados para realização de VT, após a abertura da cavidade peritoneal, realizou-se inspeção da cavidade e, a seguir, afastou-se o fígado para a direita e realizou-se secção do ramo hepático do nervo vago anterior. A seguir, observaram-se, na pequena curvatura gástrica, os ramos gástricos anterior e posterior dos respectivos nervos vagos, juntamente com artéria e veia gástricas esquerdas. Identificou-se o esôfago logo abaixo do diafragma, para que fosse reparado com fio de algodão 000. Observaram-se os troncos dos nervos vagos anterior e posterior. O nervo vago direito, ventral ou anterior, divide-se em ramos hepático e gástrico anterior; e o esquerdo, dorsal ou posterior, divide-se, acima do hiato esofágico, em ramos celíaco e gástrico posterior (Figura 2). Os ramos gástricos anterior e posterior dos nervos vagos foram ligados e seccionados juntamente com artéria e veia gástricas esquerdas. Foram obtidos fragmentos para realização de exame histológico (Figura 3). Junto ao esôfago distal, procedeu-se ao reparo dos troncos dos nervos vagos anterior e posterior, logo após a secção do ligamento gastro-esplênico, com especial atenção para se evitar lesão do baço (Figura 4). Próximos ao esôfago distal, os troncos dos nervos vagos e seus ramos foram ligados com fio de algodão 4-0 e seccionados. Também obtiveram-se fragmentos para exame histológico. Procedeu-se à minuciosa dissecção de ramos vagais em nível de fundo gástrico, parede posterior do corpo proximal e na grande curvatura gástrica médio-proximal, para se evitar a possibilidade de VT incompleta. Todos os ramos de nervos vagos observados foram seccionados (Figura 5). Desta forma, eliminou-se a inervação das porções hepática, gástrica e celíaca da distribuição vagal, sempre tomando o cuidado de se prevenir contra uma VT incompleta. A cavidade abdominal foi fechada com sutura do plano músculo-aponeurótico e pele com VicrylÒ 000 (Figura 6).
Após afastamento do fígado e secção do ramo hepático vagal, reparo do esôfago abaixo do diafragma
Pós-operatório
Observou-se a recuperação anestésica no período pós-operatório imediato. Administrou-se ração e água "ad libidum" e observou-se a evolução dos ratos, conforme planejamento do experimento.
Avaliação do experimento
Foram feitas as comparações evolutivas de peso, CNA, índice de Lee, consumo alimentar e acúmulo de tecido gorduroso perigonadal, conforme delineamento do experimento. Os segmentos dos nervos vagos obtidos dos animais dos grupos VTSAL e VTMSG foram enviados para estudo histológico.
Análise estatística
Os resultados são apresentados como média seguida de desvio padrão. Adotou-se a Análise de Variância com dois fatores e, no caso em que a interação se apresentou significativa, adotou-se o "t de Student" para as amostras independentes para as devidas avaliações, levando-se em conta a homogeneidade ou não das variâncias. Em todos os testes, adotou-se um nível de significância de 5%.
RESULTADOS
Peso dos ratos no 1° dia de vida
O peso dos animais dos grupos foram semelhantes no primeiro dia de vida (Tabela 2).
Efeito do MSG sobre peso, CNA e índice de Lee dos ratos no 30° dia de vida
Os ratos MSG apresentaram menor estatura, maior índice de Lee e peso semelhante ao dos ratos SALINA, comparados aos 30 dias de vida (Tabela 3). Aos 30 dias de vida, observou-se a semelhança entre os grupos LAPSAL e VTSAL entre si (Tabela 4), e entre LAPMSG e VTMSG entre si (Tabela 5), caracterizando homogeneidade destas amostras.
Efeito do MSG sobre peso, CNA, índice de Lee e gordura perigonadal dos ratos dos grupos LAPSAL e LAPMSG no 90° dia de vida
Os ratos do grupo LAPMSG apresentaram-se com menor peso, menor estatura, maior índice de Lee e maior percentual de gordura perigonadal que os ratos LAPSAL, aos 90 dias de vida (Tabela 6).
Efeito da vagotomia troncular sobre peso, CNA, índice de Lee e gordura perigonadal dos ratos dos grupos LAPSAL e VTSAL no 90° dia de vida
Os ratos do grupo VTSAL apresentaram-se com menor peso, menor estatura, e índice de Lee e percentual de gordura perigonadal semelhantes aos ratos LAPSAL, aos 90 dias de vida (Tabela 7).
Efeito da vagotomia troncular sobre peso, CNA, índice de Lee e gordura perigonadal dos ratos dos grupos LAPMSG e VTMSG ao 90° dia de vida
Os ratos do grupo VTMSG apresentaram-se com menor peso, menor índice de Lee, menor percentual de gordura perigonadal e uma tendência para menor estatura em relação aos ratos LAPMSG, aos 90 dias de vida (Tabela 8).
Efeito do MSG sobre o consumo alimentar dos ratos dos grupos LAPSAL e LAPMSG
Os ratos do grupo LAPMSG apresentaram maior consumo alimentar que os do grupo LAPSAL, considerando-se o período completo (30° ao 90° dia de vida), o período do 30° ao 60° dia de vida e o do 60° ao 90° dia de vida (Tabela 9) (Gráfico 1).
Efeito da vagotomia troncular sobre o consumo alimentar dos ratos dos grupos LAPSAL e VTSAL
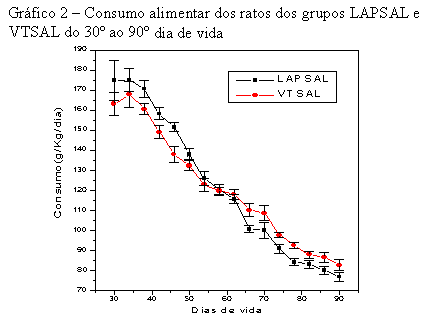
Os ratos do grupo VTSAL apresentaram consumo alimentar semelhante em todo período (30° ao 90° dia de vida), com uma tendência a menor consumo no período do 30° ao 60° dia de vida e um consumo maior no período do 60° ao 90° dia de vida, quando comparados aos do grupo LAPSAL (Tabela 10). A inversão no grupo VTSAL, de menor para maior consumo alimentar, ocorreu no 62° dia de vida (Gráfico 2).
Efeito da vagotomia troncular sobre o consumo alimentar dos ratos dos grupos LAPMSG e VTMSG
Os ratos do grupo VTMSG apresentaram consumo alimentar semelhante em todo período (do 30° ao 90° dia de vida), um menor consumo no período do 30° ao 60° dia de vida e uma tendência a maior consumo no período do 60° ao 90° dia de vida, quando comparados aos do grupo LAPMSG (Tabela 11). Esta inversão no grupo VTMSG, de menor para maior consumo alimentar, ocorreu entre o 58° e 62° dia de vida (Gráfico 3).
Exame histológico dos fragmentos de nervos vagos
Todos os fragmentos de nervos retirados dos ratos do grupo VTSAL e VTMSG foram transportados em solução de formalina, fixados em parafina e corados pelas técnicas de hematoxilina-eosina (HE) e abidina biotina peroxidase (ABP). Todos os materiais examinados à microscopia ótica confirmaram a presença de elementos compatíveis com tecido nervoso.
DISCUSSÃO
O MSG, injetado na fase neonatal, destrói seletivamente os neurônios em nível do ARC, ao contrário da eletrotermocoagulação, que acaba lesando os axônios próximos desta região (7). Não houve observou-se complicações nos locais de injeção das substâncias (9,10,11,12,7,6,4,13,14). Como existe uma resposta dependente do sexo do animal no desenvolvimento das conseqüências da lesão hipotalâmica, optou-se pela amostra exclusivamente composta por ratos machos, com o objetivo de reduzir as variáveis de influência no estudo (11,12,10). Na fase adulta, os ratos machos, injetados com solução de MSG na fase neonatal, apresentam aumento da lipogênese no fígado, retroperitônio e testículos. Nas fêmeas dos ratos, a lipogênese ocorre apenas nos ovários e retroperitônio. O acúmulo de gordura dá-se em razão do aumento da lipogênese "de novo" (síntese de ácidos graxos) e reduzida mobilização de gorduras. Existe uma atividade aumentada da enzima LPL (lipase lipoproteica) no retroperitônio e gônadas, sendo que apenas nos ratos machos observa-se aumento da atividade da LPL no fígado. No tecido gorduroso de retroperitônio e gônadas, observa-se hipertrofia celular, caracterizando adiposidade do tipo hipertrófico (10). Utilizou-se o índice de Lee como um dos parâmetros biométricos, o qual foi criado por LEE para indicar o estado nutricional do animal, e não se correlaciona adequadamente com o percentual de gordura corporal em ratos normais. O CNA é um índice insatisfatório para estimar a massa livre de gordura em animais de mesma idade e de mesma história nutricional (15). Segundo SEPHENS, caso a adiposidade não seja acompanhada por aumento de peso, não haverá correlação de obesidade e índice de Lee, visto que, por definição, obesidade é o aumento da gordura corporal acompanhada de aumento do peso. Apesar disto, nos animais com obesidade devido à lesão hipotalâmica, existe excelente correlação da gordura corporal com o índice de Lee, tanto na fase adulta, quanto na fase de desmame (16,15). COSCINA observou correlação satisfatória do índice de Lee com tecido gorduroso branco perigonadal e tecido gorduroso marrom interescapular (17). Quando a proporção de peso ocupado por gordura fosse relativamente mais elevada que a massa gordurosa livre, haveria uma fidedignidade muito aceitável para o índice de Lee. Este é o padrão de distribuição gordurosa encontrado nos animais com obesidade induzida por lesão hipotalâmica (15). Portanto, apesar de todas as ressalvas conhecidas, com intuito de avaliação propriamente dito, optou-se pela utilização do índice de Lee, juntamente com a mensuração direta da gordura perigonadal.
Os efeitos do MSG em nível de hipotálamo ocorrem pela sua ação neurotóxica causando destruição de 80% a 90% dos neurônios da região do ARC, eminência média e outras áreas cerebrais (8,11,12,9,10,6,7,4,18). Nestes ratos, observa-se redução da capacidade termogênica do tecido adiposo e menor taxa metabólica basal, devido à menor atividade do SNA simpático neste tecido (5,8), além de aumento de triglicerídeos e ácidos graxos no plasma (4). A única síndrome de obesidade conhecida em que o ganho de peso é devido à hiperinsulinemia primária é a de lesão experimental do VMH. Em seres humanos, na obesidade de causa hipotalâmica não há comprovação se a hiperinsulinemia é primária ou secundária à obesidade (2). Apesar da incerteza da presença de danos no VMH, pela proximidade do local da lesão induzida, acredita-se que a hiperatividade parassimpática produzida naquela topografia possa explicar a hiperinsulinemia dos animais injetados com MSG (19). O índice de Lee dos ratos MSG, aos 30 e 90 dias, apresentou-se maior que o dos ratos SALINA devido ao menor CNA. A redução do peso do ratos MSG aos 90 dias, não foi suficiente para reduzir o índice de Lee em relação aos ratos SALINA. Aos 90 dias, os ratos MSG apresentaram-se com menor peso, muito provavelmente devido à menor estatura, apesar do maior consumo alimentar. O grupo LAPMSG apresentou um índice de gordura perigonadal e índice de Lee maiores, podendo-se, então, constatar maior adiposidade destes animais. Maior adiposidade e decréscimo na estatura do animal aos 30 e 90 dias de vida, componentes da síndrome induzida pelo MSG injetado na fase neonatal, na dose referida, também foram obtidos em outros trabalhos (5,8,7,9,10,6). Os animais, submetidos à injeção de MSG na fase neonatal, apresentam diminuição de estatura e de vários órgãos devido ao hipossomatotropismo, com redução dos níveis circulantes de GH, tanto sua secreção basal como pulsátil, em virtude da destruição permanente dos neurônios do ARC, local de produção do GH-RH (18,17,20). Como o GH é hormônio calorífero, que normalmente produz lipólise e anabolismo, além de ser diabetogênico, com a sua deficiência na fase adulta, acarretaria na falha da mobilização das gorduras, com aumento da glicólise e glicogenólise, hipoglicemia de jejum, adiposidade e intolerância ao frio, contribuindo para o aumento do tecido gorduroso destes animais (5,8,18,4). Portanto, a deficiência de GH por lesão hipotalâmica induzida pela injeção do MSG pode participar no mecanismo de adiposidade destes animais (3). Apesar disto, a participação da hiperatividade vagal, com hiperinsulinemia e adiposidade, é provavelmente muito relevante nestes animais. Neste estudo, observou-se redução da adiposidade peri-epididimal e do índice de Lee nos ratos injetados com MSG, após serem vagotomizados. Esta fato evidencia a importância do mecanismo vagal na gênese da adiposidade destes animais, pois, neste experimento, não houve reposição do GH. Neste estudo, foram utilizados ratos injetados com MSG que não foram vagotomizados (grupo LAPMSG), de forma que foi possível isolar a participação vagal neste tipo de adiposidade induzida pelo MSG. Nos ratos injetados com MSG, observa-se hiperinsulinemia, devido ao aumento da secreção de células b das ilhotas pancreáticas, que está associada à lipogênese e atividade da LPL (21). A hiperinsulinemia está relacionada à hipertonicidade vagal, conseqüente da hiperatividade colinérgica hipotalâmica, causada pela injeção do MSG (21,8).
Os ratos MSG apresentaram maior consumo alimentar, em relação aos ratos SALINA, caracterizando-se a hiperfagia destes animais, desde o 30° até o 90° dia de vida, resultados também encontrados por alguns autores (5,19), apesar da discordância de outros autores, afirmando que são hipofágicos quando comparados ao controle, e que a adiposidade, nestes casos, seria exclusivamente metabólica (6,20). Em estudo imunohistoquímico, observou-se que a injeção de MSG na fase neonatal causa desaparecimento dos corpos celulares positivos para NPY no ARC, além da redução destes corpos celulares no PVN (22). Normalmente, o NPY é encontrado no ARC e no PVN (23). Com a lesão do ARC, pela injeção do MSG, a secreção ficaria a cargo do PVN, ou seja, não ocorre a eliminação completa do NPY neste nível. Sabe-se que o NPY injetado diretamente no PVN induz hiperfagia, redução do gasto energético, obesidade e hiperinsulinemia (23,1), acompanhado de aumento da lipogênese hepática, de maior atividade da LPL e de maior captação de glicose pelo adipócito (1). O NPY está mais relacionado com a ingestão e utilização dos carbohidratos. Nos ratos, injetados com MSG, a hiperfagia poderia ser atribuída, inclusive, à participação do NPY remanescente. A leptina, um peptídeo de 16 kD com função hormonal, é secretada por adipócitos do tecido gorduroso branco, e está relacionada com a regulação da ingestão alimentar e balanço energético em nível de SNC. Postula-se a existência de receptores com RNA mensageiro para leptina no hipotálamo, mais exatamente no ARC, como potencial local de ação da mesma. Como o ARC é afetado pela injeção de MSG, com destruição dos receptores de leptina a este nível, haveria uma atenuação no feedback gerado pela leptina circulante, explicando um possível mecanismo de ação do MSG. Habitualmente, a leptina pode suprimir o ganho de peso, diminuir o depósito de gordura corporal e perigonadal, reduzir a ingestão alimentar noturna e reduzir a insulinemia em ratos normais, mas isto não ocorre nos ratos injetados com MSG. Os ratos, injetados com MSG, apresentam-se com elevados níveis de leptina circulantes e desenvolvem adiposidade intensa, mesmo com a administração exógena de leptina, e mesmo pela ação da leptina endógena secretada pelos adipócitos. A leptina não seria efetiva na redução da ingestão alimentar nos animais injetados com MSG, pois não haveria receptores íntegros em nível de ARC. Outro possível mecanismo que estaria afetado nestes ratos, injetados com MSG, seria o poder da leptina em suprimir o apetite através da inibição da ação ou da síntese do NPY. Este mecanismo estaria afetado porque haveria uma deficiência de NPY no ARC, apesar de uma relativa eficiência do NPY em nível de PVN, o qual não é afetado pelo MSG (4).
O índice de Lee não apresentou-se diferente nos ratos controle vagotomizados, porque houve redução do CNA e também do peso. Estas reduções, provavelmente, foram devidas ao menor consumo alimentar nos primeiros 32 dias de pós-operatório. A VT não influiu na concentração de gordura perigonadal do rato normal. A partir do 32° dia após a VT, observou-se uma tendência a menor consumo alimentar. Em ratos normais, a VT bloqueia o reflexo orofaríngeo alimentar, impedindo hiperinsulinemia sob estímulo do alimento na orofaringe (24), mas não afeta, significativamente, o fluxo sangüíneo de ilhotas pancreáticas e insulinemia basal (25,26). A VT, em ratos normais, provoca hipofagia e perda de peso, dependente da idade e peso inicial, distensão, impacção e estase gástrica importantes (29), aversão à ingestão alimentar, náuseas, disfagia, dificuldade de esvaziamento gástrico, redução da habilidade de digestão alimentar e desconforto visceral nos ratos (13,26). Estes efeitos são mais evidentes em ratos com lesão hipotalâmica (13,27). CAMPFIELD observou que a VT promove perda de peso e redução de ingestão alimentar de forma mais importante nos primeiros 21 dias de pós-operatório de VT (28). OPSAHL e POWLEY observaram distensão gástrica constante e conteúdo gástrico alimentar presente após um período de 24 horas de jejum (27). Uma das explicações para este fato, é que a VT bloqueia o efeito inibitório alimentar (saciedade induzida) da distensão gástrica. Estes animais ingerem menores quantidades e mais freqüentes porções de dieta líquida, e maiores quantidades e menos freqüentes porções de dieta sólida, provavelmente, devido a redução do tônus do esfíncter pilórico. Com isto, tendem a reter alimentos sólidos por até 48 horas após a última ingestão. E, também, tendem a esvaziar mais rapidamente o estômago quando ingerem alimentos líquidos ("dumping syndrome"), o que provoca afluxo de plasma e líquido intersticial para o lume intestinal. Este fato provoca uma elevação na insulinemia em resposta ao rápido esvaziamento gástrico de carbohidratos. Ambas as situações causam aversão alimentar ao animal (30). O menor índice de Lee dos animais do grupo VTMSG, foi devido à redução do peso dos mesmos, apesar de uma tendência à menor estatura. Os ratos MSG, submetidos a VT, apresentaram hipofagia nos primeiros 30 dias de pós-operatório e significativa redução do percentual de gordura perigonadal. A partir destas observações, pode-se inferir que a redução de peso, mais especificamente da massa gordurosa destes animais, ocorre por dois possíveis mecanismos. Um deles, seria a redução da ingestão alimentar, pelo menos nas primeiras quatro semanas de pós-operatório, e o outro, seria a interrupção do estímulo vagal ao pâncreas, reduzindo a hiperinsulinemia e, por sua vez, a deposição gordurosa.
Nos ratos, com lesão bilateral do VMH, a VT abole a hiperinsulinemia de jejum (26,30), reduz a hiperacidez gástrica (26), reduz o fluxo sangüíneo mesentérico, inclusive em ilhotas pancreáticas (25), reduz a motilidade intestinal e secreção de enzimas e hormônios, diminui a ingestão alimentar e reduz o peso dos ratos (13), ou seja, gera um desequilíbrio no SNA revertendo a obesidade destes ratos (26). Redução do índice de Lee e do percentual de gordura perigonadal foram observadas nos ratos MSG e submetidos a VT, demonstrando uma correlação significativa de adiposidade nestes valores, inferindo que a VT poderia reduzir o componente adiposo nestes ratos, através de interrupção de mecanismo vagotônico hiperinsulinêmico (25). O consumo alimentar nos primeiros 30 dias após a VT foi menor, mais significativamente nos ratos MSG, e entre 30 e 60 dias foi maior, mais significativamente nos ratos SALINA. A redução da ingestão alimentar na fase inicial da VT, e, a seguir, um aumento da ingestão, poderia ser explicado pela regeneração da inervação vagal, e uma tentativa de compensação da primeira fase. Outra prova deste fato, é que a VT, realizada antes da lesão hipotalâmica, produz poucos efeitos sobre a mesma, e a explicação é dada pela regeneração das fibras vagais (13). Em nosso estudo, o efeito do MSG, além da participação do nervo vago, tem a participação do hipossomatotropismo, que contribui para obesidade e não pode ser revertida ou atenuada pela VT. Contudo a VT exerce importante papel na atenuação do acúmulo de gordura quando se verificam seus efeitos, muito mais intensos e marcantes sobre o rato injetado com MSG, quando comparado ao rato normal vagotomizado.
CONCLUSÃO
1- A injeção de MSG em ratos, na fase neonatal, não altera o peso, produz redução do CNA e aumento do índice de Lee aos 30 dias de vida.
2- A injeção de MSG em ratos, na fase neonatal, produz redução do peso, redução do CNA, aumento do índice de Lee e aumento do percentual de gordura perigonadal aos 90 dias de vida.
3- A VT subdiafragmática, realizada aos 30 dias de vida, em ratos injetados com MSG na fase neonatal, produz redução do peso, uma tendência à redução do CNA, bem como redução do índice de Lee e do percentual de gordura perigonadal aos 90 dias de vida.
4- A injeção de MSG em ratos, na fase neonatal, produz aumento do consumo alimentar total no período dos 30 aos 90 dias, bem como nos períodos dos 30 aos 60 dias e dos 60 aos 90 dias de vida.
5- A VT subdiafragmática, realizada aos 30 dias de vida, em ratos injetados com MSG na fase neonatal, não altera o consumo alimentar total no período dos 30 aos 90 dias de vida, porém produz redução do consumo alimentar total no período dos 30 aos 60 dias e, além disso, produz uma tendência a aumento de consumo alimentar total dos 60 aos 90 dias de vida.
Souza F, Marchesini JB, Campos ACL, Malafaia O, Monteiro OG, Ribeiro FB, Alves HFP, Siroti FJ, Meister H, Mathias PCF. Effect of vagotomy in rats neonatally injected with monosodium glutamate: biometry study. Acta Cir Bras [serial online] 2001 Jan-Mar;16(1). Available from: URL: http://www.scielo.br/acb.
ABSTRACT: The hypothalamus and autonomic nervous system exert strong influence in food intake. Hypothalamic lesion could cause neuroendocrine obesity. Hypothalamic obesity could be reproduced through ventromedial hypothalamic lesion in animals and attenuated that by subdiafragmatic vagotomy (VT). VT decreased food intake and insulinemia. Injection of monosodium glutamate (MSG) neonatally in rats produces adiposity, hyperphagia, without weight gain because it injures arcuate nucleus of the hypothalamus, site of growth hormone releasing hormone production. The purpose of this paper is to evaluate the MSG effect on weight, stature, Lee index (3 square root body weight/nasoanal length), gonadal fat (GF) and food intake in rats and the VT effect on these parameters. Fifty-two male Wistar rats were used, divided into two groups of twenty-six animals each. SALINA group was subcutaneously injected with saline solution 12.5% (1.25 mg/g body weight/dayly/5days) and MSG group was injected with MSG solution 24% (4 mg/g body weight/dayly/5 days), after the rats were born, in the posterior cervical region without adverse reaction. On the 30th day of life, the animals were divided into four groups and submitted to operation, sham operated groups (LAPSAL and LAPMSG) and vagotomized groups (VTSAL and VTMSG). On operation day, weight, nasoanal length (NAL) and Lee index were obtained. Food intake were obtained from the 30th day until 90th day. On 90th day, the animals were sacrificed and weight, NAL, Lee index and GF were obtained. Statistic analysis was performed by the Students t test. NAL was smaller and Lee index was greater in MSG group than in SALINA group on the 30th day. Weight gain and NAL were smaller and Lee index and GF were greater on the 90th day, and food intake from the 30th to 90th day period was greater in LAPMSG group than in LAPSAL group. Weight gain, Lee index and GF were smaller and NAL trended smaller in VTMSG group than in LAPMSG group on the 90th day. Food intake from the 30th to 60th day period was smaller and trended greater from the 60th to 90th day period in VTMSG group than in LAPMSG. Vagal regeneration could explain that food intake inversion. These results suggest that MSG injected neonatally increases food intake and adiposity, and decreases stature and weight gain from the 30th to 90th day period. VT performed on the 30th day decreases food intake in first 30 days and decreases weight gain, Lee index and GF in MSG injected rats.
SUBJECT HEADINGS: Obesity. Rats. MSG. Vagotomy.
Endereço para correspondência:
Fernando de Souza
Av. Luiz Teixeira Mendes, 1888 Z. 05
Maringá PR
87015-000
e-mail: gastro@teracom.com.br
Data do recebimento: 02/09/2000
Data da revisão: 01/11/2000
Data da aprovação: 18/12/2000
- 1. Rohner-jeanrenaud F. A neuroendocrine reappraisal of the dual-centre hypothesis: its implications for obesity and insulin resistance. In: -----. International Journal of Obesity. Geneva, Switzerland: Stockton Press; 1995. p 517-34.
- 2. Flier JS, Foster DW. Eating disorders: obesity, anorexia nervosa, and bulimia nervosa. In: Wilson JD, Foster DW, Kronenberg HM, Larsen PR Williams textbook of endocrinology. Philadelphia, Pennsylvania: W B Saunders Company; 1998. p 1061-97.
- 3. Bray GA. Obesity. Dis Mon 1979;26(1):1-85.
- 4. Dawson R, Pelleymounter MA, Millard WJ, Liu S, Eppler B. Attenuation of leptin-mediated effects by monosodium glutamate-induced arcuate nucleus damage. Am J Physiol 1997;273(1PT1):E202-6.
- 5. Remke H, Wilsdorf A, Muller F. Development of hypothalamic obesity in growing rats. Exp Pathol 1988;33(4):223-32.
- 6. Nemeroff CB, Grant LD, Bissette G, Ervin GN, Harrel LE, Prange Jr. AJ. Growth, endocrinological and behavioral deficits after monosodium L-glutamate in the neonatal rat: possible involvement of arcuate dopamine neuron damage. Psychoneuroendocrinology 1977;2(2):179-96.
- 7. Miskowiak B, Partika M. Effects of neonatal treatment with MSG (Monosodium glutamate) on hypothalamo-pituitary-thyroid axis in adult male rats. Histol Histopathol 1993;8(4):731-4.
- 8. Silva ACM. Obesidade hipotalâmica induzida por glutamato monossódico: alteraçőes pancreáticas [Tese - Mestrado]. Universidade Estadual de Maringá; 1995.
- 9. Seress L. Divergent effects of acute and chronic primary monosodium L-glutamate treatment on the anterior and posterior parts of the arcuate nucleus. Neuroscience 1982;7(9):2207-16.
- 10. Nascimento Curi CMO, Marmo MR, Egami M, Ribeiro EB, Andrade IS, Dolnikoff MS. Effect of monosodium glutamate treatment during neonatal development on lipogenesis rate and lipoprotein lipase activity in adult rats. Biochem Int 1991;24(5):927-35.
- 11. Holzwarth-Mcbride MA, Hurst EM, Knigge KM. Monosodium glutamate induced lesions of the arcuate nucleus. I: Endocrine deficiency and ultrastructure of the median eminence. Anat Rec 1976;186(2):185-96.
- 12. Holzwarth-Mcbride MA, Sladek Jr JR, Knigge, KM. Monosodium glutamate induced lesions of the arcuate nucleus. II: Fluorescence histochemistry of catecholamines. Anat Rec 1976;186(2):197-205.
- 13. Sclafani A. Effects of gastrointestinal surgery on ingestive behavior in animals. Gastroenterol Clin North Am 1987;16(3):461-77.
- 14. Souza F. Induçăo de obesidade em ratos através do glutamato monossódico: Avaliaçăo biométrica [Tese - Mestrado]. Universidade Federal do Paraná; 1999.
- 15. Stephens DN. Does the Lee obesity index measure general obesity? Physiol Behav 1980;25(2):313-5.
- 16. Bernardis LL, Patterson BD. Correlation between "Lee index" and carcass fat content in weanling and adult female rats with hypothalamic lesions. J Endocrinol 1968;40(4):527-8.
- 17. Coscina DV, Chambers JW, Park I, Hogan S, Himms-Hagen J. Impaired diet-induced thermogenesis in brown adipose tissue from rats made obese with parasagittal hypothalamic knife-cuts. Brain Res Bull 1985;14(6):585-93.
- 18. Dolnikoff MS, Kater CE, Egami M, Andrade IS, Marmo MR. Neonatal treatment with monosodium glutamate increases plasma corticosterone in the rats. Neuroendocrionology 1988;48(6):645-9.
- 19. Bonfleur ML. Atividade colinérgica de áreas cerebrais em ratos tratados com glutamato monossódico [Dissertaçăo - Bacharelado]. Universidade Estadual do Oeste do Paraná; 1998.
- 20. Kanarek RB, Marks-Kaufman R. Increased carbohydrate consumption induced by neonatal administration of monosodium glutamate to rats. Neurobehav Toxicol Teratol 1981;3(3):343-50.
- 21. Chikamori K, Masuda K, Isumi H, Isaka K, Tezuka, U. Effect of vagotomy on hyperinsulinemia in obese rats with hypothalamic lesions. Endocrinol Jpn 1977;24(3):251-8.
- 22. Abe M, Saito M, Shimazu T. Neuropeptide Y in the specific hypothalamic nuclei of rats treated neonatally with monosodium glutamate. Brain Res Bull 1990;24(2):289-91.
- 23. Frankish HM, Dryden S, Hopkins D, Wang Q, Williams G. Neuropeptide Y, the hypothalamus, and diabetes: insights into the central control of metabolism. Peptides 1995;16(4):757-71.
- 24. Miller RE. Pancreatic neuroendocrinology: peripheral neural mechanisms in the regulation of the islets of Langerhans. Endocr Rev 1981;2(4):471-94.
- 25. Atef N, Ktorza A, Picon L, Pénicaud L. Increased islet blood flow in obese rats: role of the autonomic nervous system. Am J Physiol 1992;262(5PT1):E736-40.
- 26. Powley TL, Opsahl CA. Autonomic components of the hypothalamic feeding syndromes. In: Novin D, Wyrwicka W, Bray G. Hunger: basic mechanisms and clinical implications. New York: Raven Press; 1976. p 313-26.
- 27. Opsahl AC, Powley TL. Failure of vagotomy to reverse obesity in the genetically obese Zucker rat. Am J Physiol 1974;226(1):34-8.
- 28. Campfield LA, Smith FJ, LeMagnen J. Altered endocrine pancreatic function following vagotomy: possible behavioral and metabolic bases for assessing completeness of vagotomy. J Auton Nerv Syst 1983;9(1):283-300.
- 29. Sclafani A, Lucas F. Abdominal vagotomy does not block carbohydrate-conditioned flavor preferences in rats. Physiol Behav 1996;60(2):447-53.
- 30. King BM, Phelps GR, Frohman LA. Hypothalamic obesity in female rats in absence of vagally mediated hyperinsulinemia. Am J Physiol 1980;239(6):E437-41.
Datas de Publicação
-
Publicação nesta coleção
11 Set 2003 -
Data do Fascículo
Mar 2001
Histórico
-
Aceito
18 Dez 2000 -
Revisado
01 Nov 2000 -
Recebido
02 Set 2000